The Regulatory Role of CircAGGF1 in Myogenic Differentiation and Skeletal Muscle Development


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture
2.3. RNA Extraction and qRT-PCR
2.4. Immunofluorescence Staining
2.5. Luciferase Reporter Assay
2.6. Vector Construction and siRNA
2.7. Western Blot
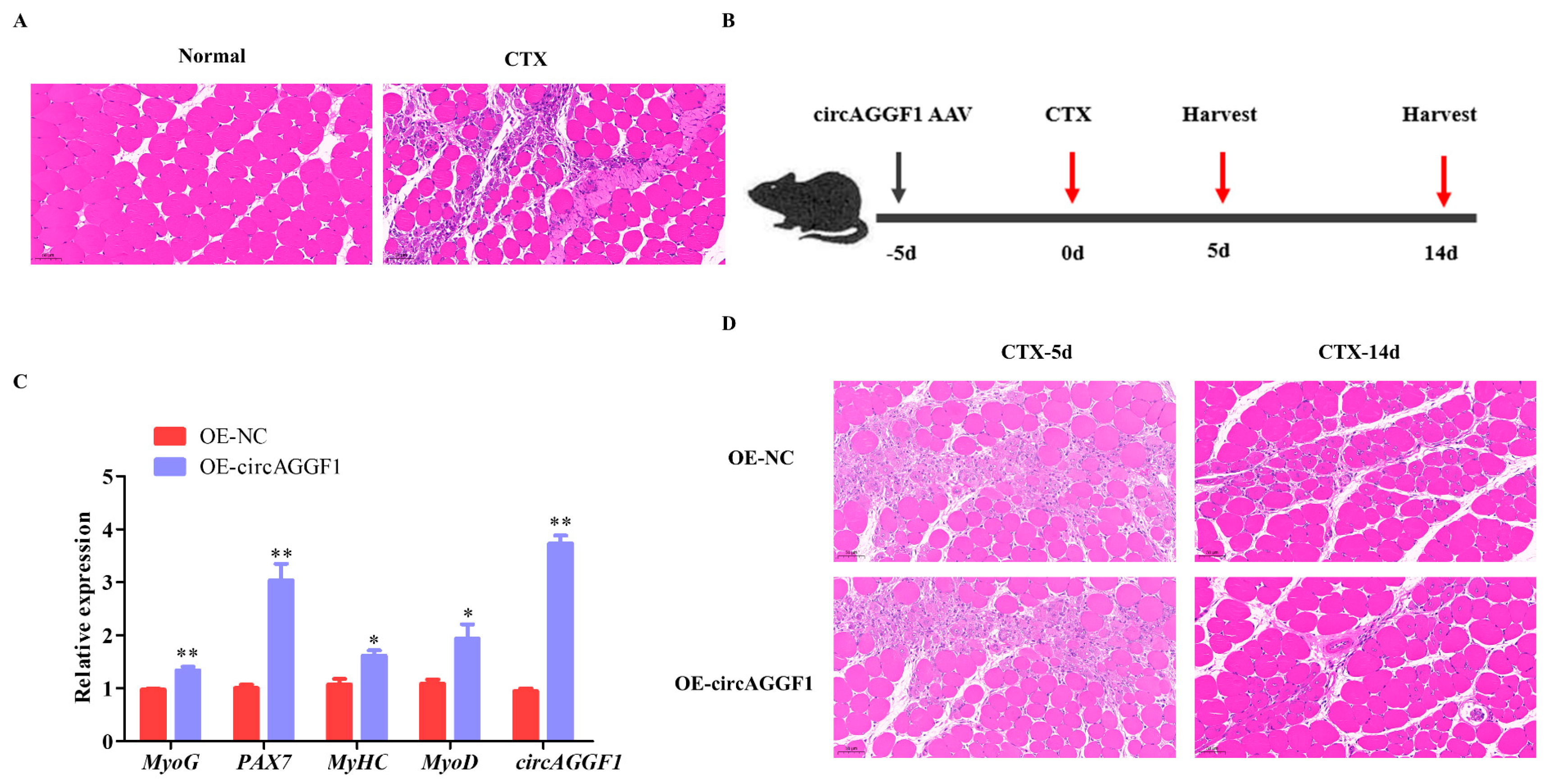
2.8. Animal Treatment and Establishment of Muscle Injury Animal Model
2.9. Statistical Analysis
3. Results
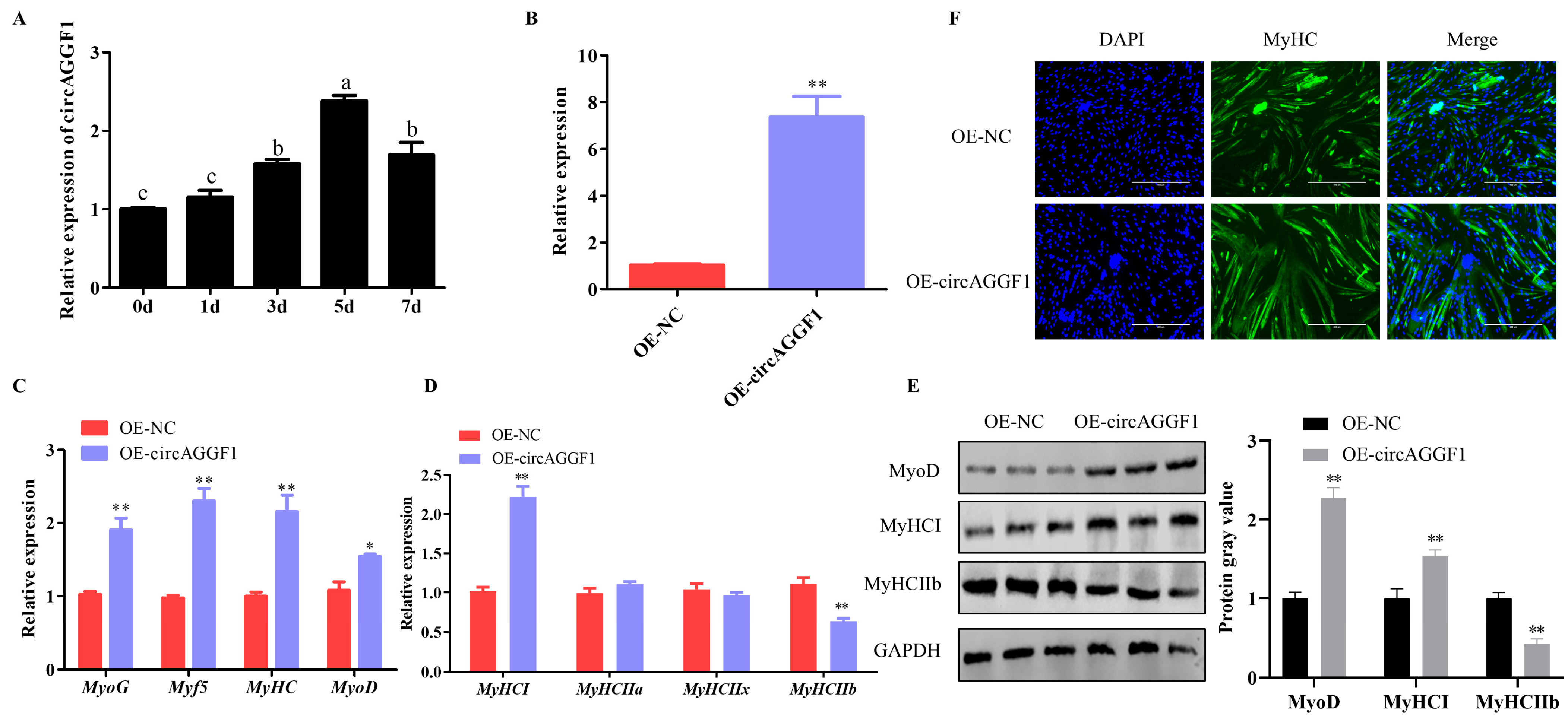
3.1. circAGGF1 Promotes Differentiation and Alters the Muscle Fiber Composition in C2C12 Cells
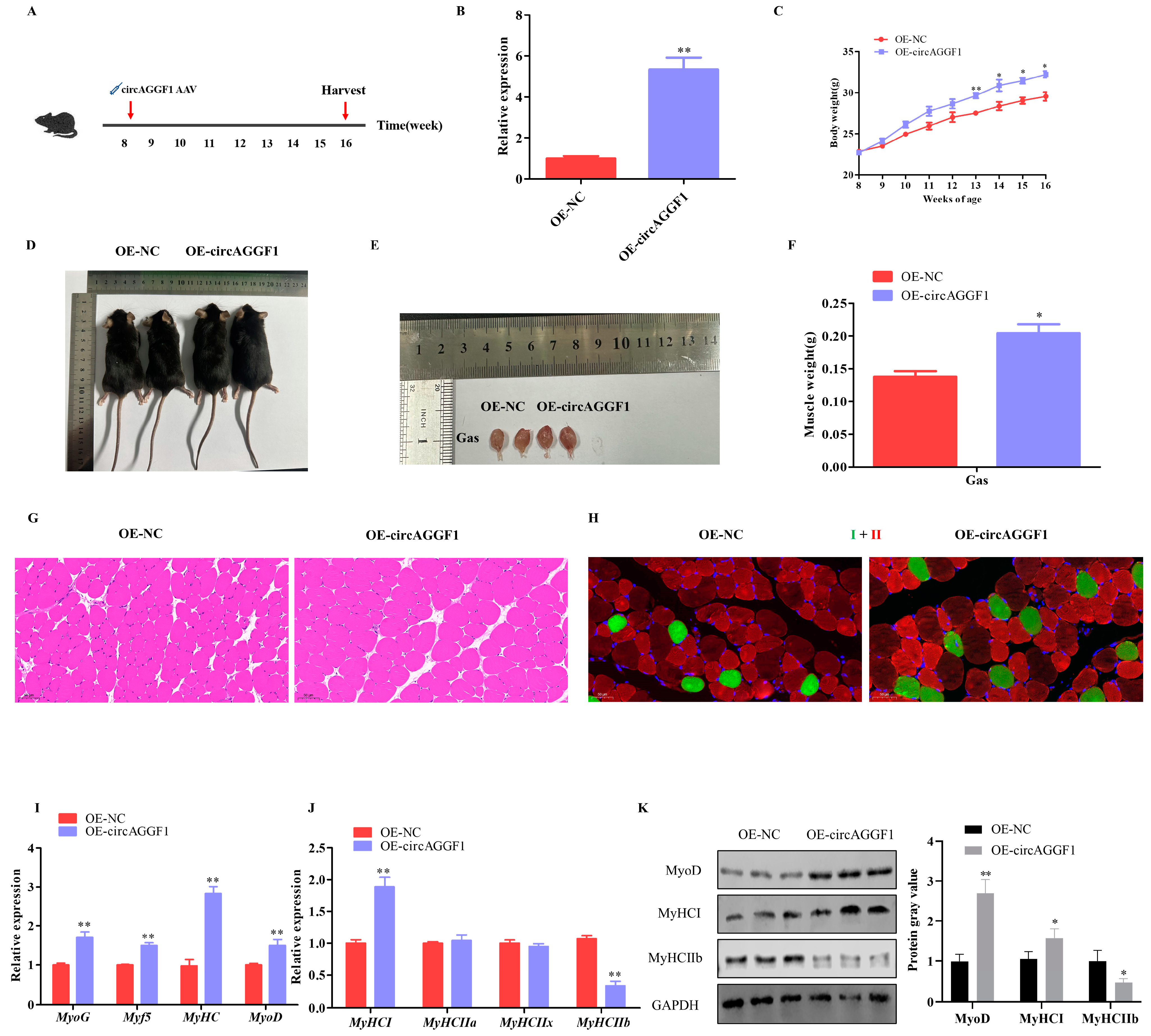
3.2. circAGGF1 Promotes Mice Myogenesis In Vivo
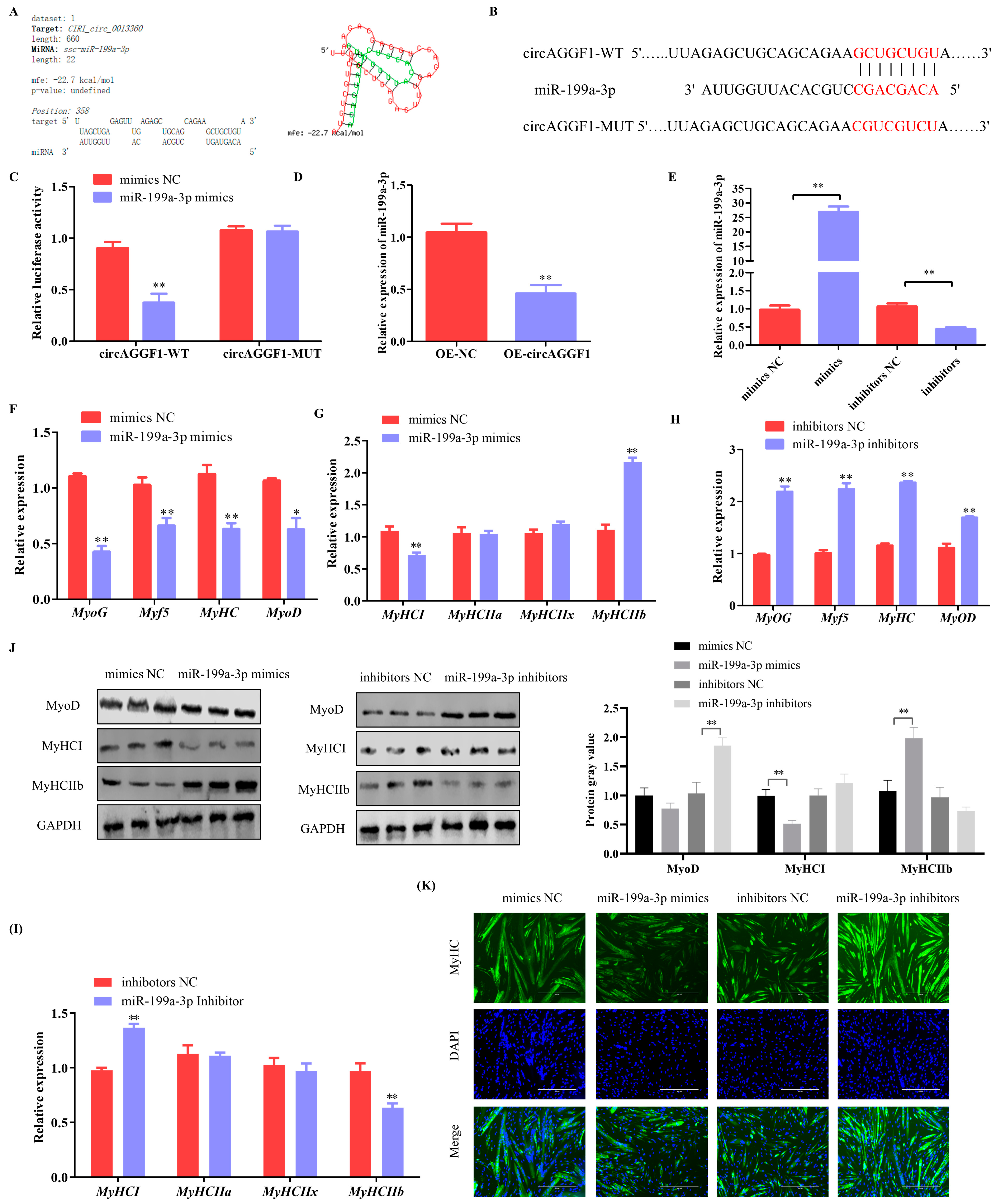
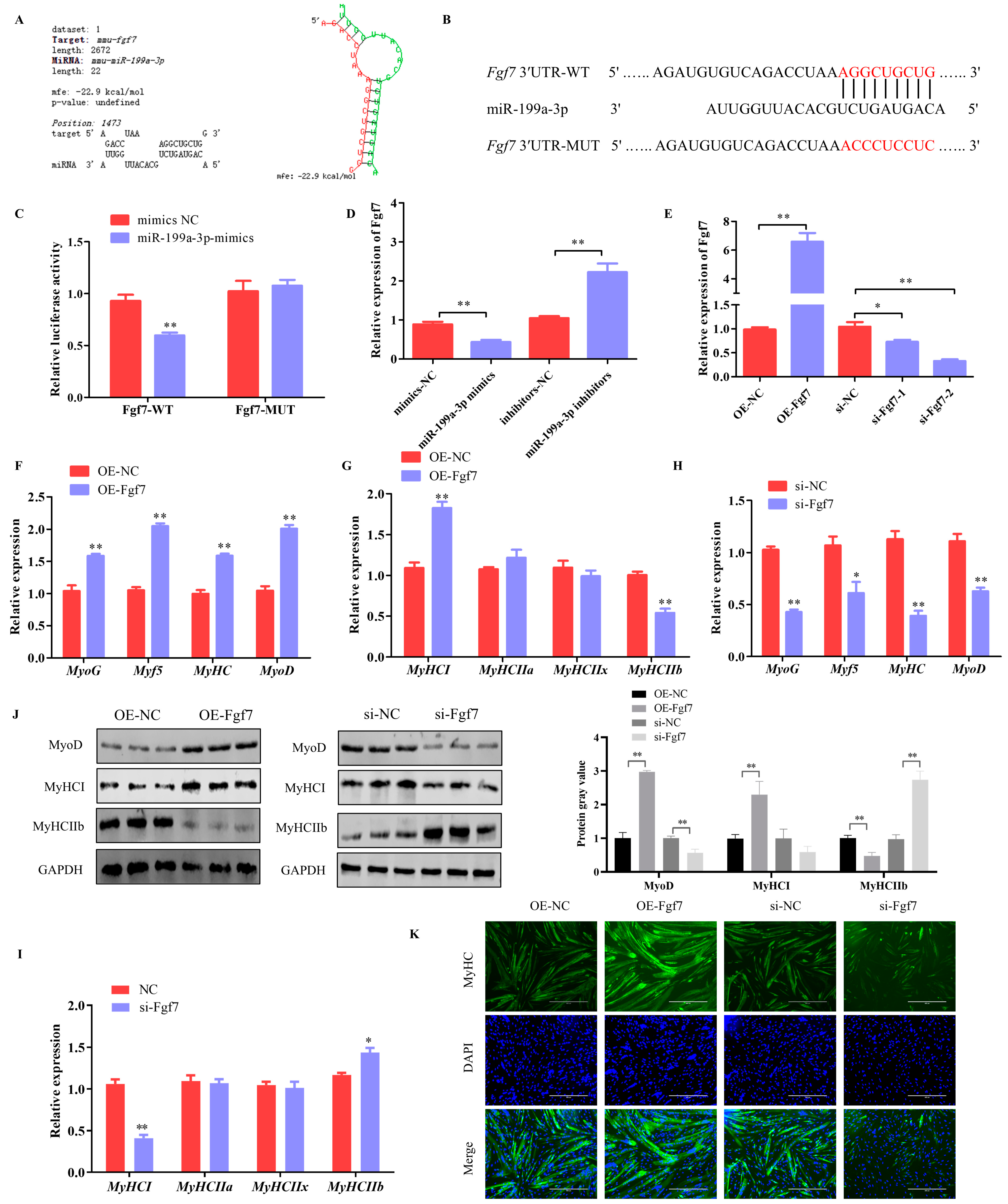
3.3. miR-199a-3p Inhibits Differentiation and Alters the Muscle Fiber Composition in C2C12 Cells
3.4. Fgf7 Promotes Differentiation and Alters the Muscle Fiber Composition in C2C12 Cells
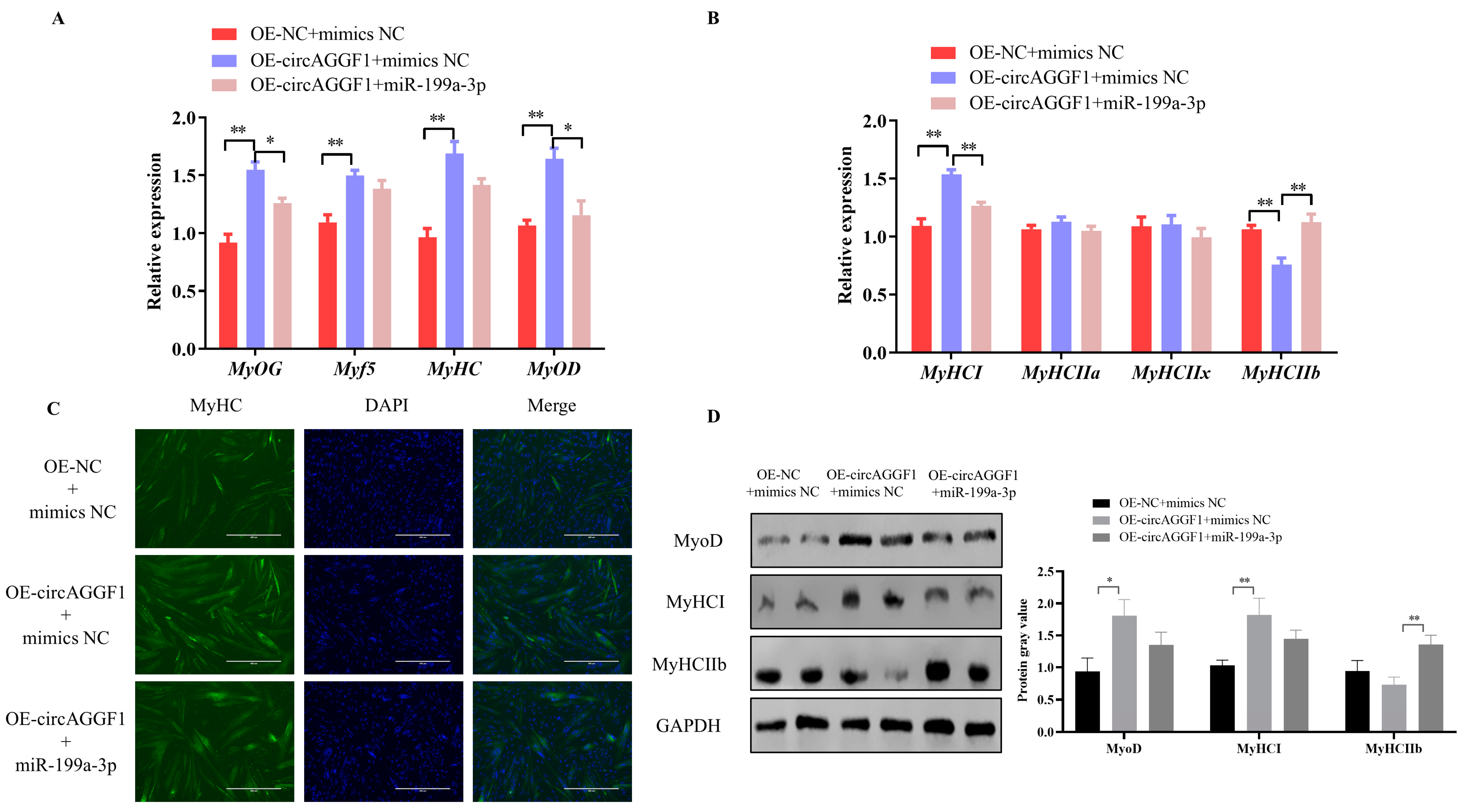
3.5. CircAGGF1 Promotes Myoblast Differentiation and Myofiber Type Transformation via miR-199a-3p
4. Discussion
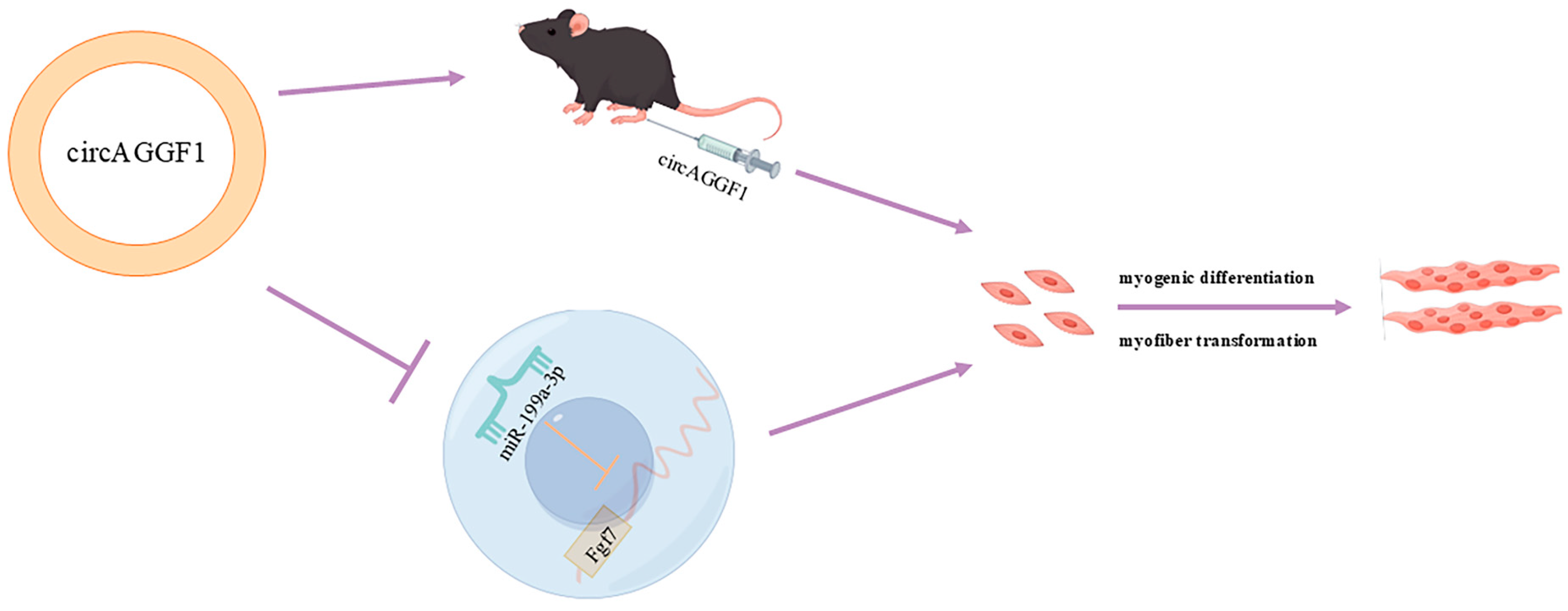
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dayanidhi, S.; Lieber, R.L. Skeletal muscle satellite cells: Mediators of muscle growth during development and implications for developmental disorders. Muscle Nerve 2014, 50, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Kopantseva, E.E.; Belyavsky, A.V. [Key regulators of skeletal myogenesis]. Mol. Biol. 2016, 50, 195–222. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Cui, C.; Wang, Y.; He, H.; Liu, Z.; Shen, X.; Chen, Y.; Li, D.; Zhu, Q.; Yin, H. Knockdown of CSRP3 inhibits differentiation of chicken satellite cells by promoting TGF-β/Smad3 signaling. Gene 2019, 707, 36–43. [Google Scholar] [CrossRef]
- Liang, G.; Yang, Y.; Niu, G.; Tang, Z.; Li, K. Genome-wide profiling of Sus scrofa circular RNAs across nine organs and three developmental stages. DNA Res. 2017, 24, 523–535. [Google Scholar] [CrossRef]
- Ouyang, H.; Chen, X.; Wang, Z.; Yu, J.; Jia, X.; Li, Z.; Luo, W.; Abdalla, B.A.; Jebessa, E.; Nie, Q.; et al. Circular RNAs are abundant and dynamically expressed during embryonic muscle development in chickens. DNA Res. 2018, 25, 71–86. [Google Scholar] [CrossRef]
- Chen, W.; Schuman, E. Circular RNAs in Brain and Other Tissues: A Functional Enigma. Trends Neurosci. 2016, 39, 597–604. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, X.; Odame, E.; Xu, X.; Chen, Y.; Ye, J.; Zhou, H.; Dai, D.; Kyei, B.; Zhan, S.; et al. CircRNA-Protein Interactions in Muscle Development and Diseases. Int. J. Mol. Sci. 2021, 22, 3262. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Nie, L.; Ding, X.; Zhang, X.; Zhao, W.; Xu, X.; Kyei, B.; Dai, D.; Zhan, S.; et al. MyoD-induced circular RNA CDR1as promotes myogenic differentiation of skeletal muscle satellite cells. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 807–821. [Google Scholar] [CrossRef]
- Shi, Y.; Jia, X.; Xu, J. The new function of circRNA: Translation. Clin. Transl. Oncol. 2020, 22, 2162–2169. [Google Scholar] [CrossRef]
- Xu, J.; Wen, Y.; Li, X.; Peng, W.; Zhang, Z.; Liu, X.; Yang, P.; Chen, N.; Lei, C.; Zhang, J.; et al. Bovine enhancer-regulated circSGCB acts as a ceRNA to regulate skeletal muscle development via enhancing KLF3 expression. Int. J. Biol. Macromol. 2024, 261, 129779. [Google Scholar] [CrossRef]
- Qi, A.; Ru, W.; Yang, H.; Yang, Y.; Tang, J.; Yang, S.; Lan, X.; Lei, C.; Sun, X.; Chen, H. Circular RNA ACTA1 Acts as a Sponge for miR-199a-5p and miR-433 to Regulate Bovine Myoblast Development through the MAP3K11/MAP2K7/JNK Pathway. J. Agric. Food Chem. 2022, 70, 3357–3373. [Google Scholar] [CrossRef]
- Chen, M.; Wei, X.; Song, M.; Jiang, R.; Huang, K.; Deng, Y.; Liu, Q.; Shi, D.; Li, H. Circular RNA circMYBPC1 promotes skeletal muscle differentiation by targeting MyHC. Mol. Therapy. Nucleic Acids 2021, 24, 352–368. [Google Scholar] [CrossRef]
- Lin, Z.; Xie, F.; He, X.; Wang, J.; Luo, J.; Chen, T.; Jiang, Q.; Xi, Q.; Zhang, Y.; Sun, J. A novel protein encoded by circKANSL1L regulates skeletal myogenesis via the Akt-FoxO3 signaling axis. Int. J. Biol. Macromol. 2024, 257, 128609. [Google Scholar] [CrossRef]
- Li, M.; Zhang, N.; Zhang, W.; Hei, W.; Cai, C.; Yang, Y.; Lu, C.; Gao, P.; Guo, X.; Cao, G.; et al. Comprehensive analysis of differentially expressed circRNAs and ceRNA regulatory network in porcine skeletal muscle. BMC Genom. 2021, 22, 320. [Google Scholar] [CrossRef]
- Mohammadabadi, M.; Bordbar, F.; Jensen, J.; Du, M.; Guo, W. Key Genes Regulating Skeletal Muscle Development and Growth in Farm Animals. Animals 2021, 11, 835. [Google Scholar] [CrossRef]
- Weskamp, K.; Olwin, B.B.; Parker, R. Post-Transcriptional Regulation in Skeletal Muscle Development, Repair, and Disease. Trends Mol. Med. 2021, 27, 469–481. [Google Scholar] [CrossRef]
- Ding, H.; Heng, B.; He, W.; Shi, L.; Lai, C.; Xiao, L.; Ren, H.; Mo, S.; Su, Z. Chronic reactive oxygen species exposure inhibits glucose uptake and causes insulin resistance in C2C12 myotubes. Biochem. Biophys. Res. Commun. 2016, 478, 798–803. [Google Scholar] [CrossRef]
- Wong, C.Y.; Al-Salami, H.; Dass, C.R. C2C12 cell model: Its role in understanding of insulin resistance at the molecular level and pharmaceutical development at the preclinical stage. J. Pharm. Pharmacol. 2020, 72, 1667–1693. [Google Scholar] [CrossRef] [PubMed]
- Nezhad, F.Y.; Riermeier, A.; Schönfelder, M.; Becker, L.; de Angelis, M.H.; Wackerhage, H. Skeletal muscle phenotyping of Hippo gene-mutated mice reveals that Lats1 deletion increases the percentage of type I muscle fibers. Transgenic Res. 2022, 31, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, D.L.; Roberts, M.D.; Haun, C.T.; Schoenfeld, B.J. Muscle Fiber Type Transitions with Exercise Training: Shifting Perspectives. Sports 2021, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yang, T.; Zheng, Y.; Nie, Q.; Chen, J.; Li, Q.; Ren, X.; Yin, X.; Wang, S.; Yan, Y.; et al. EIF4A3-Induced Circular RNA CircDdb1 Promotes Muscle Atrophy through Encoding a Novel Protein CircDdb1-867aa. Adv. Sci. 2024, 11, e2406986. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Fang, Q.; Ma, W.; Zhang, Q.; Qiu, J.; Gu, X.; Yang, H.; Sun, H. Skeletal Muscle Atrophy Was Alleviated by Salidroside Through Suppressing Oxidative Stress and Inflammation During Denervation. Front. Pharmacol. 2019, 10, 997. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Mammucari, C.; Milan, G.; Romanello, V.; Masiero, E.; Rudolf, R.; Del Piccolo, P.; Burden, S.J.; Di Lisi, R.; Sandri, C.; Zhao, J.; et al. FoxO3 controls autophagy in skeletal muscle in vivo. Cell Metab. 2007, 6, 458–471. [Google Scholar] [CrossRef]
- Zhao, J.; Brault, J.J.; Schild, A.; Cao, P.; Sandri, M.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 2007, 6, 472–483. [Google Scholar] [CrossRef]
- Chen, R.; Yang, T.; Jin, B.; Xu, W.; Yan, Y.; Wood, N.; Lehmann, H.I.; Wang, S.; Zhu, X.; Yuan, W.; et al. CircTmeff1 Promotes Muscle Atrophy by Interacting with TDP-43 and Encoding A Novel TMEFF1-339aa Protein. Adv. Sci. 2023, 10, e2206732. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Lian, C.; Chen, G.; Zou, P.; Qin, B.G. CircRNA FUT10 regulates the regenerative potential of aged skeletal muscle stem cells by targeting HOXA9. Aging 2021, 13, 17428–17441. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, X.; Xu, P.; Wang, H.; Wang, S.; Zhang, L.; Li, Z.; Xie, L.; Sun, G.; Xia, Y.; et al. Circular RNA circLMO7 acts as a microRNA-30a-3p sponge to promote gastric cancer progression via the WNT2/β-catenin pathway. J. Exp. Clin. Cancer Res. 2021, 40, 6. [Google Scholar] [CrossRef]
- Li, H.; Wei, X.; Yang, J.; Dong, D.; Hao, D.; Huang, Y.; Lan, X.; Plath, M.; Lei, C.; Ma, Y.; et al. circFGFR4 Promotes Differentiation of Myoblasts via Binding miR-107 to Relieve Its Inhibition of Wnt3a. Mol. Therapy. Nucleic Acids 2018, 11, 272–283. [Google Scholar] [CrossRef]
- Ebbesen, K.K.; Kjems, J.; Hansen, T.B. Circular RNAs: Identification, biogenesis and function. Biochim. Biophys. Acta 2016, 1859, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, H.; Dai, Y.; Li, S.; Gu, J.; Wu, R.; Jia, J.; Shen, J.; Zhang, Y.; Li, H.; et al. CircITGB5 regulates the proliferation and adipogenic differentiation of chicken intramuscular preadipocytes through the miR-181b-5p/CPT1A axis. Int. J. Biol. Macromol. 2024, 283, 137608. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, Y.; Deng, K.; Liang, Y.; Zhang, G.; Gao, X.; El-Samahy, M.A.; Zhang, Y.; Deng, M.; Wang, F. Circular RNA circUSP13 sponges miR-29c to promote differentiation and inhibit apoptosis of goat myoblasts by targeting IGF1. FASEB J. 2022, 36, e22097. [Google Scholar] [CrossRef]
- Wu, S.; Shao, T.; Xie, J.; Li, J.; Sun, L.; Zhang, Y.; Zhao, L.; Wang, L.; Li, X.; Zhang, L.; et al. MiR-199a-3p regulates HCT-8 cell autophagy and apoptosis in response to Cryptosporidium parvum infection by targeting MTOR. Commun. Biol. 2024, 7, 924. [Google Scholar] [CrossRef]
- Zhuo, L.; Zhou, Y.; Tian, J.; Li, Y.; Xie, Z.; Pei, C.; Yan, B.; Ma, L. The role of miR-199a-3p in inhibiting the proliferation of spermatogonial stem cells under heat stress. Theriogenology 2023, 211, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, X.; Li, H.; Mou, Y.; Cui, G. SP1 promotes high glucose-induced lens epithelial cell viability, migration and epithelial-mesenchymal transition via regulating FGF7 and PI3K/AKT pathway. Int. Ophthalmol. 2024, 44, 316. [Google Scholar] [CrossRef]
- Fu, W.; Liu, L.; Tong, S. Berberine inhibits the progression of breast cancer by regulating METTL3-mediated m6A modification of FGF7 mRNA. Thorac. Cancer 2024, 15, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, W.H.; Yang, B.; Milman Krentsis, I. Primer on fibroblast growth factor 7 (FGF 7). Differentiation 2024, 139, 100801. [Google Scholar] [CrossRef]
- Zinkle, A.; Mohammadi, M. Structural Biology of the FGF7 Subfamily. Front. Genet. 2019, 10, 102. [Google Scholar] [CrossRef]
- Forbes, S.J.; Themis, M.; Alison, M.R.; Sarosi, I.; Coutelle, C.; Hodgson, H.J. Synergistic growth factors enhance rat liver proliferation and enable retroviral gene transfer via a peripheral vein. Gastroenterology 2000, 118, 591–598. [Google Scholar] [CrossRef]
- Takase, H.M.; Itoh, T.; Ino, S.; Wang, T.; Koji, T.; Akira, S.; Takikawa, Y.; Miyajima, A. FGF7 is a functional niche signal required for stimulation of adult liver progenitor cells that support liver regeneration. Genes Dev. 2013, 27, 169–181. [Google Scholar] [CrossRef]
- Alwahsh, S.M.; Qutachi, O.; Starkey Lewis, P.J.; Bond, A.; Noble, J.; Burgoyne, P.; Morton, N.; Carter, R.; Mann, J.; Ferreira-Gonzalez, S.; et al. Fibroblast growth factor 7 releasing particles enhance islet engraftment and improve metabolic control following islet transplantation in mice with diabetes. Am. J. Transplant. 2021, 21, 2950–2963. [Google Scholar] [CrossRef]
- Ma, L.; Meng, Y.; An, Y.; Han, P.; Zhang, C.; Yue, Y.; Wen, C.; Shi, X.; Jin, J.; Yang, G.; et al. Single-cell RNA-seq reveals novel interaction between muscle satellite cells and fibro-adipogenic progenitors mediated with FGF7 signalling. J. Cachexia Sarcopenia Muscle 2024, 15, 1388–1403. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primers Sequences (5′→3′) | Annealing Temperature (°C) | Product Length (bp) |
|---|---|---|---|
| circAGGF1 | F: ACTGTGATGTGGAAAGTGGTCG R: CTTCGTTTCTCCCACAATGGAG | 59 | 189 |
| MyoD | F: TGCTCTGATGGCATGATGGATT R: AGATGCGCTCCACTATGCTG | 60 | 163 |
| MyoG | F: GTCCCAACCCAGGAGATCAT R: CACGATGGACGTAAGGGAG | 58 | 77 |
| Myf5 | F: CTCTGACGGCATGCCTGAAT R: AGCTCGGATGGCTCTGTAG | 58 | 177 |
| MyHC | F:GGATGGGATATAAAGGGGCTGG R: ATCCAGAGATCCTGGGTTGGA | 59 | 72 |
| MyHCIIx | F: GCATCCCTAAAGGCAGGCT R: GTTCTGAGCCTCGATTCGCT | 60 | 139 |
| MyHCIIa | F:AGGAGCTTACTTACCAGACAGA R: TCGCTTCGGTCATTCCACAG | 57 | 299 |
| MyHCIIb | F: CTCACCTACCAGACCGAGGA R: CTCCTGTCACCTCTCAACAGA | 59 | 290 |
| MyHCI | F: AGGTCTGGCTCTGAGCATTC R: CCTTTCTCGGAGCCACCTTG | 60 | 244 |
| GAPDH | F: CTTCTCCTGCAGCCTCGT R: ATGAAGGGGTCGTTGATGGC | 59 | 137 |
| U6 | F: CTCGCTTCGGCAGCACA R: AACGCTTCACGAATTTGCGT | 60 | 185 |
| PAX7 | F: GAGAACCCCGGGATGTTCAG R: ATCGAACTCACTGAGGGCAC | 60 | 92 |
| miR-199a-3p | GCGCGACAGTAGTCTGCACAT | ||
| Fgf7 | F: TATTCATGAACACCCGGGGC R: CAGTTCACACTCGTAGCCGT | 60 | 168 |
| Names | Sequences (5′→3′) |
|---|---|
| miR-199a-3p mimics | F: ACAGUAGUCUGCACAUUGGUUAACC R: AAUGUGCAGACUACUGUUU |
| miR-199a-3p inhibitor | UAACCAAUGUGCAGACUACUGU |
| si-Fgf7-1 | CACCUAUGCAUCAGCUAAATT |
| si-Fgf7-2 | GGUACCUGAGGAUUGACAATT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hei, W.; Gong, Y.; Cai, W.; Li, R.; Chen, J.; Zhang, W.; Ji, M.; Li, M.; Yang, Y.; Cai, C.; et al. The Regulatory Role of CircAGGF1 in Myogenic Differentiation and Skeletal Muscle Development. Animals 2025, 15, 708. https://doi.org/10.3390/ani15050708
Hei W, Gong Y, Cai W, Li R, Chen J, Zhang W, Ji M, Li M, Yang Y, Cai C, et al. The Regulatory Role of CircAGGF1 in Myogenic Differentiation and Skeletal Muscle Development. Animals. 2025; 15(5):708. https://doi.org/10.3390/ani15050708
Chicago/Turabian StyleHei, Wei, Yuxuan Gong, Wenrun Cai, Ruotong Li, Jiayi Chen, Wanfeng Zhang, Mengting Ji, Meng Li, Yang Yang, Chunbo Cai, and et al. 2025. "The Regulatory Role of CircAGGF1 in Myogenic Differentiation and Skeletal Muscle Development" Animals 15, no. 5: 708. https://doi.org/10.3390/ani15050708
APA StyleHei, W., Gong, Y., Cai, W., Li, R., Chen, J., Zhang, W., Ji, M., Li, M., Yang, Y., Cai, C., Guo, X., & Li, B. (2025). The Regulatory Role of CircAGGF1 in Myogenic Differentiation and Skeletal Muscle Development. Animals, 15(5), 708. https://doi.org/10.3390/ani15050708