
Effects of Climate and Land Use on Different Facets of Mammal Diversity in Giant Panda Range

 , and
, and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area and Species Distribution Data
2.2. Calculation of Diversity Indices
2.3. Environmental Predictors of Mammal Diversity
2.4. Statistical Analysis
3. Results
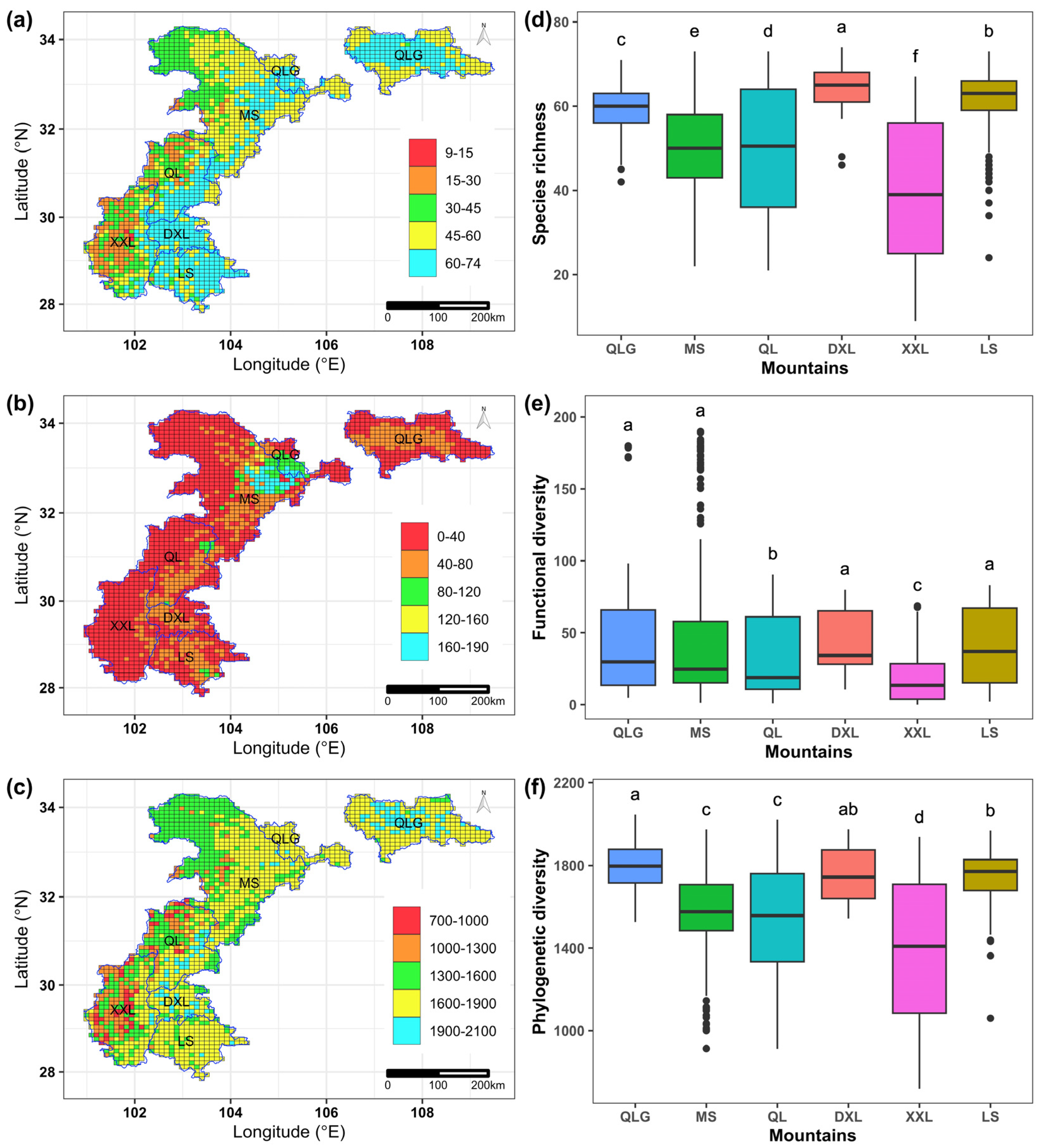
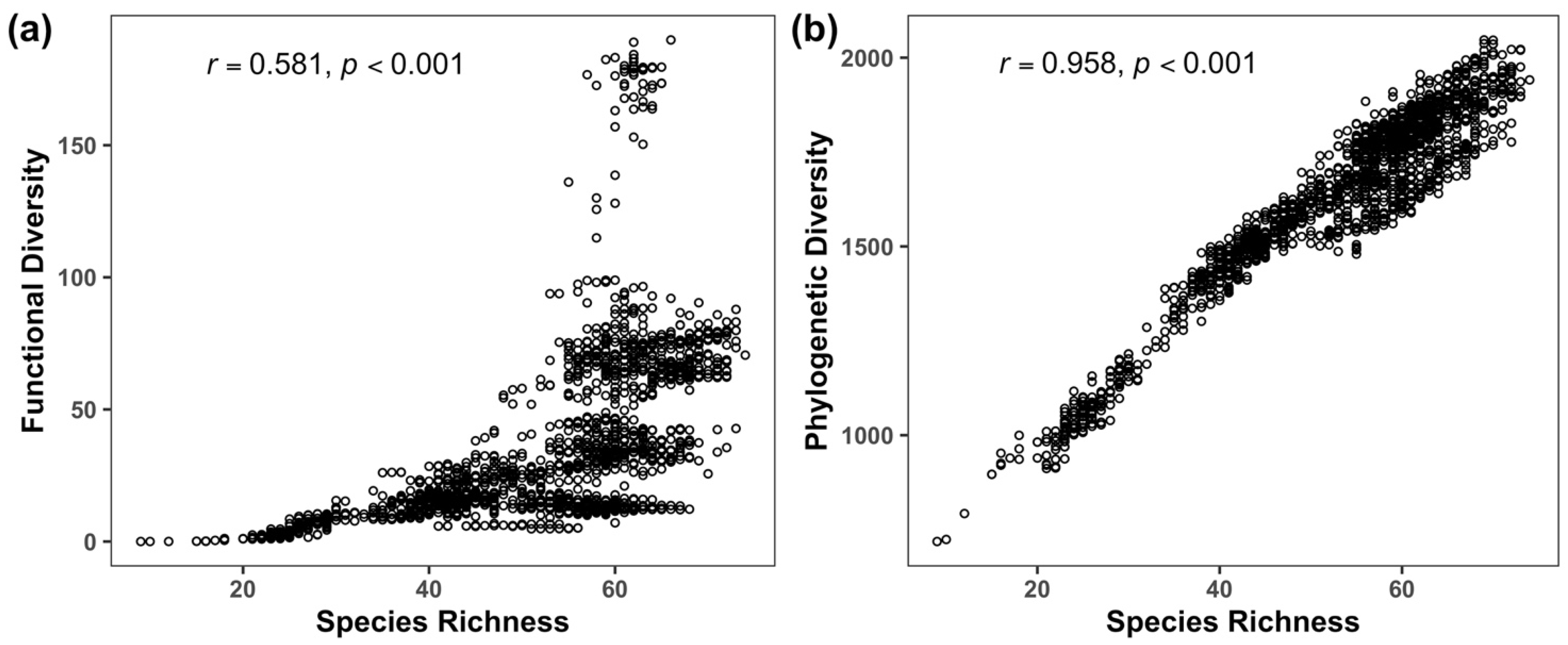
3.1. Geographic Patterns of Taxonomic, Functional and Phylogenetic Diversity
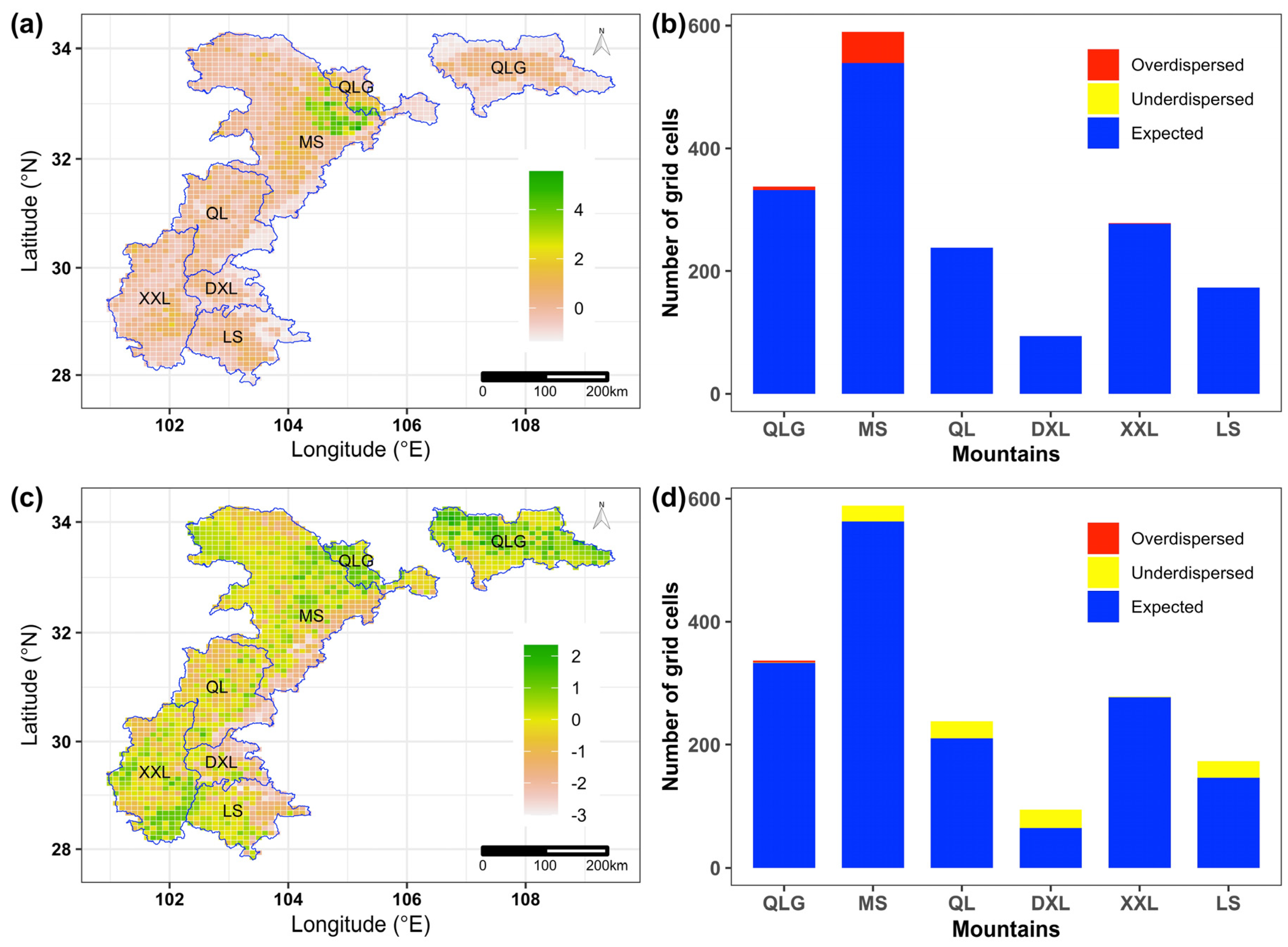
3.2. Geographic Patterns of Richness-Controlled Functional and Phylogenetic Diversity
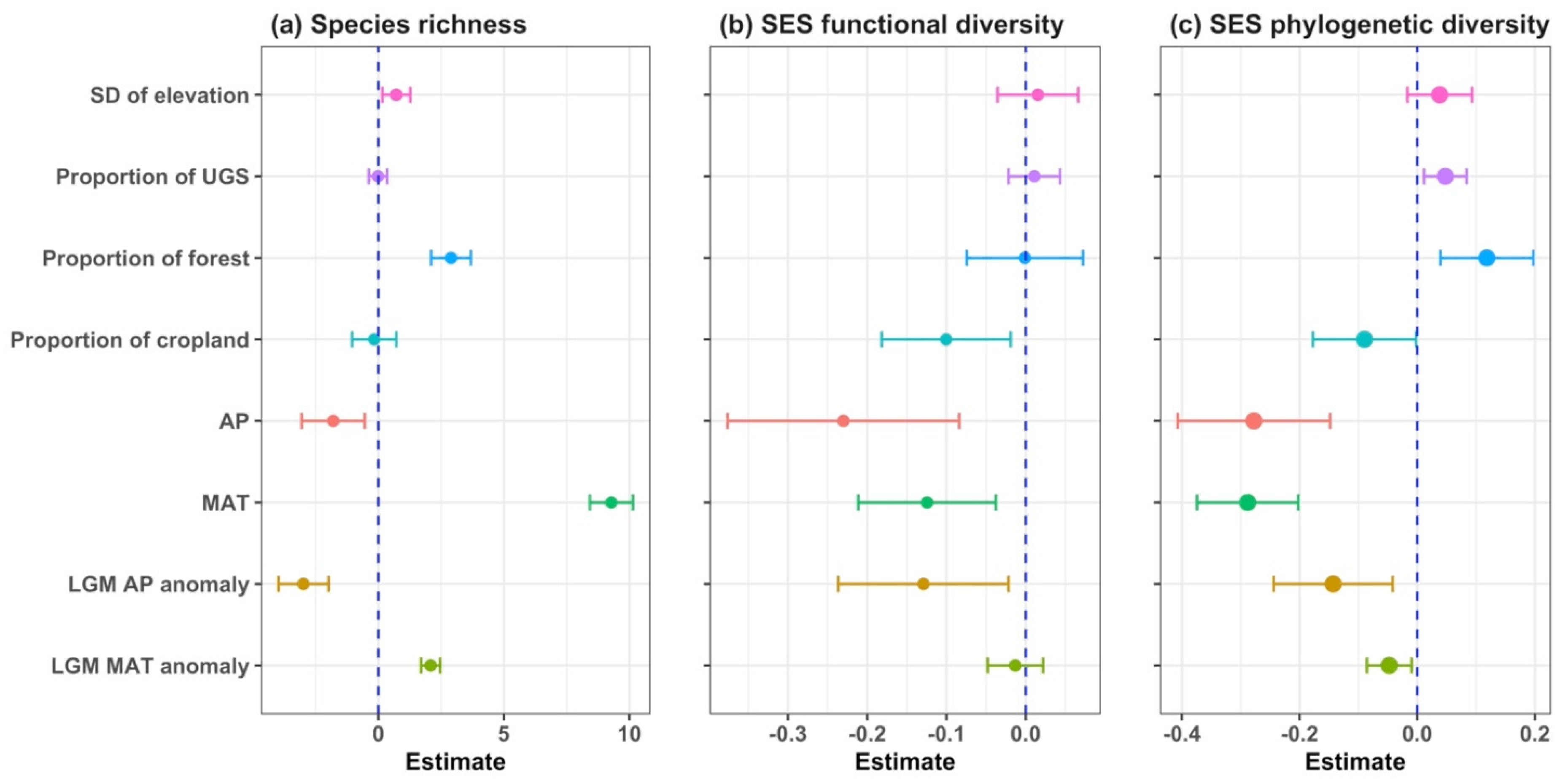
3.3. Drivers of Taxonomic, Phylogenetic and Functional Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaston, K.J. Species-range-size distributions: Patterns, mechanisms and implications. Trends Ecol. Evol. 1996, 11, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.; Yohe, G.W.; Auffhammer, M.; Huggel, C.; Molau, U.; Faus da Silva Dias, M.A.; Solow, A.; Stone, D.A.; Tibig, L.; Bouwer, L.; et al. Detection and attribution of observed impacts. In Climate Change 2014—Impacts, Adaptation and Vulnerability: Part A: Global and Sectoral Aspects: Working Group II Contribution to the IPCC Fifth Assessment Report; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef]
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef]
- Safi, K.; Cianciaruso, M.V.; Loyola, R.D.; Brito, D.; Armour-Marshall, K.; Diniz-Filho, J.A.F. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2536–2544. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Flynn, D.F.B.; Mirotchnick, N.; Jain, M.; Palmer, M.I.; Naeem, S. Functional and phylogenetic diversity as predictors of biodiversity–ecosystem-function relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Stuart-Smith, R.D.; Bates, A.E.; Lefcheck, J.S.; Duffy, J.E.; Baker, S.C.; Thomson, R.J.; Stuart-Smith, J.F.; Hill, N.A.; Kininmonth, S.J.; Airoldi, L.; et al. Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 2013, 501, 539–542. [Google Scholar] [CrossRef]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Stevens, R.D.; Tello, J.S.; Gavilanez, M.M. Stronger tests of mechanisms underlying geographic gradients of biodiversity: Insights from the dimensionality of biodiversity. PLoS ONE 2013, 8, e56853. [Google Scholar] [CrossRef]
- Montaño-Centellas, F.; Baiser, B.; McGrew, A.; Trotta, L.; Li, D. Global patterns and drivers of raptor phylogenetic and functional diversity. Glob. Ecol. Biogeogr. 2023, 32, 281–294. [Google Scholar] [CrossRef]
- Tordoni, E.; Carmona, C.P.; Toussaint, A.; Tamme, R.; Pärtel, M. Global patterns and determinants of multiple facets of plant diversity. Glob. Ecol. Biogeogr. 2024, 33, e13823. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Difficult decisions: Strategies for conservation prioritization when taxonomic, phylogenetic and functional diversity are not spatially congruent. Biol. Conserv. 2018, 225, 128–133. [Google Scholar] [CrossRef]
- Murali, G.; Gumbs, R.; Meiri, S.; Roll, U. Global determinants and conservation of evolutionary and geographic rarity in land vertebrates. Sci. Adv. 2021, 7, eabe5582. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-Y.; Serra-Diaz, J.M.; Eiserhardt, W.L.; Maitner, B.S.; Merow, C.; Violle, C.; Pound, M.J.; Sun, M.; Slik, F.; Blach-Overgaard, A.; et al. Climate change and land use threaten global hotspots of phylogenetic endemism for trees. Nat. Commun. 2023, 14, 6950. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Voskamp, A.; Santini, L.; Biber, M.F.; Devenish, A.J.M.; Hof, C.; Willis, S.G.; Tobias, J.A. Global impacts of climate change on avian functional diversity. Ecol. Lett. 2022, 25, 673–685. [Google Scholar] [CrossRef]
- Feng, G.; Ma, Z.; Sandel, B.; Mao, L.; Normand, S.; Ordonez, A.; Svenning, J. Species and phylogenetic endemism in angiosperm trees across the Northern Hemisphere are jointly shaped by modern climate and glacial–interglacial climate change. Glob. Ecol. Biogeogr. 2019, 28, 1393–1402. [Google Scholar] [CrossRef]
- Lancaster, L.T.; Humphreys, A.M. Global variation in the thermal tolerances of plants. Proc. Natl. Acad. Sci. USA 2020, 117, 13580–13587. [Google Scholar] [CrossRef]
- Penjor, U.; Jamtsho, R.; Sherub, S. Anthropogenic land-use change shapes bird diversity along the eastern Himalayan altitudinal gradient. J. Appl. Ecol. 2022, 59, 847–859. [Google Scholar] [CrossRef]
- Cooke, R.S.C.; Eigenbrod, F.; Bates, A.E. Projected losses of global mammal and bird ecological strategies. Nat. Commun. 2019, 10, 2279. [Google Scholar] [CrossRef]
- Hughes, E.C.; Edwards, D.P.; Bright, J.A.; Capp, E.J.; Cooney, C.R.; Varley, Z.K.; Thomas, G.H. Global biogeographic patterns of avian morphological diversity. Ecol. Lett. 2022, 25, 598–610. [Google Scholar] [CrossRef]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogues-Bravo, D.; Rasmussen, C.M.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Qian, S. Geographic patterns of taxonomic and phylogenetic β-diversity of angiosperm genera in regional floras across the world. Plant Divers. 2023, 45, 491–500. [Google Scholar] [CrossRef] [PubMed]
- State Forestry Administration. The Foruth National Survey Report on Giant Panda in China; Science Press: Beijing, China, 2021.
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species. Version 2016.3. 2016. Available online: http://www.iucnredlist.org/ (accessed on 9 June 2023).
- Li, S.; McShea, W.J.; Wang, D.; Gu, X.; Zhang, X.; Zhang, L.; Shen, X. Retreat of large carnivores across the giant panda distribution range. Nat. Ecol. Evol. 2020, 4, 1327–1331. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, X.; Wei, W.; Hong, M.; Zhou, H.; Tang, J.; Zhang, Z. Predicting range shifts of the giant pandas under future climate and land use scenarios. Ecol. Evol. 2022, 12, e9298. [Google Scholar] [CrossRef]
- Li, D.; Wang, X.; Zhao, X.; Zhou, H.; Hong, M.; Wei, W.; Han, H.; Tang, J.; Zhang, Z. The relative role of climate, land-use and spatial structure on spatial distribution pattern of species richness of large and medium mammals sympatric with giant pandas. Acta Theriol. Sin. 2021, 41, 377–387. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, S.; Che, Q.; Wei, W.; Zhao, X.; Tang, J. Impacts of climate change on the distributions and diversity of the giant panda with its sympatric mammalian species. Ecol. Indic. 2022, 144, 109452. [Google Scholar] [CrossRef]
- Tang, J.; Swaisgood, R.R.; Owen, M.A.; Zhao, X.; Wei, W.; Pilfold, N.W.; Wei, F.; Yang, X.; Gu, X.; Yang, Z.; et al. Climate change and landscape-use patterns influence recent past distribution of giant pandas. Proc. R. Soc. B 2020, 287, 20200358. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Ding, C.; Liang, D.; Xin, W.; Li, C.; Ameca, E.I.; Jiang, Z. A dataset on the morphological, life-history and ecological traits of the mammals in China. Biodivers. Sci. 2022, 30, 21520. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qian, H.U. PhyloMaker: An R package that can generate large phylogenetic trees for plants and animals. Plant Divers. 2023, 45, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Upham, N.S.; Esselstyn, J.A.; Jetz, W. Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 2019, 17, e3000494. [Google Scholar] [CrossRef] [PubMed]
- Jarzyna, M.A.; Jetz, W. Detecting the multiple facets of biodiversity. Trends Ecol. Evol. 2016, 31, 527–538. [Google Scholar] [CrossRef]
- Strona, G.; Nappo, D.; Boccacci, F.; Fattorini, S.; San-Miguel-Ayanz, J. A fast and unbiased procedure to randomize ecological binary matrices with fixed row and column totals. Nat. Commun. 2014, 5, 4114. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Peter Linder, H.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef]
- Li, X.; Yu, L.; Sohl, T.; Clinton, N.; Li, W.; Zhu, Z.; Liu, X.; Gong, P. A cellular-automata downscaling based 1 km global land use datasets (2010–2100). Sci. Bull. 2016, 61, 1651–1661. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Andrew, S.C.; Mokany, K.; Falster, D.S.; Wenk, E.; Wright, I.J.; Merow, C.; Adams, V.; Gallagher, R.V. Functional diversity of the Australian flora: Strong links to species richness and climate. J. Veg. Sci. 2021, 32, e13018. [Google Scholar] [CrossRef]
- Hu, Y.; Fan, H.; Chen, Y.; Chang, J.; Zhan, X.; Wu, H.; Zhang, B.; Wang, M.; Zhang, W.; Yang, L.; et al. Spatial patterns and conservation of genetic and phylogenetic diversity of wildlife in China. Sci. Adv. 2021, 7, eabd5725. [Google Scholar] [CrossRef]
- Xu, D.; Cong, L.; Wall, G. Tourists’ spatio-temporal behaviour and concerns in park tourism: Giant Panda National Park, Sichuan, China. Asia Pac. J. Tour. Res. 2019, 24, 924–943. [Google Scholar] [CrossRef]
- Ricotta, C. A note on functional diversity measures. Basic Appl. Ecol. 2005, 6, 479–486. [Google Scholar] [CrossRef]
- Forest, F.; Grenyer, R.; Rouget, M.; Davies, T.J.; Cowling, R.M.; Faith, D.P.; Balmford, A.; Manning, J.C.; Procheş, Ş.; Van Der Bank, M.; et al. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 2007, 445, 757–760. [Google Scholar] [CrossRef]
- Kamilar, J.M.; Beaudrot, L.; Reed, K.E. Climate and species richness predict the phylogenetic structure of African mammal communities. PLoS ONE 2015, 10, e0121808. [Google Scholar] [CrossRef]
- Wilcox, T.M.; Schwartz, M.K.; Lowe, W.H. Evolutionary community ecology: Time to think outside the (taxonomic) box. Trends Ecol. Evol. 2018, 33, 240–250. [Google Scholar] [CrossRef]
- Pellissier, V.; Barnagaud, J.Y.; Kissling, W.D.; Şekercioğlu, Ç.; Svenning, J.C. Niche packing and expansion account for species richness–productivity relationships in global bird assemblages. Glob. Ecol. Biogeogr. 2018, 27, 604–615. [Google Scholar] [CrossRef]
- Pigot, A.L.; Bregman, T.; Sheard, C.; Daly, B.; Etienne, R.S.; Tobias, J.A. Quantifying species contributions to ecosystem processes: A global assessment of functional trait and phylogenetic metrics across avian seed-dispersal networks. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161597. [Google Scholar] [CrossRef]
- Bai, W.; Huang, Q.; Zhang, J.; Stabach, J.; Huang, J.; Yang, H.; Songer, M.; Connor, T.; Liu, J.; Zhou, S.; et al. Microhabitat selection by giant pandas. Biol. Conserv. 2020, 247, 108615. [Google Scholar] [CrossRef]
- Wei, W.; Swaisgood, R.R.; Dai, Q.; Yang, Z.; Yuan, S.; Owen, M.A.; Pilfold, N.W.; Yang, X.; Gu, X.; Zhou, H.; et al. Giant panda distributional and habitat-use shifts in a changing landscape. Conserv. Lett. 2018, 11, e12575. [Google Scholar] [CrossRef]
- Wei, W.; Swaisgood, R.R.; Pilfold, N.W.; Owen, M.A.; Dai, Q.; Wei, F.; Han, H.; Yang, Z.; Yang, X.; Gu, X.; et al. Assessing the effectiveness of China’s panda protection system. Curr. Biol. 2020, 30, 1280–1286.e2. [Google Scholar] [CrossRef]
- Sobral, F.L.; Jardim, L.; Lemes, P.; Machado, N.; Loyola, R.; Cianciaruso, M.V. Spatial conservation priorities for top predators reveal mismatches among taxonomic, phylogenetic and functional diversity. Nat. Conserv. 2014, 12, 150–155. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Indices | Explanatory Variables | SE | t | Pr (>|t|) | |
|---|---|---|---|---|---|
| Species richness | Intercept | 51.071 | 0.559 | 91.406 | 1.00 × 10−90 |
| SD of elevation | 0.712 | 0.282 | 2.524 | 0.012 | |
| Proportion of UGS | −0.019 | 0.187 | −0.099 | 0.921 | |
| Proportion of forest | 2.895 | 0.404 | 7.162 | 1.18 × 10−12 | |
| Proportion of cropland | −0.167 | 0.449 | −0.373 | 0.709 | |
| AP | −1.799 | 0.642 | −2.804 | 0.005 | |
| MAT | 9.282 | 0.436 | 21.276 | 2.89 × 10−89 | |
| LGM AP anomaly | −2.984 | 0.508 | −5.875 | 5.07 × 10−9 | |
| LGM MAT anomaly | 2.080 | 0.194 | 10.695 | 7.01 × 10−26 | |
| SES functional diversity | Intercept | −0.124 | 0.086 | −1.443 | 0.149 |
| SD of elevation | 0.015 | 0.026 | 0.587 | 0.557 | |
| Proportion of UGS | 0.011 | 0.016 | 0.654 | 0.513 | |
| Proportion of forest | −0.001 | 0.037 | −0.032 | 0.975 | |
| Proportion of cropland | −0.100 | 0.042 | −2.414 | 0.016 | |
| AP | −0.230 | 0.075 | −3.087 | 0.002 | |
| MAT | −0.125 | 0.044 | −2.813 | 0.005 | |
| LGM AP anomaly | −0.129 | 0.055 | −2.354 | 0.019 | |
| LGM MAT anomaly | −0.013 | 0.018 | −0.742 | 0.458 | |
| SES phylogenetic diversity | Intercept | −0.263 | 0.06 | −4.405 | 1.12 × 10−5 |
| SD of elevation | 0.038 | 0.028 | 1.351 | 0.177 | |
| Proportion of UGS | 0.047 | 0.018 | 2.566 | 0.010 | |
| Proportion of forest | 0.118 | 0.04 | 2.933 | 0.003 | |
| Proportion of cropland | −0.09 | 0.045 | −2.014 | 0.044 | |
| AP | −0.278 | 0.066 | −4.205 | 2.75 × 10−5 | |
| MAT | −0.288 | 0.044 | −6.573 | 6.52 × 10−5 | |
| LGM AP anomaly | −0.143 | 0.052 | −2.772 | 0.006 | |
| LGM MAT anomaly | −0.048 | 0.019 | −2.489 | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che, Q.; Li, C.; Zhao, X.; Zhang, J.; Tang, J.; Zhou, C. Effects of Climate and Land Use on Different Facets of Mammal Diversity in Giant Panda Range. Animals 2025, 15, 630. https://doi.org/10.3390/ani15050630
Che Q, Li C, Zhao X, Zhang J, Tang J, Zhou C. Effects of Climate and Land Use on Different Facets of Mammal Diversity in Giant Panda Range. Animals. 2025; 15(5):630. https://doi.org/10.3390/ani15050630
Chicago/Turabian StyleChe, Qibing, Chunxiao Li, Xuzhe Zhao, Jindong Zhang, Junfeng Tang, and Caiquan Zhou. 2025. "Effects of Climate and Land Use on Different Facets of Mammal Diversity in Giant Panda Range" Animals 15, no. 5: 630. https://doi.org/10.3390/ani15050630
APA StyleChe, Q., Li, C., Zhao, X., Zhang, J., Tang, J., & Zhou, C. (2025). Effects of Climate and Land Use on Different Facets of Mammal Diversity in Giant Panda Range. Animals, 15(5), 630. https://doi.org/10.3390/ani15050630