

Assessing Heat Resistance and Selecting Heat-Resistant Individuals of Largemouth Bass (Micropterus salmoides) with Tiered Thermal Exposure
 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Maintenance
2.2. Determination of Critical Thermal Maximum
2.3. Static Heat Exposure
2.4. Tiered Heat Exposure and Calculation of Lethal Cumulative Temperature (LCT)
2.5. Identification and Characterization of Heat-Sensitive LMB Individuals
2.5.1. Histological Analysis and TUNEL Staining
2.5.2. Total RNA Isolation and Quantitative PCR
2.6. Application of the Tiered Exposure Method
2.6.1. Comparing Heat Resistance of LMB Juveniles and Adults
2.6.2. Selecting Heat-Resistant Individuals from the LMB Population
2.7. Statistics
3. Results
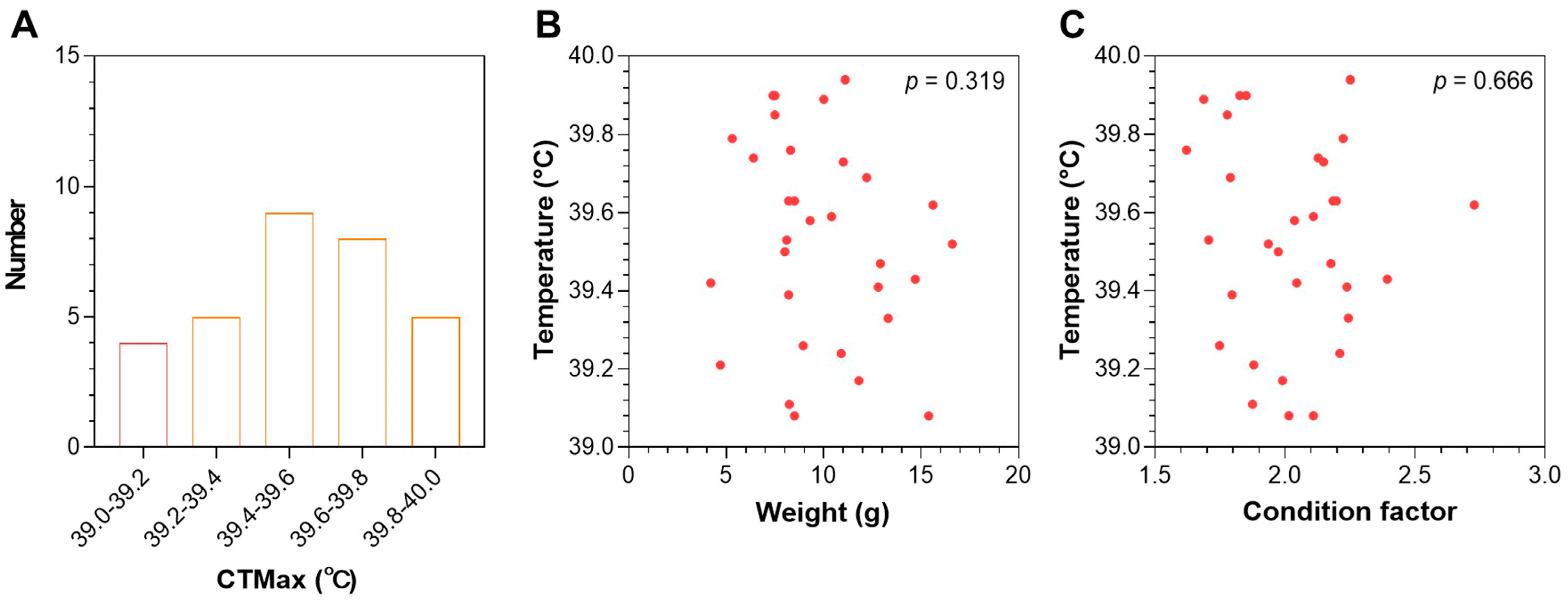
3.1. CTmax of Largemouth Bass Juveniles
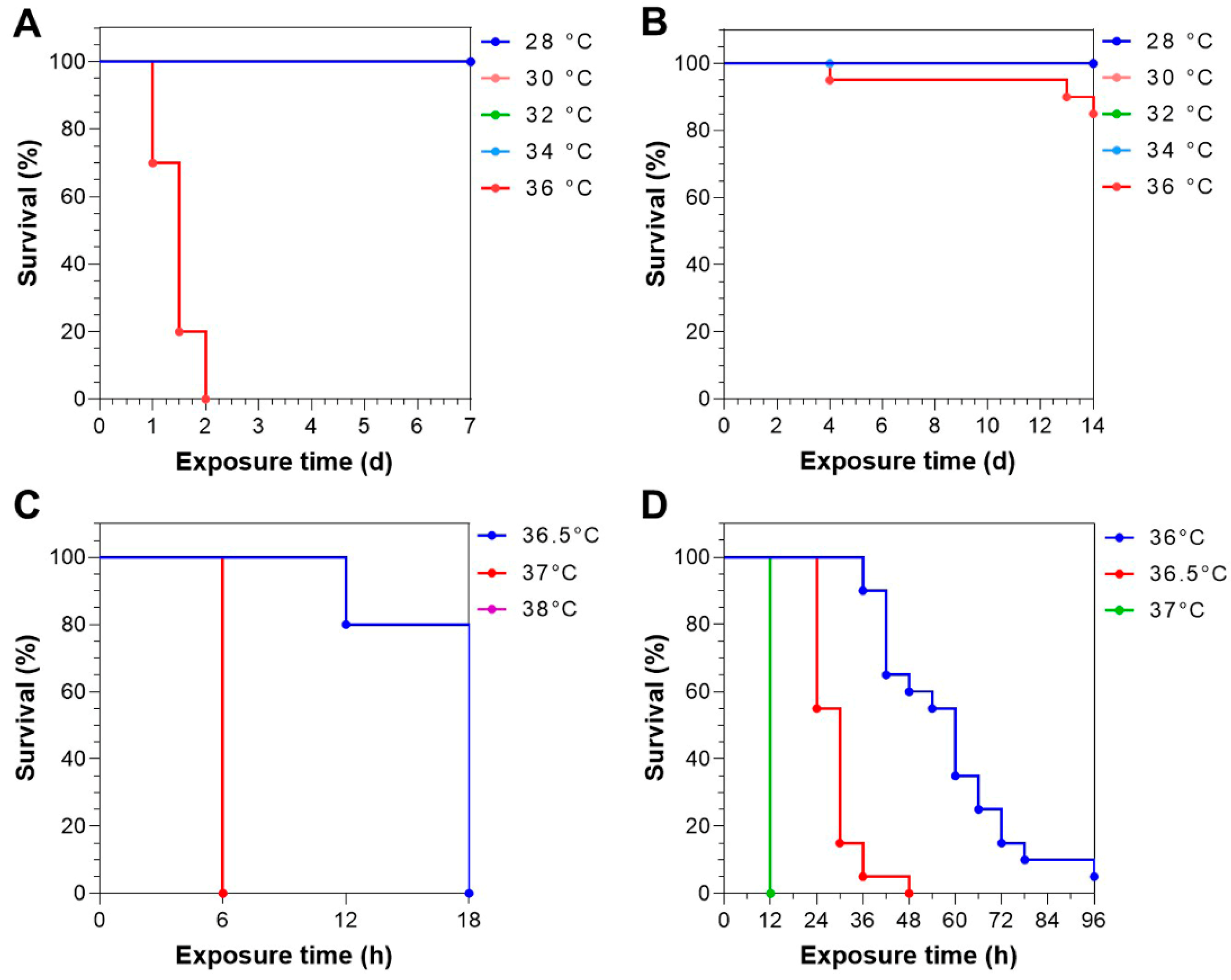
3.2. Survival Times of Largemouth Bass upon Static Heat Exposure
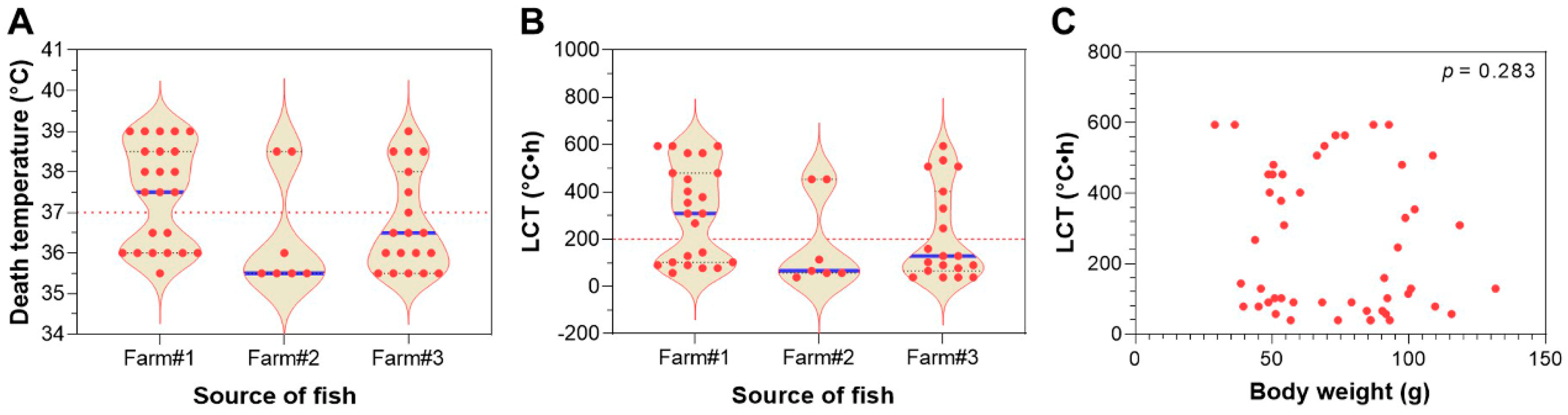
3.3. Lethal Cumulative Temperature of Largemouth Bass upon Tiered Heat Exposure
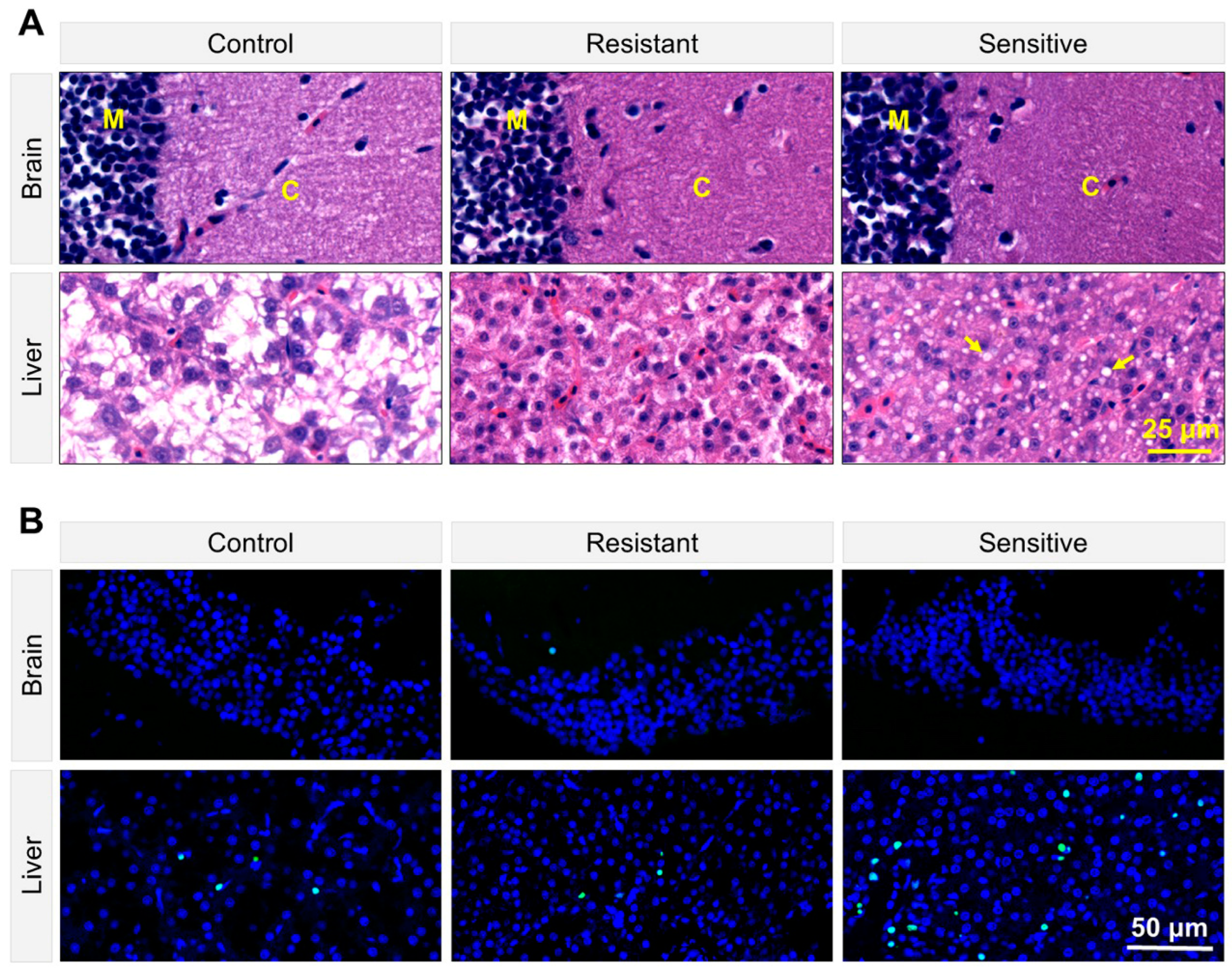
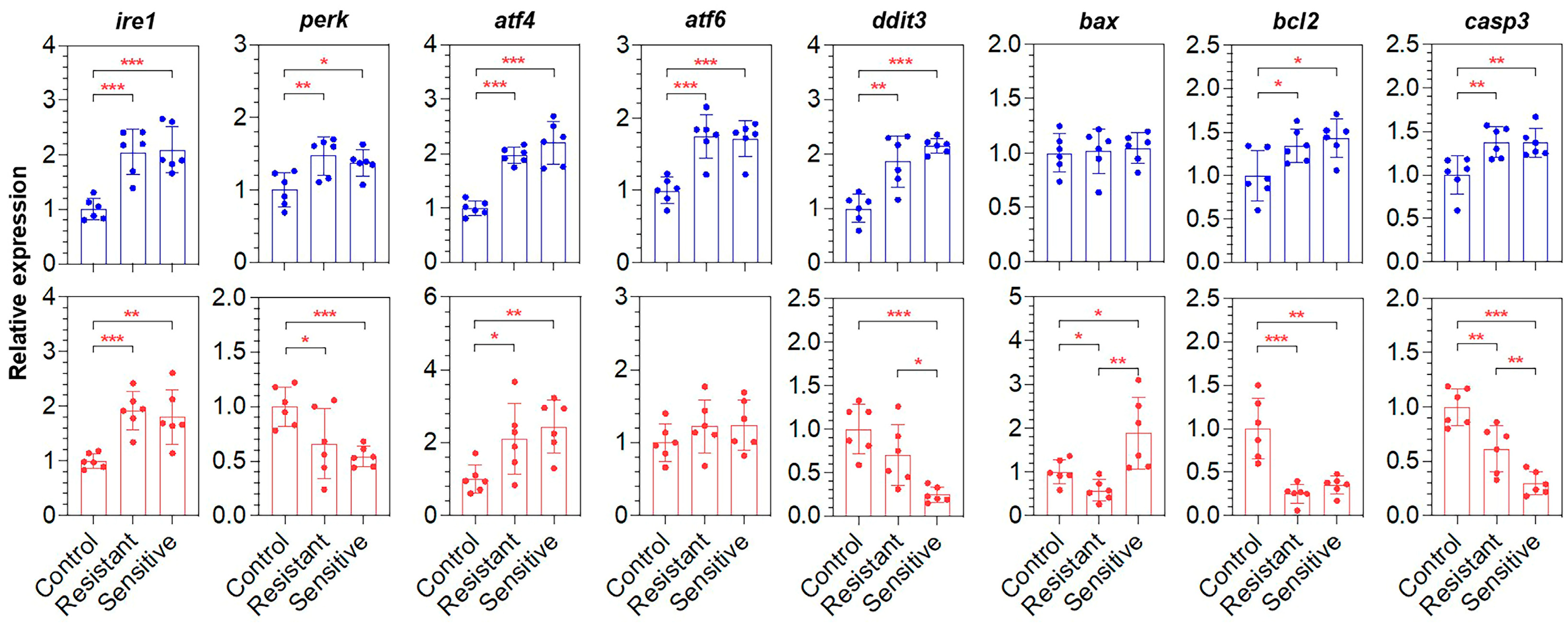
3.4. Heat-Sensitive Fish Demonstrated Tissue Damage and Apoptosis in the Liver
3.5. Expressions of the ER Stress-Response- and Apoptosis-Associated Genes
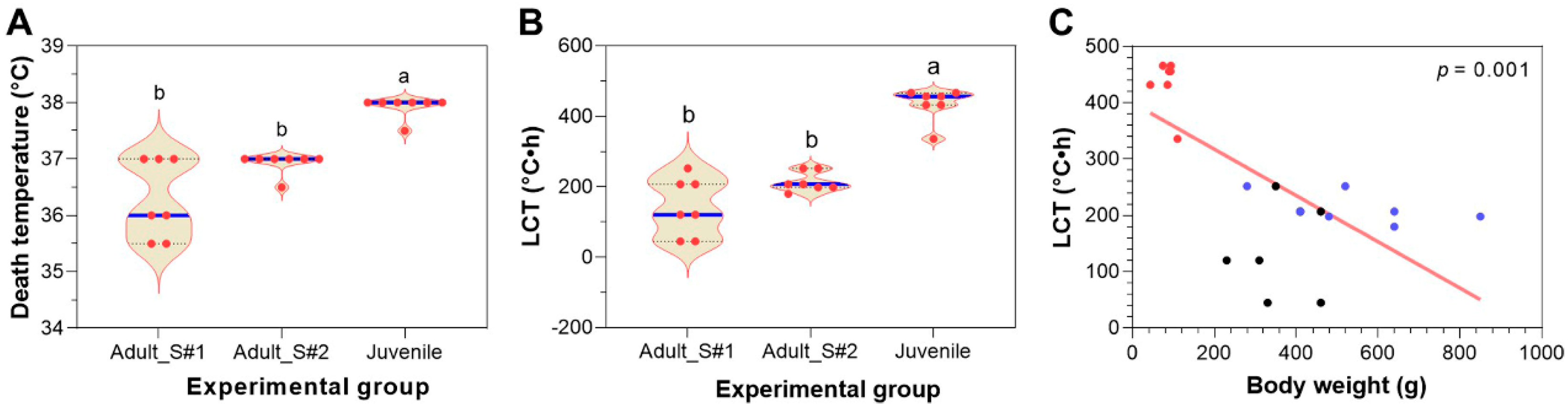
3.6. Comparing Heat Resistance of Adult and Juvenile Largemouth Bass
3.7. Selection of Heat-Resistant Largemouth Bass from the Population
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2022. [Google Scholar]
- Reid, G.K.; Gurney-Smith, H.J.; Flaherty, M.; Garber, A.F.; Forster, I.; Brewer-Dalton, K.; Knowler, D.; Marcogliese, D.J.; Chopin, T.; Moccia, R.D.; et al. Climate change and aquaculture considering adaptation potential. Aquac. Environ. Interact. 2019, 11, 603–624. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.S.; McKay, N.P. Technical Note: Past and future warming–direct comparison on multi-century timescales. Clim. Past 2022, 18, 911–917. [Google Scholar] [CrossRef]
- Barbarossa, V.; Bosmans, J.; Wanders, N.; King, H.; Bierkens, M.F.P.; Huijbregts, M.A.J.; Schipper, A.M. Threats of global warming to the world’s freshwater fishes. Nat. Commun. 2021, 12, 1701. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef]
- Huang, M.; Ding, L.; Wang, J.; Ding, C.; Tao, J. The impacts of climate change on fish growth: A summary of conducted studies and current knowledge. Ecol. Indic. 2021, 121, 106976. [Google Scholar] [CrossRef]
- Lema, S.C.; Luckenbach, J.A.; Yamamoto, Y.; Housh, M.J. Fish reproduction in a warming world: Vulnerable points in hormone regulation from sex determination to spawning. Philos. Trans. R. Soc. B Biol. Sci. 2024, 379, 20220516. [Google Scholar] [CrossRef] [PubMed]
- González-Ferreras, A.M.; Barquín, J.; Blyth, P.S.; Hawksley, J.; Kinsella, H.; Lauridsen, R.; Morris, O.F.; Peñas, F.J.; Thomas, G.E.; Woodward, G. Chronic exposure to environmental temperature attenuates the thermal sensitivity of salmonids. Nat. Commun. 2023, 14, 8309. [Google Scholar] [CrossRef] [PubMed]
- Little, A.G.; Loughland, I.; Seebacher, F. What do warming waters mean for fish physiology and fisheries? J. Fish Biol. 2020, 97, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Hussein, G.H.G.; Chen, M.; Qi, P.P.; Cui, Q.K.; Yu, Y.; Hu, W.H.; Tian, Y.; Fan, Q.X.; Gao, Z.X.; Feng, M.W.; et al. Aquaculture industry development, annual price analysis and out-of-season spawning in largemouth bass Micropterus salmoides. Aquaculture 2020, 519, 734901. [Google Scholar] [CrossRef]
- Fantini, L.E.; Smith, M.A.; Jones, M.; Roy, L.A.; Lochmann, R.; Kelly, A.M. Growth parameters in northern largemouth bass Micropterus salmoides salmoides raised near their upper thermal tolerance for 28 days. Aquacult. Rep. 2021, 21, 100845. [Google Scholar] [CrossRef]
- Zhao, X.; Li, L.; Li, C.; Liu, E.; Zhu, H.; Ling, Q. Heat stress-induced endoplasmic reticulum stress promotes liver apoptosis in largemouth bass (Micropterus salmoides). Aquaculture 2022, 546, 737401. [Google Scholar] [CrossRef]
- Yang, C.; Dong, J.J.; Sun, C.F.; Li, W.H.; Tian, Y.Y.; Liu, Z.G.; Gao, F.Y.; Ye, X. Exposure to heat stress causes downregulation of immune response genes and weakens the disease resistance of Micropterus salmoides. Comp. Biochem. Phys. D 2022, 43, 101011. [Google Scholar] [CrossRef] [PubMed]
- Xv, Z.; Chen, S.; Song, G.; Hu, H.; Lin, S.; Long, Y. Biochemical, histological and transcriptomic analyses for the immunological organs provide insights into heat stress-induced disease susceptibility in Largemouth Bass. Sci. Total Environ. 2024, 912, 168758. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Mao, W.J.; Lin, Z.J.; Ling, Q.F. Heat stress induced hepatocyte apoptosis in largemouth bass via IRE1α/TRAF2/ASK1/JNK pathway. J. Oceanol. Limnol. 2024, 42, 988–1000. [Google Scholar] [CrossRef]
- Liu, E.G.; Zhao, X.Q.; Li, C.J.; Wang, Y.F.; Li, L.L.; Zhu, H.; Ling, Q.F. Effects of acute heat stress on liver damage, apoptosis and inflammation of pikeperch (Sander lucioperca). J. Therm. Biol. 2022, 106, 103251. [Google Scholar] [CrossRef]
- Zhou, F.; Qi, M.; Li, J.; Huang, Y.; Chen, X.; Liu, W.; Yao, G.; Meng, Q.; Zheng, T.; Wang, Z.; et al. Comparative Transcriptomic Analysis of Largemouth Bass (Micropterus salmoides) Livers Reveals Response Mechanisms to High Temperatures. Genes 2023, 14, 2096. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.X.; Wang, Y.Z.; Wang, Z.L.; Luo, T.M.; Huang, J.; Shao, J. Analysis of Differential Alternative Splicing in Largemouth Bass After High Temperature Exposure. Animals 2024, 14, 3005. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.D.; Long, X.R.; Zhang, X.H.; Dong, X.H.; Wang, Z.L.; Jiang, H.B.; An, M.; Chen, J.F.; Gan, L. Identification and characterization of long non-coding RNAs in intestinal immune regulation of largemouth bass, Micropterus salmoides, under acute heat stress. Comp. Biochem. Phys. D 2023, 48, 101132. [Google Scholar] [CrossRef]
- Yu, J.J.; Zhong, D.B.; Li, S.; Zhang, Z.H.; Mo, H.L.; Wang, L.X. Acute temperature stresses trigger liver transcriptome and microbial community remodeling in largemouth bass. Aquaculture 2023, 573, 739573. [Google Scholar] [CrossRef]
- Cowles, R.B.; Bogert, C.M. A preliminary study of the thermal requirements of desert reptiles. Bull. Am. Mus. Nat. Hist. 1944, 83, 263–296. [Google Scholar]
- Fry, F.; Brett, J.; Clawson, G. Lethal limits of temperature for young goldfish. Rev. Can. Biol. 1942, 1, 50–56. [Google Scholar]
- Cowan, Z.L.; Andreassen, A.H.; De Bonville, J.; Green, L.; Binning, S.A.; Silva-Garay, L.; Jutfelt, F.; Sundin, J. A novel method for measuring acute thermal tolerance in fish embryos. Conserv. Physiol. 2023, 11, coad061. [Google Scholar] [CrossRef] [PubMed]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: History and critique. Can. J. Zool. 1997, 75, 1561–1574. [Google Scholar] [CrossRef]
- Cereja, R. Critical thermal maxima in aquatic ectotherms. Ecol. Indic. 2020, 119, 106856. [Google Scholar] [CrossRef]
- Morgan, R.; Finnøen, M.H.; Jutfelt, F. CTmax is repeatable and doesn’t reduce growth in zebrafish. Sci. Rep. 2018, 8, 7099. [Google Scholar] [CrossRef]
- Yokoyama, S.; Koshio, S.; Takakura, N.; Oshida, K.; Ishikawa, M.; Gallardo-Cigarroa, F.J.; Teshima, S.-I. Dietary bovine lactoferrin enhances tolerance to high temperature stress in Japanese flounder Paralichthys olivaceus. Aquaculture 2005, 249, 367–373. [Google Scholar] [CrossRef]
- Molony, B.W.; Church, A.R.; Maguire, G.B. A comparison of the heat tolerance and growth of a selected and non-selected line of rainbow trout, Oncorhynchus mykiss, in Western Australia. Aquaculture 2004, 241, 655–665. [Google Scholar] [CrossRef]
- Liu, R.; Liu, R.; Song, G.; Li, Q.; Cui, Z.; Long, Y. Mitochondria Dysfunction and Cell Apoptosis Limit Resistance of Nile Tilapia (Oreochromis niloticus) to Lethal Cold Stress. Animals 2022, 12, 2382. [Google Scholar] [CrossRef]
- Becker, C.D.; Genoway, R.G. Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environ. Biol. Fishes 1979, 4, 245–256. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.; Lowe, S.S.; Kaminski, C.; Whitt, G.S.; Philipp, D.P. Critical and Chronic Thermal Maxima of Northern and Florida Largemouth Bass and Their Reciprocal F1 and F2 Hybrids. Trans. Am. Fish. Soc. 1987, 116, 856–863. [Google Scholar] [CrossRef]
- Currie, R.J.; Bennett, W.A.; Beitinger, T.L. Critical thermal minima and maxima of three freshwater game-fish species acclimated to constant temperatures. Environ. Biol. Fishes 1998, 51, 187–200. [Google Scholar] [CrossRef]
- Yanar, M.; Erdoğan, E.; Kumlu, M. Thermal tolerance of thirteen popular ornamental fish Species. Aquaculture 2019, 501, 382–386. [Google Scholar] [CrossRef]
- Li, T.; Mo, K.L.; Wang, J.; Chen, Q.W.; Zhang, J.Y.; Zeng, C.J.; Zhang, H.; Yang, P.S. Mismatch between critical and accumulated temperature following river damming impacts fish spawning. Sci. Total Environ. 2021, 756, 144052. [Google Scholar] [CrossRef] [PubMed]
- Tsoukali, S.; Visser, A.W.; MacKenzie, B.R. Functional responses of North Atlantic fish eggs to increasing temperature. Mar. Ecol. Prog. Ser. 2016, 555, 151–165. [Google Scholar] [CrossRef]
- Bevelhimer, M.; Bennett, W. Assessing cumulative thermal stress in fish during chronic intermittent exposure to high temperatures. Environ. Sci. Policy 2000, 3, 211–216. [Google Scholar] [CrossRef]
- Roychowdhury, P.; Aftabuddin, M.; Pati, M.K. Thermal stress-induced oxidative damages in the liver and associated death in fish. Fish Physiol. Biochem. 2021, 47, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Lindén, M.; Österlund, T.; Vannas, C.; Andersson, L.; Escobar, M.; Ståhlberg, A.; Åman, P. Identification of genomic binding sites and direct target genes for the transcription factor DDIT3/CHOP. Exp. Cell Res. 2023, 422, 113418. [Google Scholar] [CrossRef]
- Li, M.Y.; Thorne, R.F.; Shi, R.H.; Zhang, X.D.; Li, J.M.; Li, J.T.; Zhang, Q.Y.; Wu, M.A.; Liu, L.X. DDIT3 Directs a Dual Mechanism to Balance Glycolysis and Oxidative Phosphorylation during Glutamine Deprivation. Adv. Sci. 2021, 8, 2003732. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Xu, X.X.; Dong, X.F.; Yang, B.N.; Dong, W.; Luo, Y.; Liu, X.Y.; Wu, Y.R.; Wang, J.W. DDIT3/CHOP promotes autophagy in chondrocytes via SIRT1-AKT pathway. Bba-Mol. Cell Res. 2021, 1868, 119074. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.; Peltzer, N.; Walicki, J.; Yang, J.Y.; Dubuis, G.; Gardiol, N.; Held, W.; Bigliardi, P.; Marsland, B.; Liaudet, L.; et al. Caspase-3 Protects Stressed Organs against Cell Death. Mol. Cell Biol. 2012, 32, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Y.; Liu, T.; Jiang, S.; Li, R.; Ge, Y.; Wang, Y.; Tu, H.; Huang, J.; Zhu, X.; et al. Genome wide association analysis reveals the genetic sites of heat resistance in largemouth bass (Micropterus salmoides). Aquaculture 2024, 590, 741025. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Type | Experiment# | Experimental Group/Fish Source | Development Stage | Fish Number | Body Weight (g) | Standard Length (cm) | Rearing Temperature (°C) |
|---|---|---|---|---|---|---|---|
| CTmax | Experiment#1 | NA | Juvenile | 31 | 9.87 ± 3.14 | 7.78 ± 0.78 | 26–28 |
| Experiment#2 | NA | Juvenile | 19 | 15.65 ± 3.88 | 8.77 ± 0.74 | 26–28 | |
| Static exposure | Experiment#1 | 28, 30, 32, 34, 36 °C | Juvenile | 10 fish per group | 17.58 ± 5.00 | 8.38 ± 1.22 | 23–25 |
| Experiment#2 | 28, 30, 32, 34, 36 °C | Juvenile | 20 fish per group | 100.62 ± 23.31 | 15.89 ± 1.20 | 24–26 | |
| Experiment#3 | 36.5, 37, 38 °C | Juvenile | 10 fish per group | 96.01 ± 17.53 | 15.54 ± 0.97 | 24–27 | |
| Experiment#4 | 36, 36.5, 37 °C | Juvenile | 20 fish per group | 45.12 ± 10.47 | 12.32 ± 0.92 | 26–28 | |
| Tiered exposure | Experiment#1 | Farm#1 | Juvenile | 23 | 59.89 ± 23.25 | 13.59 ± 1.51 | 26–28 |
| Farm#2 | Juvenile | 7 | 78.95 ± 25.55 | 14.83 ± 1.59 | 26–28 | ||
| Farm#3 | Juvenile | 20 | 87.83 ± 19.18 | 14.94 ± 1.16 | 26–28 | ||
| Experiment#2 | Tank#1, Tank#2, Tank#3 | Juvenile | 33, 47, 48 | 23.66 ± 6.98 | 9.80 ± 0.81 | 25–28 | |
| Experiment#3 | Adult_S#1 | Adult | 7 | 364.29 ± 84.43 | 24.40 ± 1.91 | 26–28 | |
| Adult_S#2 | Adult | 7 | 545.71 ± 184.56 | 27.19 ± 3.06 | 26–28 | ||
| Juvenile | Juvenile | 7 | 84.06 ± 20.95 | 14.97 ± 1.42 | 26–28 | ||
| Experiment#4 | Tank#1, Tank#2, Tank#3, Tank#4 | Juvenile | 82, 70, 56, 55 | 22.69 ± 6.89 | 9.96 ± 2.00 | 25–28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Qiao, H.; Xv, Z.; Song, G.; Liu, S.; Luo, C.; Long, Y.; Lin, S. Assessing Heat Resistance and Selecting Heat-Resistant Individuals of Largemouth Bass (Micropterus salmoides) with Tiered Thermal Exposure. Animals 2025, 15, 128. https://doi.org/10.3390/ani15020128
Chen H, Qiao H, Xv Z, Song G, Liu S, Luo C, Long Y, Lin S. Assessing Heat Resistance and Selecting Heat-Resistant Individuals of Largemouth Bass (Micropterus salmoides) with Tiered Thermal Exposure. Animals. 2025; 15(2):128. https://doi.org/10.3390/ani15020128
Chicago/Turabian StyleChen, Haijie, Hui Qiao, Zhicheng Xv, Guili Song, Shuning Liu, Cheng Luo, Yong Long, and Shimei Lin. 2025. "Assessing Heat Resistance and Selecting Heat-Resistant Individuals of Largemouth Bass (Micropterus salmoides) with Tiered Thermal Exposure" Animals 15, no. 2: 128. https://doi.org/10.3390/ani15020128
APA StyleChen, H., Qiao, H., Xv, Z., Song, G., Liu, S., Luo, C., Long, Y., & Lin, S. (2025). Assessing Heat Resistance and Selecting Heat-Resistant Individuals of Largemouth Bass (Micropterus salmoides) with Tiered Thermal Exposure. Animals, 15(2), 128. https://doi.org/10.3390/ani15020128