

Impact of Heat Stress on Ovarian Function and circRNA Expression in Hu Sheep

 , , , and
, , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Site and Environmental Conditions
2.3. Animals and Experimental Diets
2.4. Sample Collection
2.5. Serum Biochemical Level Analysis
2.6. Histological Assay
2.7. TUNEL Assay
2.8. RNA Extraction, Small RNA Library Construction, and Sequencing
2.9. Statistical Analyses
2.10. RT-qPCR Validation
2.11. Statistical Analysis
3. Results
3.1. The Effects of HS on the Physiological Characteristics of Hu Sheep
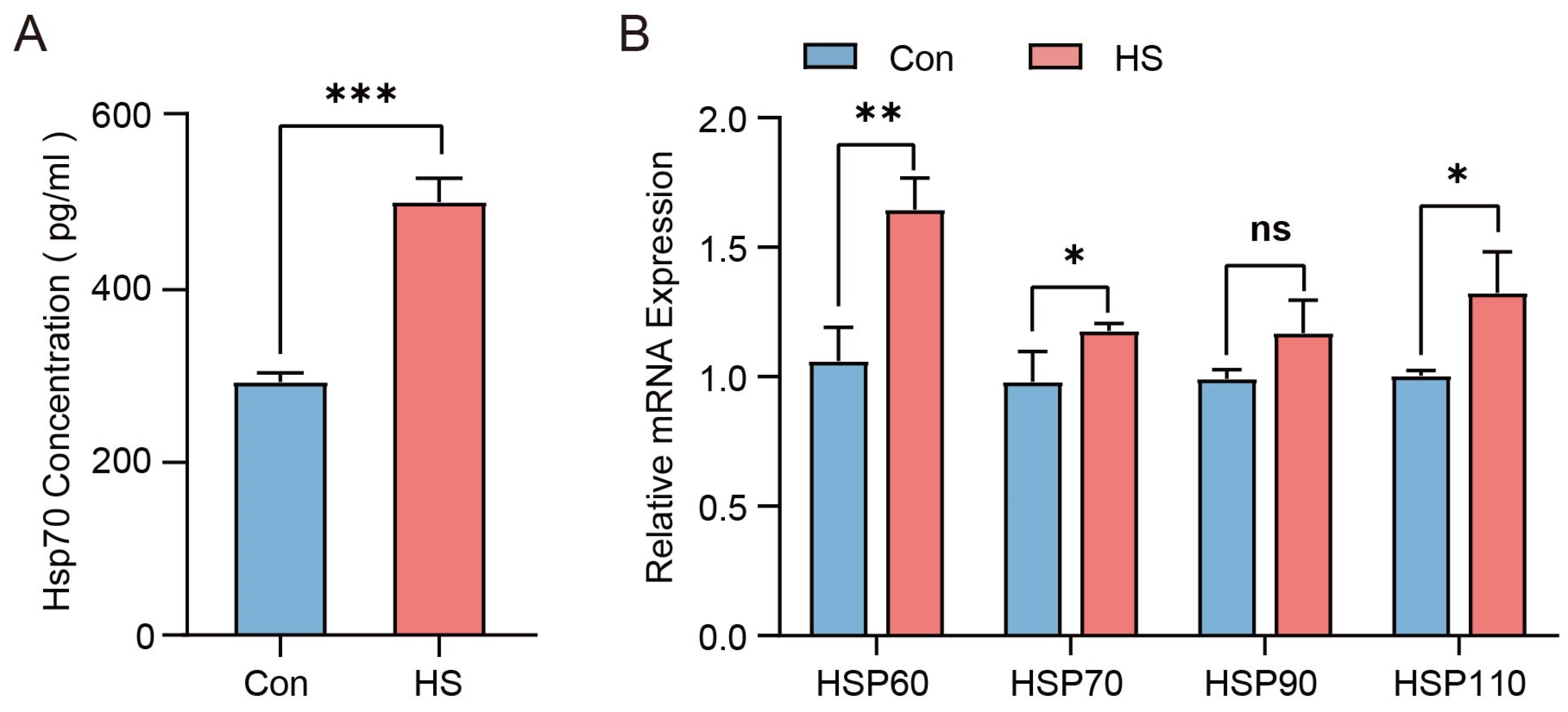
3.2. Effects of HS on Antioxidant Capacity in Hu Sheep
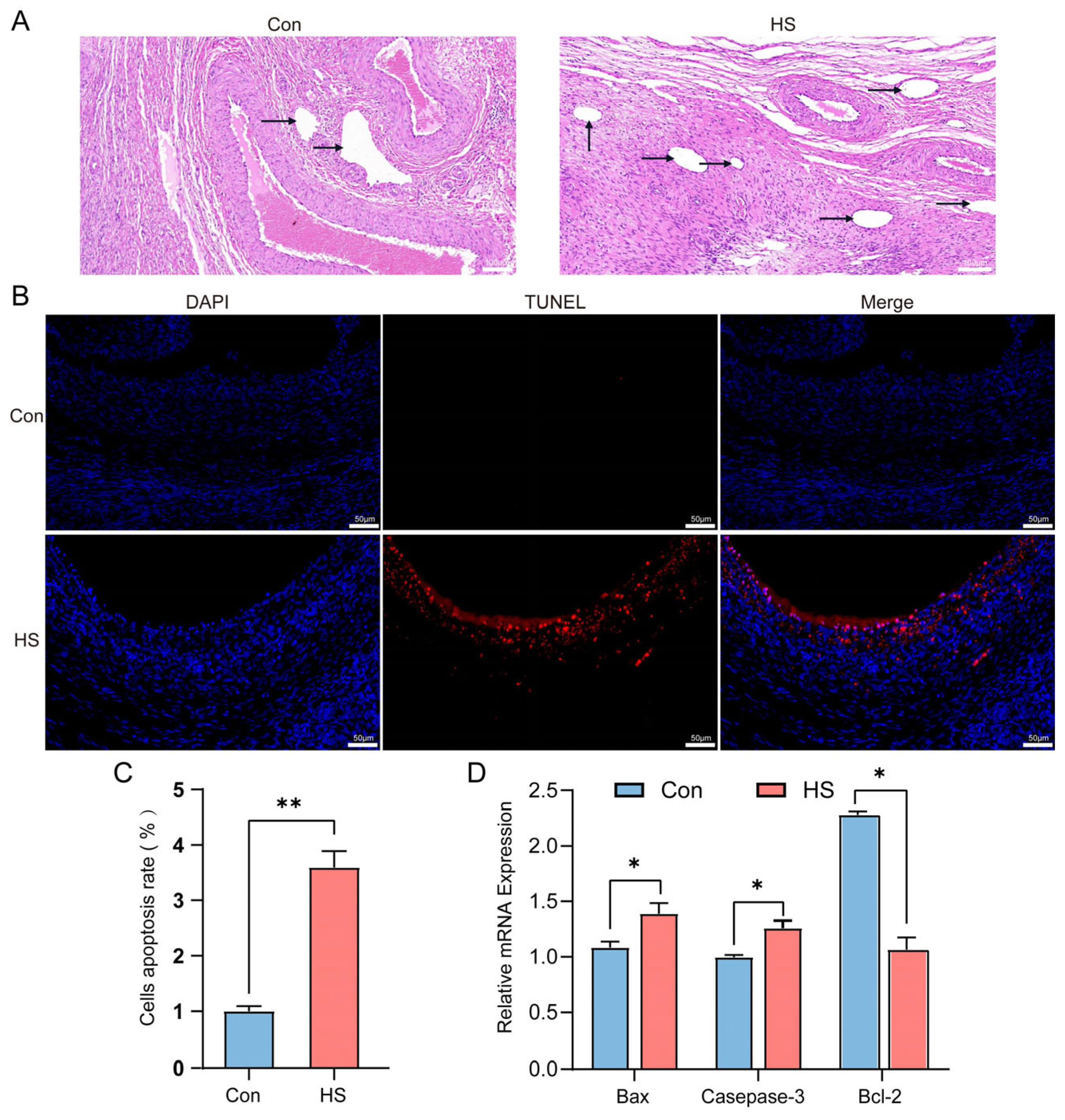
3.3. Effects of HS on Ovarian Tissue Morphology and Cell Apoptosis in Hu Sheep
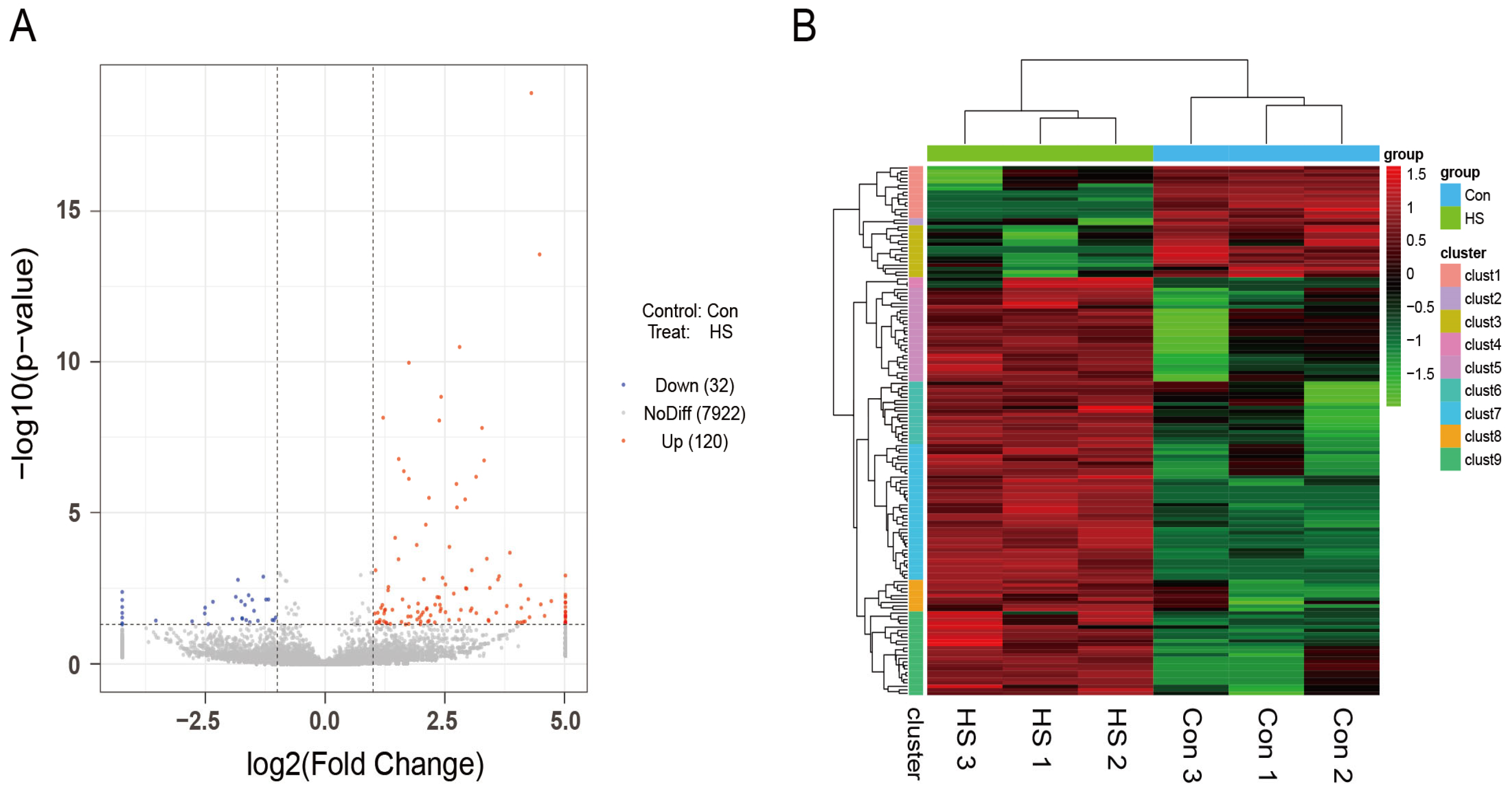
3.4. Identification and Characterization of circRNAs in Ovarian Tissues of Hu Sheep Under HS
3.5. Differential Expression Patterns of circRNAs in Hu Sheep Ovaries Under HS
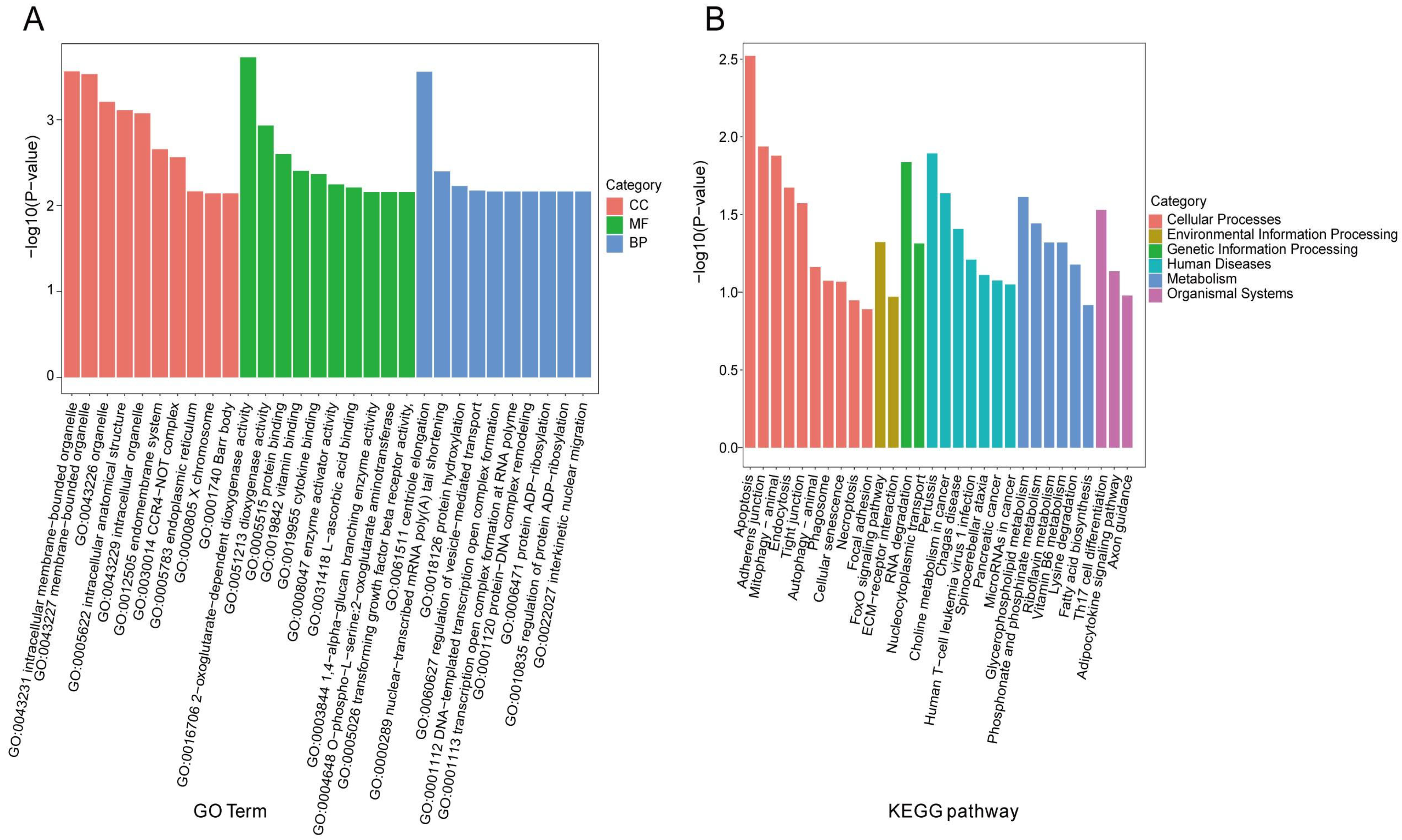
3.6. Functional Enrichment Analysis of Source Genes of Differentially Expressed circRNAs in Hu Sheep Ovaries Under HS
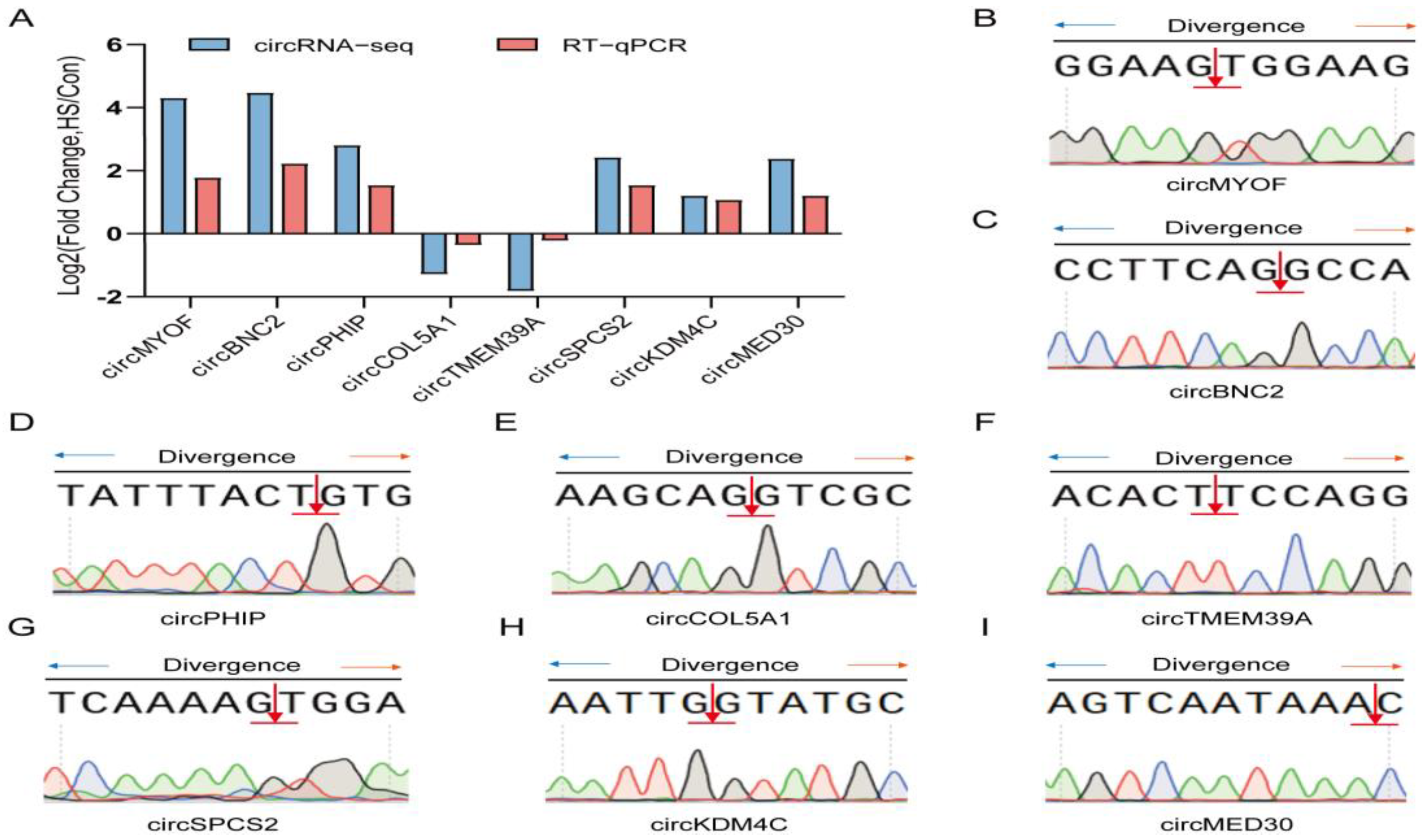
3.7. RT-qPCR Validation of circRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Wettere, W.; Culley, S.; Swinbourne, A.; Leu, S.; Lee, S.; Weaver, A.; Kelly, J.; Walker, S.; Kleemann, D.; Thomas, D.; et al. Heat stress from current and predicted increases in temperature impairs lambing rates and birth weights in the Australian sheep flock. Nat. Food 2024, 5, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Arero, G.; Ozmen, O. Effects of heat stress on reproduction and gene expression in sheep. Anim. Reprod. 2025, 22, e20240067. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Dunshea, F.; Warner, R.; DiGiacomo, K.; Osei-Amponsah, R.; Chauhan, S. Impacts of heat stress on meat quality and strategies for amelioration: A review. Int. J. Biometeorol. 2020, 64, 1613–1628. [Google Scholar] [CrossRef] [PubMed]
- van Wettere, W.H.E.J.; Kind, K.L.; Gatford, K.L.; Swinbourne, A.M.; Leu, S.T.; Hayman, P.T.; Kelly, J.M.; Weaver, A.C.; Kleemann, D.O.; Walker, S.K. Review of the impact of heat stress on reproductive performance of sheep. J. Anim. Sci. Biotechnol. 2021, 12, 26. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Wei, Y.; Zou, J.; Yang, B.; Wang, Q.; Lu, J.; Lu, J.; Zheng, Z.; Huang, Y.; et al. Effect of heat stress on growth performance, carcase characteristics, meat quality and rumen-muscle axis of Hu sheep. Ital. J. Anim. Sci. 2024, 23, 87–100. [Google Scholar] [CrossRef]
- Karthik, D.; Suresh, J.; Reddy, Y.; Sharma, G.; Ramana, J.; Gangaraju, G.; Reddy, P.; Reddy, Y.; Yasaswini, D.; Adegbeye, M.; et al. Adaptive profiles of Nellore sheep with reference to farming system and season: Physiological, hemato-biochemical, hormonal, oxidative-enzymatic and reproductive standpoint. Heliyon 2021, 7, e07117. [Google Scholar] [CrossRef]
- De, K.; Kumar, D.; Balaganur, K.; Naqvi, S. Effect of environmental factors on estrus synchronization and artificial insemination success in farmers flock in sheep under semi-arid tropical region. Reprod. Domest. Anim. 2020, 55, 777–784. [Google Scholar] [CrossRef]
- Mossa, F.; Evans, A. Review: The ovarian follicular reserve—Implications for fertility in ruminants. Animal 2023, 17, 100744. [Google Scholar] [CrossRef]
- Li, Y.; Li, F.; Shu, J.; Meng, C.; Zhang, J.; Zhang, J.; Qian, Y.; Wang, H.; Ding, Q.; Cao, S. Acute heat stress regulates estradiol synthesis in ovine ovarian granulosa cells through the SREBPs/MVK-LHR pathway. Anim. Reprod. Sci. 2025, 272, 107649. [Google Scholar] [CrossRef]
- Dalgleish, M. Where the Global Sheep Flock Can Be Found and Where It’s Going. In Agricultural Outlook; FAO: Rome, Italy, 2024. [Google Scholar]
- IWTO. World Sheep Numbers and Wool Production; IWTO: Brussels, Belgium, 2022. [Google Scholar]
- Lingying, K.; Yaojing, Y.; Jianye, L.; Bohui, Y.; Bowen, C.; Jianbin, L.; Zengkui, L. Transcriptomics and metabolomics reveal improved performance of Hu sheep on hybridization with Southdown sheep. Food Res. Int. 2023, 173, 113240. [Google Scholar] [CrossRef]
- Wu, X.; Yang, X.; Dai, Y.; Zhao, Z.; Zhu, J.; Guo, H.; Yang, R. Single-cell sequencing to multi-omics: Technologies and applications. Biomark. Res. 2024, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Haire, A.; Bai, J.; Zhao, X.; Song, Y.; Zhao, G.; Dilixiati, A.; Li, J.; Sun, W.Q.; Wan, P.; Fu, X.; et al. Identifying the heat resistant genes by multi-tissue transcriptome sequencing analysis in Turpan Black sheep. Theriogenology 2021, 179, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-D.; Wang, L.-W.; Fu, S.-Y.; E, R.-G.; Ren, X.-Q.; Sun, H.; Liu, F.; Wang, B.; An, J.-H.; Zhao, M.-R.; et al. Heat Tolerance Differences Between Hu Sheep and Hu Crossbred Sheep in Microbial Community Structure and Metabolism. Metabolites 2025, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; E, G.; Zhang, M.; Zhang, Y.; Bai, T.; Pu, X.; Liu, J.; Guo, X.; Sarker, S.; Cheng, L. No rumen fermentation profiles and associated microbial diversities difference were found between Hu sheep and Karakul sheep fed a cottonseed hull diet. Microbiome 2025, 13, 22. [Google Scholar] [CrossRef]
- Zhao, L.; Yuan, L.; Li, F.; Zhang, X.; Tian, H.; Ma, Z.; Zhang, D.; Zhang, Y.; Zhao, Y.; Huang, K.; et al. Whole-genome resequencing of Hu sheep identifies candidate genes associated with agronomic traits. J. Genet. Genom. 2024, 51, 866–876. [Google Scholar] [CrossRef]
- Wang, J.; Hua, G.; Cai, G.; Ma, Y.; Yang, X.; Zhang, L.; Li, R.; Liu, J.; Ma, Q.; Wu, K.; et al. Genome-wide DNA methylation and transcriptome analyses reveal the key gene for wool type variation in sheep. J. Anim. Sci. Biotechnol. 2023, 14, 88. [Google Scholar] [CrossRef]
- Xu, F.; Xiao, Q.; Du, W.W.; Wang, S.; Yang, B.B. CircRNA: Functions, Applications and Prospects. Biomolecules 2025, 14, 1503. [Google Scholar] [CrossRef]
- Dong, J.; Zeng, Z.; Huang, Y.; Chen, C.; Cheng, Z.; Zhu, Q. Challenges and opportunities for circRNA identification and delivery. Crit. Rev. Biochem. Mol. Biol. 2023, 58, 19–35. [Google Scholar] [CrossRef]
- Chen, L.-L.; Kim, V.N. Small and long non-coding RNAs: Past, present, and future. Cell 2024, 187, 6451–6485. [Google Scholar] [CrossRef]
- Misir, S.; Wu, N.; Yang, B.B. Specific expression and functions of circular RNAs. Cell Death Differ. 2022, 29, 481–491. [Google Scholar] [CrossRef]
- Fan, X.; Yang, Y.; Chen, C.; Wang, Z. Pervasive translation of circular RNAs driven by short IRES-like elements. Nat. Commun. 2022, 13, 3751. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Tang, Q.; Hu, S.; Chen, Z.; Zhou, X.; Zeng, B.; Wang, Y.; He, M.; Li, Y.; Gui, L.; et al. A pig BodyMap transcriptome reveals diverse tissue physiologies and evolutionary dynamics of transcription. Nat. Commun. 2021, 12, 3715. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Chen, M.; Sooranna, S.; Shi, D.; Liu, Q.; Li, H. The emerging roles of circRNAs in traits associated with livestock breeding. Wiley Interdiscip. Rev. RNA 2023, 14, e1775. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, W.; Wang, J.; Xu, X.; Zhang, T.; Wang, L.; Feng, X. Unveiling Circular RNA-Mediated Regulatory Mechanisms in Necroptosis in Premature Ovarian Failure. Front. Biosci. (Landmark Edit.) 2023, 28, 314. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Hu, B.; Xie, Y.; Wang, D.; Zhang, J.; Chen, T.; Luo, J.; Wang, S.; Jiang, Q.; et al. Emerging Roles of Heat-Induced circRNAs Related to Lactogenesis in Lactating Sows. Front. Genet. 2019, 10, 1347. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, S.; Hu, L.; Fang, H.; Chen, G.; Ma, X.; Yu, Y.; Wang, Y.; Xu, Q. Analysis of CircRNA Expression in Peripheral Blood of Holstein Cows in Response to Heat Stress. Int. J. Mol. Sci. 2023, 24, 10150. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Z.; Zhuang, X.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Identification of circRNA-Associated-ceRNA Networks Involved in Milk Fat Metabolism under Heat Stress. Int. J. Mol. Sci. 2020, 21, 4162. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, B.; Xiong, J.; Chen, T.; Xi, Q.; Luo, J.; Jiang, Q.; Sun, J.; Zhang, Y. Genomewide analysis of circular RNA in pituitaries of normal and heat-stressed sows. BMC Genom. 2019, 20, 1013. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, L.; Wang, S.; Chen, G.; Brito, L.; Li, B.; Xu, Q.; Wang, Y. Heat Tolerance-Associated circRNA3685 Regulates Apoptosis and Autophagy in Bovine Mammary Epithelial Cells via Sponging bta-miR-138. J. Agric. Food Chem. 2025, 73, 1656–1671. [Google Scholar] [CrossRef]
- Ciliberti, M.G.; Caroprese, M.; Albenzio, M. Climate resilience in small ruminant and immune system: An old alliance in the new sustainability context. Small Rumin. Res. 2022, 210, 106662. [Google Scholar] [CrossRef]
- Laporta, J.; Khatib, H.; Zachut, M. Review: Phenotypic and molecular evidence of inter- and trans-generational effects of heat stress in livestock mammals and humans. Animal 2024, 18, 101121. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, M.; Hou, P.; Wang, W.; Shen, X.; Zhang, L.; Han, S.; Pan, C. Analysis of microbial community structure and volatile compounds in pit mud used for manufacturing Taorong-type Baijiu based on high-throughput sequencing. Sci. Rep. 2022, 12, 7347. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, P.; Du, Y.; Wang, C.; Zhang, L.; Yin, L.; Zuo, F.; Huang, W. Effect of heat stress on blood biochemistry and energy metabolite of the Dazu black goats. Front. Veter. Sci. 2024, 11, 1338643. [Google Scholar] [CrossRef]
- Fontoura, A.; Javaid, A.; de la Maza-Escolà, V.S.; Salandy, N.; Fubini, S.; Grilli, E.; McFadden, J. Heat stress develops with increased total-tract gut permeability, and dietary organic acid and pure botanical supplementation partly restores lactation performance in Holstein dairy cows. J. Dairy Sci. 2022, 105, 7842–7860. [Google Scholar] [CrossRef]
- Hu, C.; Yang, J.; Qi, Z.; Wu, H.; Wang, B.; Zou, F.; Mei, H.; Liu, J.; Wang, W.; Liu, Q. Heat shock proteins: Biological functions, pathological roles, and therapeutic opportunities. MedComm 2022, 3, e161. [Google Scholar] [CrossRef]
- Xu, W.; Meng, Z.; Deng, J.; Sun, X.; Liu, T.; Tang, Y.; Zhang, Z.; Liu, Y.; Zhu, W. Metabonomic identification of serum biomarkers related to heat stress tolerance of sheep. Anim. Sci. J. 2022, 93, e13792. [Google Scholar] [CrossRef]
- Abu Rawash, R.A.; Sharaby, M.A.; Hassan, G.E.-D.A.; Elkomy, A.E.; Hafez, E.E.; Abu Hafsa, S.H.; Salem, M.M.I. Expression profiling of HSP 70 and interleukins 2, 6 and 12 genes of Barki sheep during summer and winter seasons in two different locations. Int. J. Biometeorol. 2022, 66, 2047–2053. [Google Scholar] [CrossRef]
- Sammad, A.; Luo, H.; Hu, L.; Zhu, H.; Wang, Y. Transcriptome Reveals Granulosa Cells Coping through Redox, Inflammatory and Metabolic Mechanisms under Acute Heat Stress. Cells 2022, 11, 1443. [Google Scholar] [CrossRef]
- Li, J.; Gao, H.; Tian, Z.; Wu, Y.; Wang, Y.; Fang, Y.; Lin, L.; Han, Y.; Wu, S.; Haq, I.; et al. Effects of chronic heat stress on granulosa cell apoptosis and follicular atresia in mouse ovary. J. Anim. Sci. Biotechnol. 2016, 7, 57. [Google Scholar] [CrossRef]
- Lima, E.; Carvalho, L.; Orlandi, R.; Simões, L.; Bottino, M.; Santos, A.; de Oliveira Scarpa, F.; Sales, J. Effect of maternal heat stress at different stages of pregnancy on the reproductive performance and antral follicle count of the progeny of Holstein cows. Anim. Reprod. Sci. 2025, 272, 107665. [Google Scholar] [CrossRef]
- Vanselow, J.; Vernunft, A.; Koczan, D.; Spitschak, M.; Kuhla, B. Exposure of Lactating Dairy Cows to Acute Pre-Ovulatory Heat Stress Affects Granulosa Cell-Specific Gene Expression Profiles in Dominant Follicles. PLoS ONE 2016, 11, e0160600. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Bai, X.; Xie, X.; Chen, G.; Jia, X.; Lei, M.; Li, C.; Lai, S. Negative effects of heat stress on ovarian tissue in female rabbit. Front. Veter. Sci. 2022, 9, 1009182. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Xu, Y.; Mao, C.; Wang, Z.; Guo, S.; Jin, X.; Yan, S.; Shi, B. Effects of heat stress on antioxidant status and immune function and expression of related genes in lambs. Int. J. Biometeorol. 2020, 64, 2093–2104. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Yu, W.; Liu, J.; Tang, D.; Yang, L.; Chen, X. Oxidative cell death in cancer: Mechanisms and therapeutic opportunities. Cell Death Dis. 2024, 15, 556. [Google Scholar] [CrossRef]
- Ye, X.-Q.; Zhu, Y.-R.; Yang, Y.-Y.; Qiu, S.-J.; Liu, W.-C. Biogenic Selenium Nanoparticles Synthesized with Alginate Oligosaccharides Alleviate Heat Stress-Induced Oxidative Damage to Organs in Broilers through Activating Nrf2-Mediated Anti-Oxidation and Anti-Ferroptosis Pathways. Antioxidants 2023, 12, 1973. [Google Scholar] [CrossRef]
- Jing, J.; Zeng, H.; Shao, Q.; Tang, J.; Wang, L.; Jia, G.; Liu, G.; Chen, X.; Tian, G.; Cai, J.; et al. Selenomethionine alleviates environmental heat stress induced hepatic lipid accumulation and glycogen infiltration of broilers via maintaining mitochondrial and endoplasmic reticulum homeostasis. Redox Biol. 2023, 67, 102912. [Google Scholar] [CrossRef]
- Yang, L.; Wilusz, J.; Chen, L. Biogenesis and Regulatory Roles of Circular RNAs. Annu. Rev. Cell Dev. Biol. 2022, 38, 263–289. [Google Scholar] [CrossRef]
- Yi, Q.; Feng, J.; Lan, W.; Shi, H.; Sun, W.; Sun, W. CircRNA and lncRNA-encoded peptide in diseases, an update review. Mol. Cancer 2024, 23, 214. [Google Scholar] [CrossRef]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G. Non-coding RNAs in disease: From mechanisms to therapeutics. Nat. Rev. Genet. 2024, 25, 211–232. [Google Scholar] [CrossRef]
- Roach, C.; Bidne, K.; Romoser, M.; Ross, J.; Baumgard, L.; Keating, A. Impact of heat stress on prolactin-mediated ovarian JAK-STAT signaling in postpubertal gilts. J. Anim. Sci. 2022, 100, skac118. [Google Scholar] [CrossRef]
- Khan, M.Z.; Khan, A.; Chen, W.; Chai, W.; Wang, C. Advancements in Genetic Biomarkers and Exogenous Antioxidant Supplementation for Safeguarding Mammalian Cells against Heat-Induced Oxidative Stress and Apoptosis. Antioxidants 2024, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.-F.; Xu, J.; Wang, X.-L.; Li, S.-J.; Han, Z.-Y. Nicotinamide mononucleotide alleviates heat stress-induced oxidative stress and apoptosis in BMECs through reducing mitochondrial damage and endoplasmic reticulum stress. Ecotoxicol. Environ. Saf. 2022, 235, 113441. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, M.Z.; Umer, S.; Khan, I.M.; Xu, H.; Zhu, H.; Wang, Y. Cellular and Molecular Adaptation of Bovine Granulosa Cells and Oocytes under Heat Stress. Animals 2020, 10, 110. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Q.; Wei, C.; Sun, Y.; Li, Y.; Wei, Y.; Jiang, Q.; Huang, Y. EGCG Alleviates Skeletal Muscle Oxidative Damage in Heat-Stressed Pigs via Keap1/PGAM5 Complex-Mediated Mitophagy. J Agric. Food Chem. 2024, 73, 425–437. [Google Scholar] [CrossRef]
- Ning, Z.; Deng, X.; Li, L.; Feng, J.; Du, X.; Amevor, F.K.; Tian, Y.; Li, L.; Rao, Y.; Yi, Z.; et al. miR-128-3p regulates chicken granulosa cell function via 14-3-3β/FoxO and PPAR-γ/LPL signaling pathways. Int. J. Biol. Macromol. 2023, 241, 124654. [Google Scholar] [CrossRef]
- Liu, T.; Yuan, L.; Zou, X. Circular RNA circ-BNC2 (hsa_circ_0008732) inhibits the progression of ovarian cancer through microRNA-223-3p/FBXW7 axis. J. Ovarian Res. 2022, 15, 95. [Google Scholar] [CrossRef]
- Cesaratto, L.; Grisard, E.; Coan, M.; Zandonà, L.; De Mattia, E.; Poletto, E.; Cecchin, E.; Puglisi, F.; Canzonieri, V.; Mucignat, M.; et al. BNC2 is a putative tumor suppressor gene in high-grade serous ovarian carcinoma and impacts cell survival after oxidative stress. Cell Death Dis. 2016, 7, e2526. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, Y.; Dong, J.; Zhi, Y.; Wei, G.; Kang, X.; Liu, X. Exploring the functional variations of key candidate genes affecting egg production by hypothalamic-pituitary-ovarian axis in chickens. Poult. Sci. 2025, 104, 105027. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Content | Nutritional Level | Content |
|---|---|---|---|
| Soybean meal | 5 | Dry Matter (DM)/% | 58.52 |
| Silage corn | 48 | Digestible Energy (DE)/(MJ/Kg) | 15.18 |
| Peanut vines | 11 | Crude Protein (CP)/% | 8.88 |
| Rice bran | 8.3 | Crude Fat (EE)/% | 1.54 |
| Dried cassava distillers’ grains | 7.7 | Crude Fiber (CF)/% | 8.98 |
| Premix 1 | 20 | Crude Ash (Ash)/% | 6.43 |
| Total | 100 | Calcium (Ca)/% | 0.28 |
| Total Phosphorus (TP)/% | 0.19 |
| Items | Con Group (Mean ± SEM) | HS Group (Mean ± SEM) |
|---|---|---|
| THI | 69.36 ± 1.9 b | 91.54 ± 0.93 a |
| RR (times/min) | 29.61 ± 1.03 b | 100.17 ± 1.75 a |
| PR (beats/min) | 72.04 ± 0.69 b | 132.83 ± 1.54 a |
| RT (°C) | 39.19 ± 0.04 b | 39.90 ± 0.05 a |
| Items | Con Group (n = 3) | HS Group (n = 3) |
|---|---|---|
| Primordial follicle | 334.67 ± 40.55 a | 234.83 ± 50.65 b |
| Primary follicle | 123.33 ± 43.82 a | 68.50 ± 25.56 b |
| Secondary follicle | 47.17 ± 18.00 a | 19.17 ± 4.67 b |
| Mature follicle | 6.17 ± 1.47 a | 2.83 ± 0.75 b |
| Antral follicle | 82.50 ± 10.97 b | 101.50 ± 12.85 a |
| Ovarian weight (g) | 1.40 ± 0.16 a | 1.01 ± 0.13 b |
| Items | Total Reads | GC Content (%) | Total Mapped | Multiple Mapped | Unique Mapped |
|---|---|---|---|---|---|
| Con 1 | 77,664,144 | 52.45 | 72,839,894 (93.79%) | 16,106,111 (22.11%) | 56,733,783 (77.89%) |
| Con 2 | 71,290,888 | 50.56 | 67,100,196 (94.12%) | 11,664,988 (17.38%) | 55,435,208 (82.62%) |
| Con 3 | 65,846,436 | 50.91 | 62,029,594 (94.20%) | 10,195,125 (16.44%) | 51,834,469 (83.56%) |
| HS 1 | 66,346,110 | 50.68 | 62,322,041 (93.93%) | 11,382,140 (18.26%) | 50,939,901 (81.74%) |
| HS 2 | 65,255,110 | 48.61 | 61,110,183 (93.65%) | 11,746,421 (19.22%) | 49,363,762 (80.78%) |
| HS 3 | 75,806,612 | 47.53 | 71,265,914 (94.01%) | 12,548,269 (17.61%) | 58,717,645 (82.39%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Wei, L.; Mo, Z.; Liang, Y.; Lu, J.; Zou, J.; Wang, F.; Wu, S.; He, H.; Li, W.; et al. Impact of Heat Stress on Ovarian Function and circRNA Expression in Hu Sheep. Animals 2025, 15, 2063. https://doi.org/10.3390/ani15142063
Zou J, Wei L, Mo Z, Liang Y, Lu J, Zou J, Wang F, Wu S, He H, Li W, et al. Impact of Heat Stress on Ovarian Function and circRNA Expression in Hu Sheep. Animals. 2025; 15(14):2063. https://doi.org/10.3390/ani15142063
Chicago/Turabian StyleZou, Jianwei, Lili Wei, Zhihua Mo, Yishan Liang, Jun Lu, Juhong Zou, Fan Wang, Shaoqiang Wu, Hai’en He, Wenman Li, and et al. 2025. "Impact of Heat Stress on Ovarian Function and circRNA Expression in Hu Sheep" Animals 15, no. 14: 2063. https://doi.org/10.3390/ani15142063
APA StyleZou, J., Wei, L., Mo, Z., Liang, Y., Lu, J., Zou, J., Wang, F., Wu, S., He, H., Li, W., Huang, Y., & Jiang, Q. (2025). Impact of Heat Stress on Ovarian Function and circRNA Expression in Hu Sheep. Animals, 15(14), 2063. https://doi.org/10.3390/ani15142063