
Analysis of the Alterations in Symbiotic Microbiota and Their Correlation with Intestinal Metabolites in Rainbow Trout (Oncorhynchus mykiss) Under Heat Stress Conditions


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Facilities
2.2. Sample Collection
2.3. Microbiota Analyses
2.4. Metabolic Profile Analyses of Gut Content
2.5. Data Statistics and Analysis
3. Results
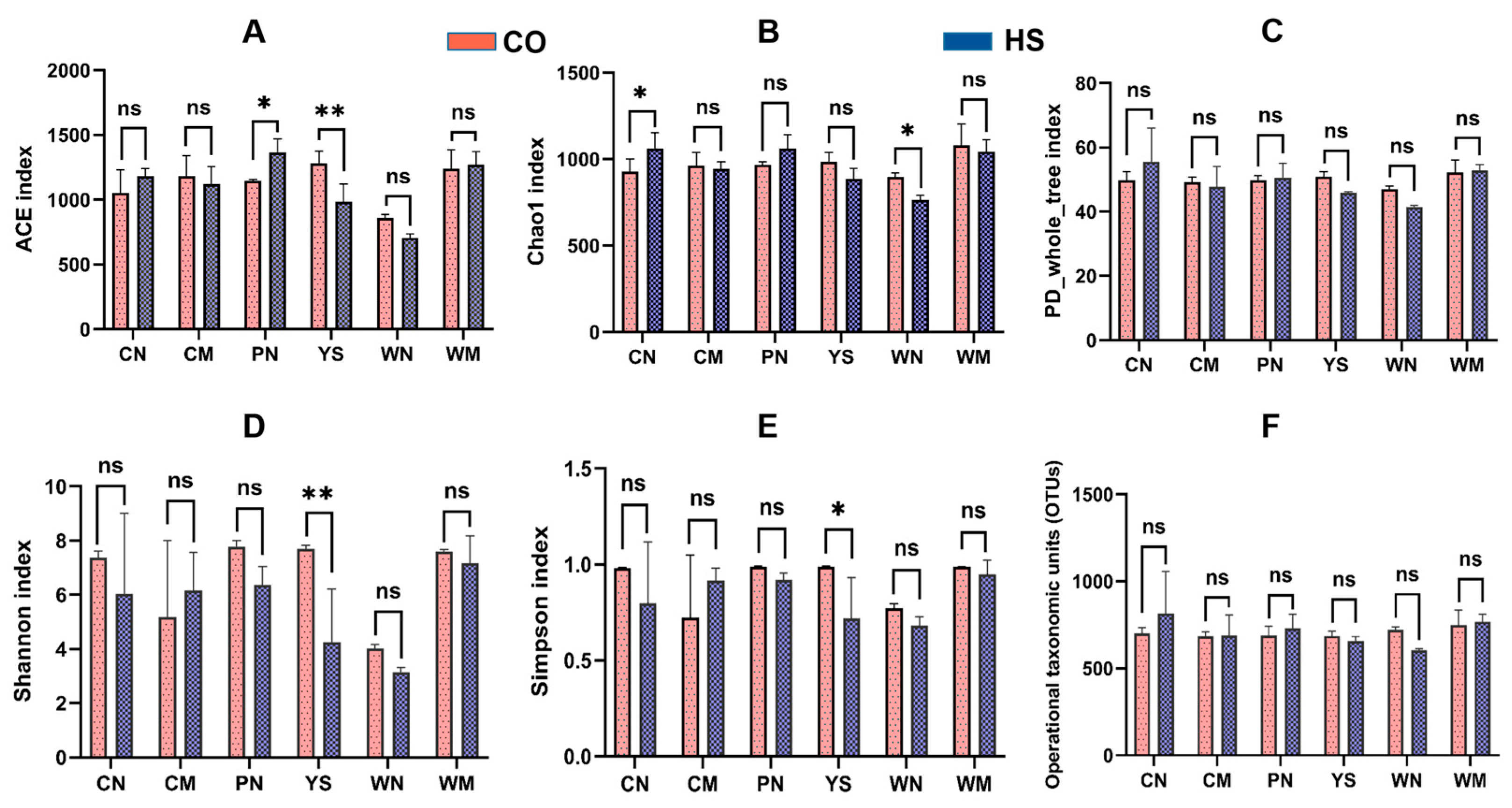
3.1. Diversity and Structure of the Bacterial Communities Under Heat Stress
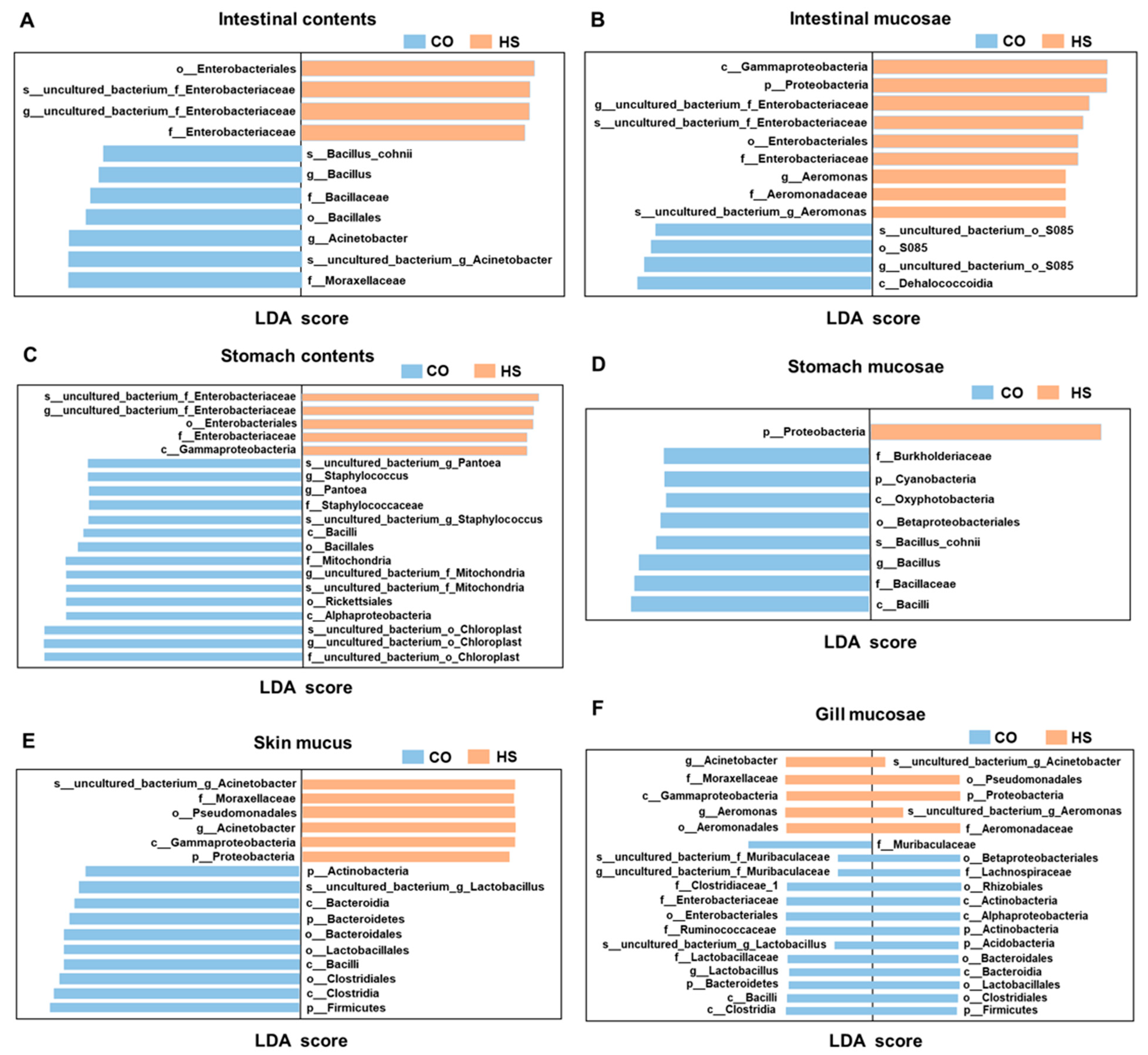
3.2. Taxonomic Shifts in Bacterial Populations Triggered by Heat Stress
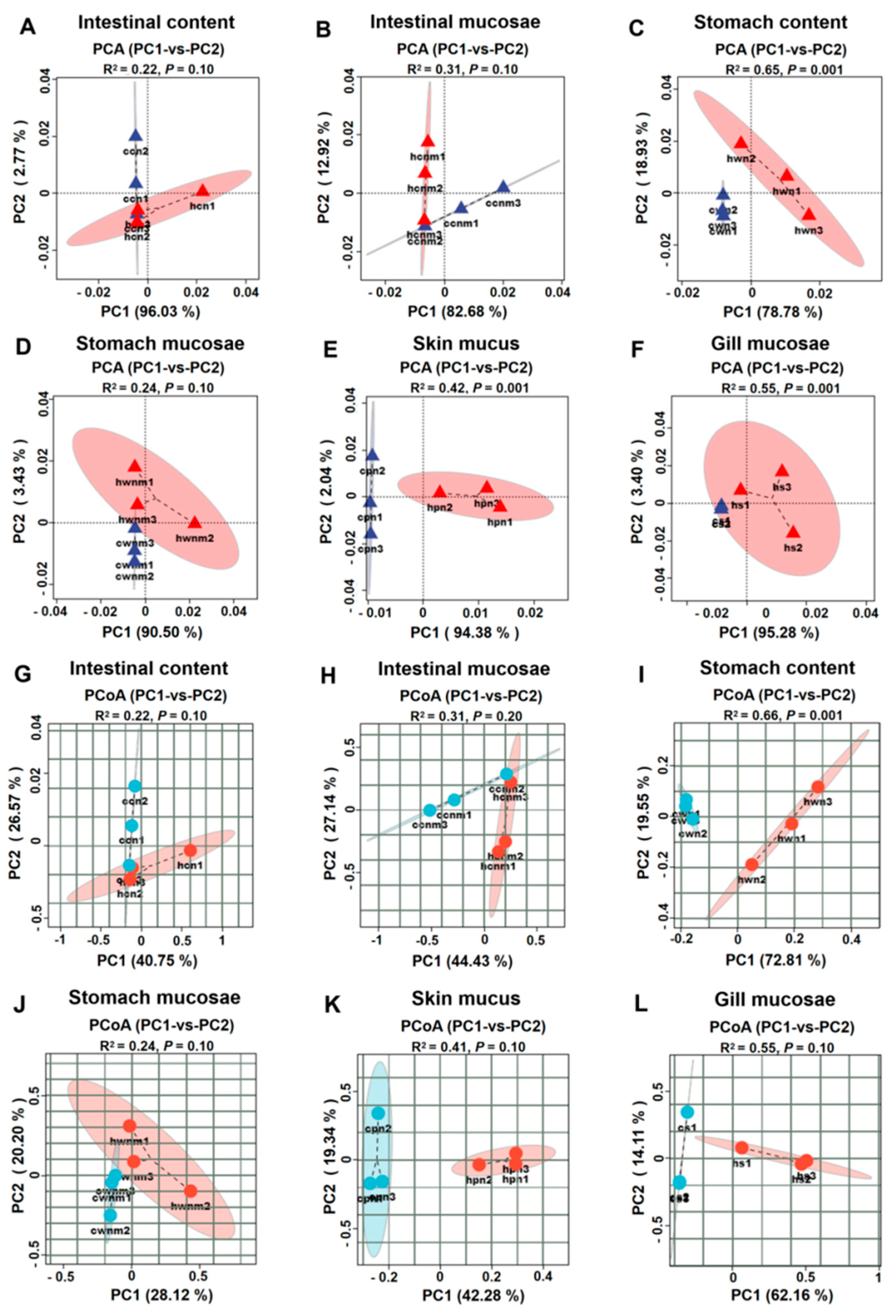
3.3. Comparative Analysis of Microbiota Across Anatomical Sites Under Heat Stress
3.4. Effects of Heat Stress on Intestinal Metabolism
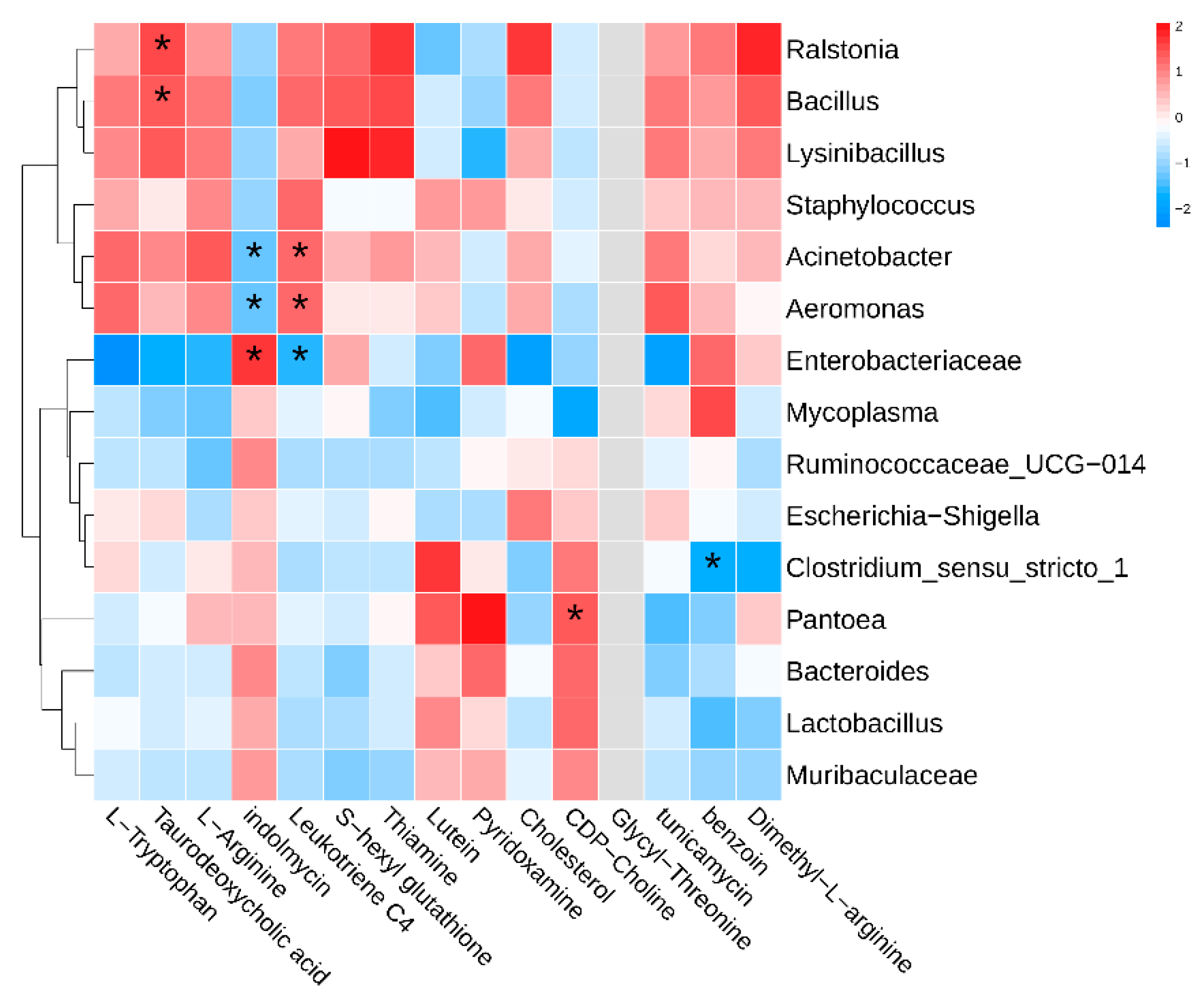
3.5. Intestinal Metabolomic Landscape and Its Association with Gut Microbiota
4. Discussion
4.1. Disruption of Commensal Microbiota Under Heat Stress Conditions
4.2. Association Analysis of Gut Microbiota and Its Metabolites Under Heat Stress Condition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huyben, D.; Vidakovic, A.; Sundh, H.; Sundell, K.; Kiessling, A.; Lundh, T. Haematological and intestinal health parameters of rainbow trout are influenced by dietary live yeast and increased water temperature. Fish Shellfish Immunol. 2019, 89, 525–536. [Google Scholar] [PubMed]
- Zhou, C.Q.; Zhou, P.; Ren, Y.L.; Cao, L.H.; Wang, J.L. Physiological response and miRNA-mRNA interaction analysis in the head kidney of rainbow trout exposed to acute heat stress. J. Therm. Biol. 2019, 83, 134–141. [Google Scholar]
- Bonga, S.E.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar]
- Zhu, L.; Zhu, W.; Zhao, T.; Chen, H.; Zhao, C.; Xu, L.; Chang, Q.; Jiang, J. Environmental Temperatures Affect the Gastrointestinal Microbes of the Chinese Giant Salamander. Front. Microbiol. 2021, 12, 543767. [Google Scholar]
- Fontaine, S.S.; Novarro, A.J.; Kohl, K.D. Environmental temperature alters the digestive performance and gut microbiota of a terrestrial amphibian. J. Exp. Biol. 2018, 221 Pt 20, jeb187559. [Google Scholar]
- Bestion, E.; Jacob, S.; Zinger, L.; Di Gesu, L.; Richard, M.; White, J.; Cote, J. Climate warming reduces gut microbiota diversity in a vertebrate ectotherm. Nat. Ecol. Evol. 2017, 1, 161. [Google Scholar]
- Zhu, L.; Liao, R.; Wu, N.; Zhu, G.; Yang, C. Heat stress mediates changes in fecal microbiome and functional pathways of laying hens. Appl. Microbiol. Biotechnol. 2019, 103, 461–472. [Google Scholar]
- Huyben, D.; Sun, L.; Moccia, R.; Kiessling, A.; Dicksved, J. Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 2018, 124, 1377–1392. [Google Scholar]
- Mongkol, P.; Bunphimpapha, P.; Rungrassamee, W.; Arayamethakorn, S.; Klinbunga, S.; Menasveta, P.; Chaiyapechara, S. Bacterial community composition and distribution in different segments of the gastrointestinal tract of wild-caught adult Penaeus monodon. Aquac. Res. 2018, 49, 378–392. [Google Scholar]
- Gajardo, K.; Rodiles, A.; Kortner, T.M.; Krogdahl, Å.; Bakke, A.M.; Merrifield, D.L.; Sørum, H. A high-resolution map of the gut microbiota in Atlantic salmon (Salmo salar): A basis for comparative gut microbial research. Sci. Rep. 2016, 6, 30893. [Google Scholar]
- Pérez, T.; Alba, C.; Aparicio, M.; de Andrés, J.; Santa Quiteria, J.A.R.; Rodríguez, J.M.; Gibello, A. Abundant bacteria in the proximal and distal intestine of healthy Siberian sturgeons (Acipenser baerii). Aquaculture 2019, 506, 325–336. [Google Scholar]
- Wan, X.-L.; McLaughlin, R.W.; Zheng, J.-S.; Hao, Y.-J.; Fan, F.; Tian, R.-M.; Wang, D. Microbial communities in different regions of the gastrointestinal tract in East Asian finless porpoises (Neophocaena asiaeorientalis sunameri). Sci. Rep. 2018, 8, 14142. [Google Scholar]
- Zhang, Z.; Li, D.; Xu, W.; Tang, R.; Li, L. Microbiome of co-cultured fish exhibits host selection and niche differentiation at the organ scale. Front. Microbiol. 2019, 10, 2576. [Google Scholar]
- Turnbaugh, P.J.; Gordon, J.I. An invitation to the marriage of metagenomics and metabolomics. Cell Metab. 2009, 134, 708–713. [Google Scholar]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.Z.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar]
- Tian, H.; Wen, Z.; Liu, Z.; Guo, Y.; Liu, G.; Sun, B. Comprehensive analysis of microbiome, metabolome and transcriptome revealed the mechanisms of Moringa oleifera polysaccharide on preventing ulcerative colitis. Int. J. Biol. Macromol. 2022, 222 Pt A, 573–586. [Google Scholar]
- Merrifield, D.L.; Rodiles, A. The fish microbiome and its interactions with mucosal tissues. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 273–295. [Google Scholar]
- Yu, Y.Y.; Ding, L.G.; Huang, Z.Y.; Xu, H.Y.; Xu, Z. Commensal bacteria-immunity crosstalk shapes mucosal homeostasis in teleost fish. Rev. Aquac. 2021, 13, 2322–2343. [Google Scholar]
- Meng, K.F.; Ding, L.G.; Wu, S.; Wu, Z.B.; Cheng, G.F.; Zhai, X.; Sun, R.H.; Xu, Z. Interactions Between Commensal Microbiota and Mucosal Immunity in Teleost Fish During Viral Infection with SVCV. Front. Immunol. 2021, 12, 654758. [Google Scholar]
- Sylvain, F.; Holland, A.; Bouslama, S.; Audet-Gilbert, É.; Lavoie, C.; Val, A.L.; Derome, N. Fish Skin and Gut Microbiomes Show Contrasting Signatures of Host Species and Habitat. Appl. Environ. Microbiol. 2020, 86, e00789-20. [Google Scholar]
- Legrand, T.; Catalano, S.R.; Wos-Oxley, M.L.; Stephens, F.; Landos, M.; Bansemer, M.S.; Stone, D.A.J.; Qin, J.G.; Oxley, A.P.A. The Inner Workings of the Outer Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail Kingfish. Front. Microbiol. 2017, 8, 2664. [Google Scholar]
- Llewellyn, M.S.; McGinnity, P.; Dionne, M.; Letourneau, J.; Thonier, F.; Carvalho, G.R.; Creer, S.; Derome, N. The biogeography of the Atlantic salmon (Salmo salar) gut microbiome. ISME J. 2016, 10, 1280–1284. [Google Scholar] [PubMed]
- Sylvain, F.; Holland, A.; Audet-Gilbert, É.; Luis Val, A.; Derome, N. Amazon fish bacterial communities show structural convergence along widespread hydrochemical gradients. Mol. Ecol. 2019, 28, 3612–3626. [Google Scholar] [PubMed]
- Mohammed, H.H.; Arias, C.R. Potassium permanganate elicits a shift of the external fish microbiome and increases host susceptibility to columnaris disease. Vet. Res. 2015, 46, 82. [Google Scholar] [PubMed]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar]
- Zhang, X.; Ding, L.; Yu, Y.; Kong, W.; Yin, Y.; Huang, Z.; Zhang, X.; Xu, Z. The Change of Teleost Skin Commensal Microbiota Is Associated with Skin Mucosal Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front. Immunol. 2018, 9, 2972. [Google Scholar]
- de Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 2018, 94, fix161. [Google Scholar]
- Lowrey, L.; Woodhams, D.C.; Tacchi, L.; Salinas, I. Topographical Mapping of the Rainbow Trout (Oncorhynchus mykiss) Microbiome Reveals a Diverse Bacterial Community with Antifungal Properties in the Skin. Appl. Environ. Microbiol. 2015, 81, 6915–6925. [Google Scholar]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar]
- Hao, J.; Cai, W.; Li, K.; Guan, H.; Yang, C.; Gao, Q.; Liu, D.; Nie, M.; Qi, D.; Zhang, C. High-throughput sequencing was used to analyze the microbial diversity of skin and gut in Microcephalus. Acta Microbiol. Sin. China 2023, 63, 233–250. [Google Scholar]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar]
- Schmidt, V.T.; Smith, K.F.; Melvin, D.W.; Amaral-Zettler, L.A. Community assembly of a euryhaline fish microbiome during salinity acclimation. Mol. Ecol. 2015, 24, 2537–2550. [Google Scholar]
- Lokesh, J.; Kiron, V. Transition from freshwater to seawater reshapes the skin-associated microbiota of Atlantic salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar]
- Geng, C.; Sun, Y.; Li, B.; Ding, L.; Liu, Y.; Wei, X.; Liu, W.; Han, L.; Yuan, F.; Wang, P.; et al. Physiological response to heat stress in the gill of rainbow trout (Oncorhynchus mykiss) revealed based on UPLC-Q-TOF/MS metabolomics. J. Hydrobiol. 2023, 47, 1–11. [Google Scholar]
- Hess, S.; Wenger, A.S.; Ainsworth, T.D.; Rummer, J.L. Exposure of clownfish larvae to suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and microbiome. Sci. Rep. 2015, 5, 10561. [Google Scholar]
- Rudi, K.; Angell, I.L.; Pope, P.B.; Vik, J.O.; Sandve, S.R.; Snipen, L.-G. Stable core gut microbiota across the freshwater-to-saltwater transition for farmed Atlantic salmon. Appl. Environ. Microbiol. 2018, 84, e01974-17. [Google Scholar]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. BioMed Res. Int. 2017, 2017, 9351507. [Google Scholar]
- Zhou, C.; Yang, S.; Ka, W.; Gao, P.; Li, Y.; Long, R.; Wang, J. Association of gut microbiota with metabolism in rainbow trout under acute heat stress. Front. Microbiol. 2022, 13, 846336. [Google Scholar]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte metabolism shapes the gut microbiota. Science 2018, 362, 6418. [Google Scholar]
- Jia, Z.; Wu, A.; He, M.; Zhang, L.; Wang, C.; Chen, A. Metabolites of stable fly reduce diarrhea in mice by modulating the immune system, antioxidants, and composition of gut microbiota. Microb. Pathog. 2019, 134, 103557. [Google Scholar]
- He, K.; Yan, W.; Sun, C.; Liu, J.; Bai, R.; Wang, T.; Qian, W. Alterations in the diversity and composition of gut microbiota in weaned piglets infected with Balantioides coli. Vet. Parasitol. 2020, 288, 109298. [Google Scholar] [PubMed]
- Cardman, Z.; Arnosti, C.; Durbin, A.; Ziervogel, K.; Cox, C.; Steen, A.; Teske, A. Verrucomicrobia are candidates for polysaccharide-degrading bacterioplankton in an arctic fjord of Svalbard. Appl. Environ. Microbiol. 2014, 80, 3749–3756. [Google Scholar]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Hermie, J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar]
- Hao, Y.T.; Wu, S.G.; Jakovlić, I.; Zou, H.; Li, W.X.; Wang, G.T. Impacts of diet on hindgut microbiota and short-chain fatty acids in grass carp (Ctenopharyngodon idellus). Aquac. Res. 2017, 48, 5595–5605. [Google Scholar]
- Cao, S.; Geng, Y.; Yu, Z.; Deng, L.; Gan, W.; Wang, K.; Ou, Y.; Chen, D.; Huang, X.; Zuo, Z.; et al. Acinetobacter lwoffii, an emerging pathogen for fish in Schizothorax genus in China. Transbound. Emerg. Dis. 2018, 65, 1816–1822. [Google Scholar]
- Reid, K.M.; Patel, S.; Robinson, A.J.; Bu, L.; Jarungsriapisit, J.; Moore, L.J.; Salinas, I. Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.) post-smolts. PLoS ONE 2017, 12, e0172856. [Google Scholar]
- Jin, W.; Han, K.; Dong, S.; Yang, Y.; Mao, Z.; Su, M.; Zeng, M. Modifications in gut microbiota and fermentation metabolites in the hindgut of rats after the consumption of galactooligosaccharide glycated with a fish peptide. Food Funct. 2018, 9, 2853–2864. [Google Scholar]
- Austin, B.; Austin, D.A.; Munn, C. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Yu, Y.Y.; Kong, W.G.; Xu, H.Y.; Huang, Z.Y.; Zhang, X.T.; Ding, L.G.; Dong, S.; Yin, G.M.; Dong, F.; Cao, J.F.; et al. Convergent Evolution of Mucosal Immune Responses at the Buccal Cavity of Teleost Fish. iScience 2019, 19, 821–835. [Google Scholar]
- Salosensaari, A.; Laitinen, V.; Havulinna, A.S.; Meric, G.; Cheng, S.; Perola, M.; Valsta, L.; Alfthan, G.; Inouye, M.; Watrous, J.D.; et al. Taxonomic signatures of cause-specific mortality risk in human gut microbiome. Nat. Commun. 2021, 12, 2671. [Google Scholar]
- Bozzi, D.; Rasmussen, J.A.; Carøe, C.; Sveier, H.; Nordøy, K.; Gilbert, M.T.P.; Limborg, M.T. Salmon gut microbiota correlates with disease infection status: Potential for monitoring health in farmed animals. Anim. Microbiome 2021, 3, 30. [Google Scholar]
- Ofek, T.; Lalzar, M.; Izhaki, I.; Halpern, M. Intestine and spleen microbiota composition in healthy and diseased tilapia. Anim. Microbiome 2022, 4, 50. [Google Scholar]
- Lyons, P.; Turnbull, J.; Dawson, K.A.; Crumlish, M. Phylogenetic and functional characterization of the distal intestinal microbiome of rainbow trout Oncorhynchus mykiss from both farm and aquarium settings. J. Appl. Microbiol. 2017, 122, 347–363. [Google Scholar]
- Zarkasi, K.Z.; Abell, G.C.; Taylor, R.S.; Neuman, C.; Hatje, E.; Tamplin, M.L.; Katouli, M.; Bowman, J.P. Pyrosequencing-based characterization of gastrointestinal bacteria of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl. Microbiol. 2014, 117, 18–27. [Google Scholar]
- Huyben, D.; Grobler, T.; Matthew, C.; Bou, M.; Ruyter, B.; Glencross, B. Requirement for omega-3 long-chain polyunsaturated fatty acids by Atlantic salmon is relative to the dietary lipid level. Aquaculture 2021, 531, 735805. [Google Scholar]
- Burns, A.R.; Watral, V.; Sichel, S.; Spagnoli, S.; Banse, A.V.; Mittge, E.; Sharpton, T.J.; Guillemin, K.; Kent, M.L. Transmission of a common intestinal neoplasm in zebrafish by cohabitation. J. Fish Dis. 2018, 41, 569–579. [Google Scholar]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar]
- VanInsberghe, D.; Maas, K.R.; Cardenas, E.; Strachan, C.R.; Hallam, S.J.; Mohn, W.W. Non-symbiotic Bradyrhizobium ecotypes dominate North American forest soils. ISME J. 2015, 9, 2435–2441. [Google Scholar] [PubMed]
- Wang, D.; Ma, X.; Guo, S.; Wang, Y.; Li, T.; Zou, D.; Song, H.; Yang, W.; Ge, Y. Effect of Huangqin Tang on Urine Metabolic Profile in Rats with Ulcerative Colitis Based on UPLC-Q-Exactive Orbitrap MS. Evid.-Based Complement Altern. Med. eCAM 2020, 2020, 1874065. [Google Scholar]
- Hong, Z.C.; Cai, Q.; Wu, H.Z.; Yang, Y.F.; Fan, H.; Duan, X.Y. Compound Sophorae Decoction: Treating ulcerative colitis by affecting multiple metabolic pathways. Chin. J. Nat. Med. 2021, 19, 267–283. [Google Scholar]
- Filimoniuk, A.; Daniluk, U.; Samczuk, P.; Wasilewska, N.; Jakimiec, P.; Kucharska, M.; Lebensztejn, D.M.; Ciborowski, M. Metabolomic profiling in children with inflammatory bowel disease. Adv. Med. Sci. 2020, 65, 65–70. [Google Scholar]
- Zou, Y.; Yang, Y.; Fu, X.; He, X.; Liu, M.; Zong, T.; Li, X.; Htet Aung, L.; Wang, Z.; Yu, T. The regulatory roles of aminoacyl-tRNA synthetase in cardiovascular disease. Mol. Ther. Nucleic Acids 2021, 25, 372–387. [Google Scholar] [PubMed]
- Lin, R.; Liu, W.T.; Piao, M.Y.; Zhu, H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 2017, 49, 2083–2090. [Google Scholar] [PubMed]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar]
- Taleb, S. Tryptophan Dietary Impacts Gut Barrier and Metabolic Diseases. Front. Immunol. 2019, 10, 2113. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Community Structure | |
|---|---|---|
| R2 | p | |
| Whole comparison | 0.663 | 0.001 |
| ccn vs. hcn | 0.202 | 0.301 |
| cpn vs. hpn | 0.218 | 0.200 |
| cs vs. hs | 0.242 | 0.001 |
| cwn vs. hwn | 0.264 | 0.100 |
| cwm vs. hwm | 0.207 | 0.500 |
| ccm vs. hcm | 0.204 | 0.301 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Ding, F. Analysis of the Alterations in Symbiotic Microbiota and Their Correlation with Intestinal Metabolites in Rainbow Trout (Oncorhynchus mykiss) Under Heat Stress Conditions. Animals 2025, 15, 2017. https://doi.org/10.3390/ani15142017
Zhou C, Ding F. Analysis of the Alterations in Symbiotic Microbiota and Their Correlation with Intestinal Metabolites in Rainbow Trout (Oncorhynchus mykiss) Under Heat Stress Conditions. Animals. 2025; 15(14):2017. https://doi.org/10.3390/ani15142017
Chicago/Turabian StyleZhou, Changqing, and Fengyuan Ding. 2025. "Analysis of the Alterations in Symbiotic Microbiota and Their Correlation with Intestinal Metabolites in Rainbow Trout (Oncorhynchus mykiss) Under Heat Stress Conditions" Animals 15, no. 14: 2017. https://doi.org/10.3390/ani15142017
APA StyleZhou, C., & Ding, F. (2025). Analysis of the Alterations in Symbiotic Microbiota and Their Correlation with Intestinal Metabolites in Rainbow Trout (Oncorhynchus mykiss) Under Heat Stress Conditions. Animals, 15(14), 2017. https://doi.org/10.3390/ani15142017