
Highly Pathogenic Avian Influenza (H5N1) Clade 2.3.4.4b in Cattle: A Rising One Health Concern



Simple Summary
Abstract
1. Introduction
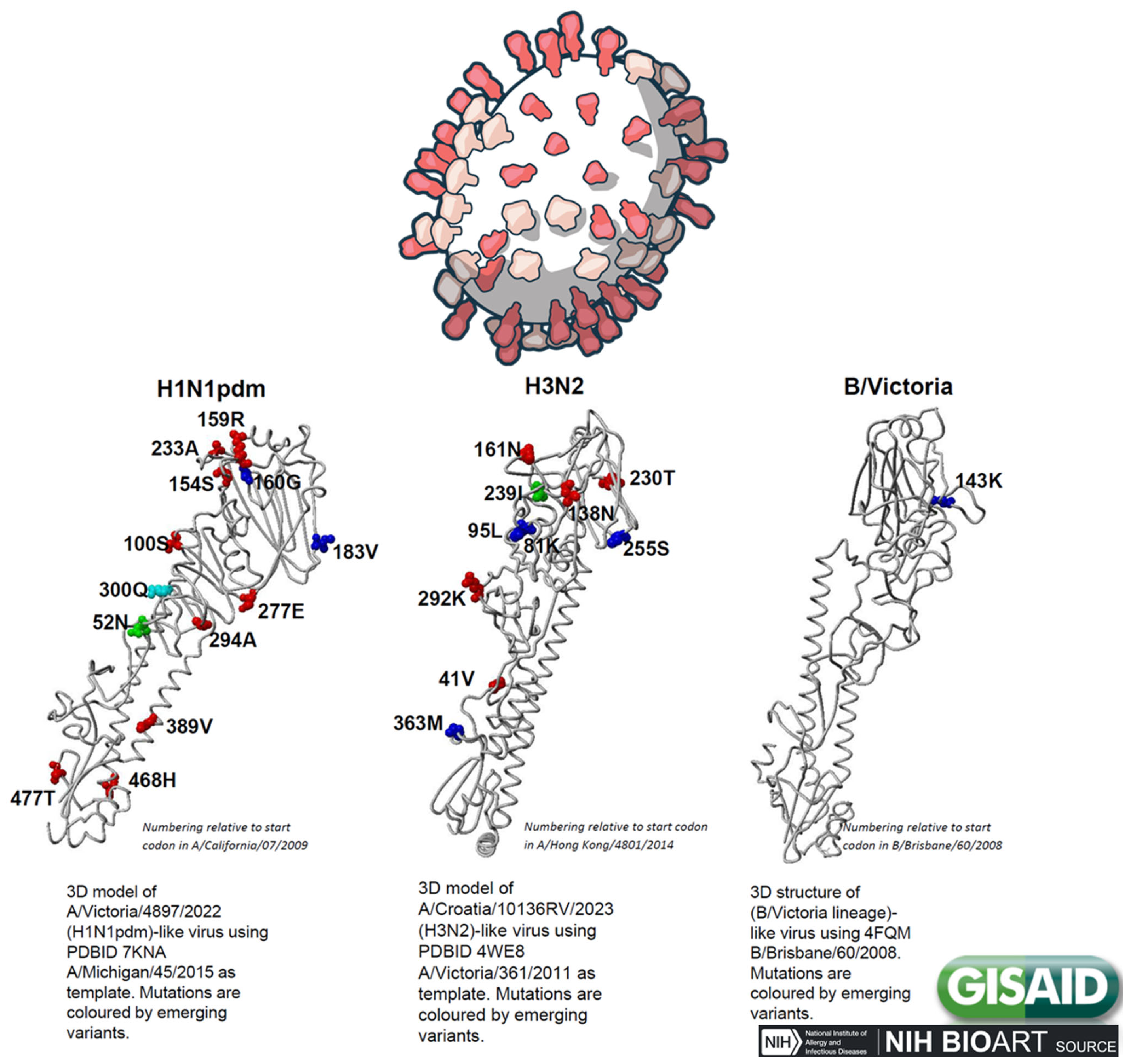
2. Nature and Classification of Influenzavirus
3. Worldwide Distribution of H5N1
4. History of Influenza Virus Infection in Livestock
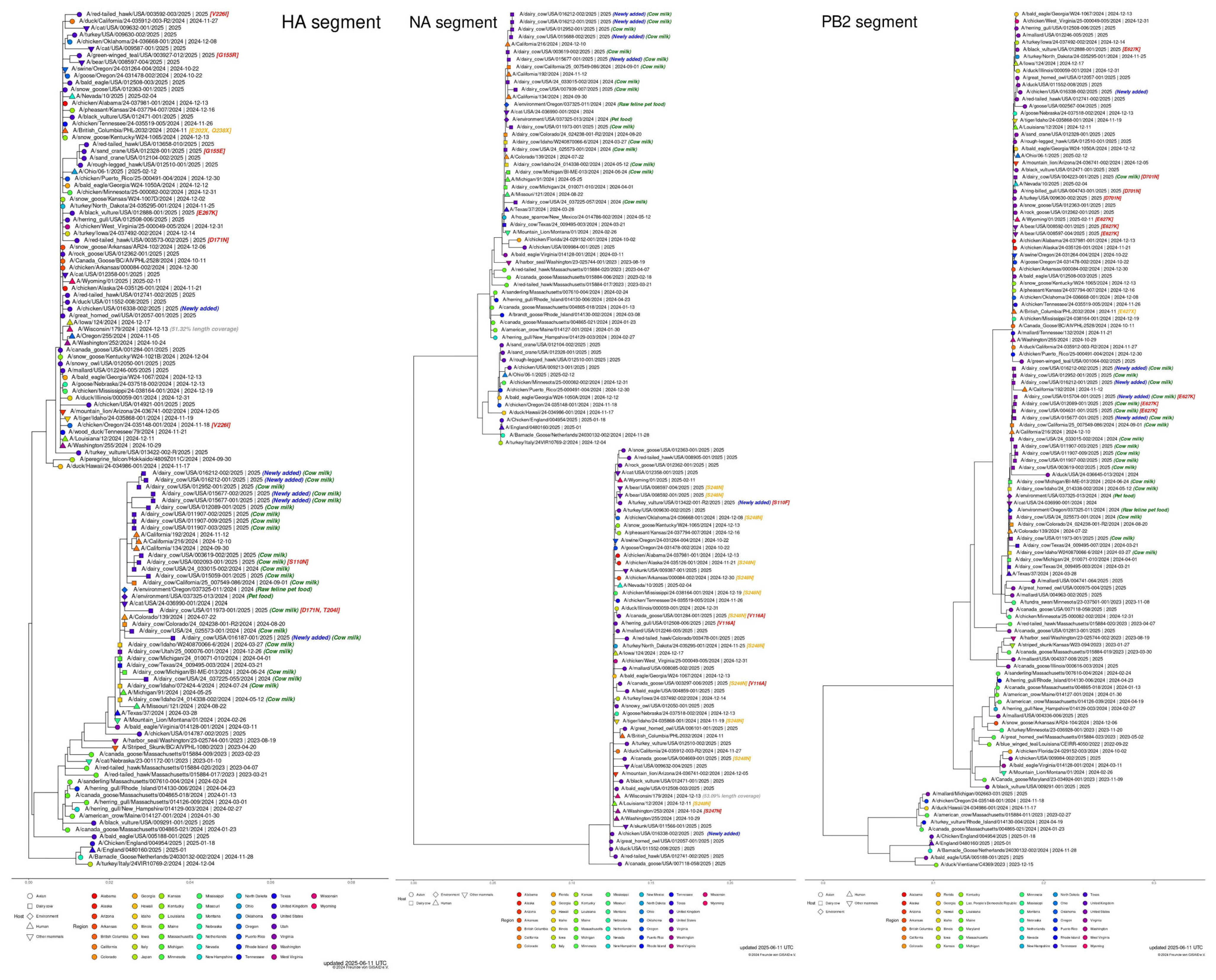
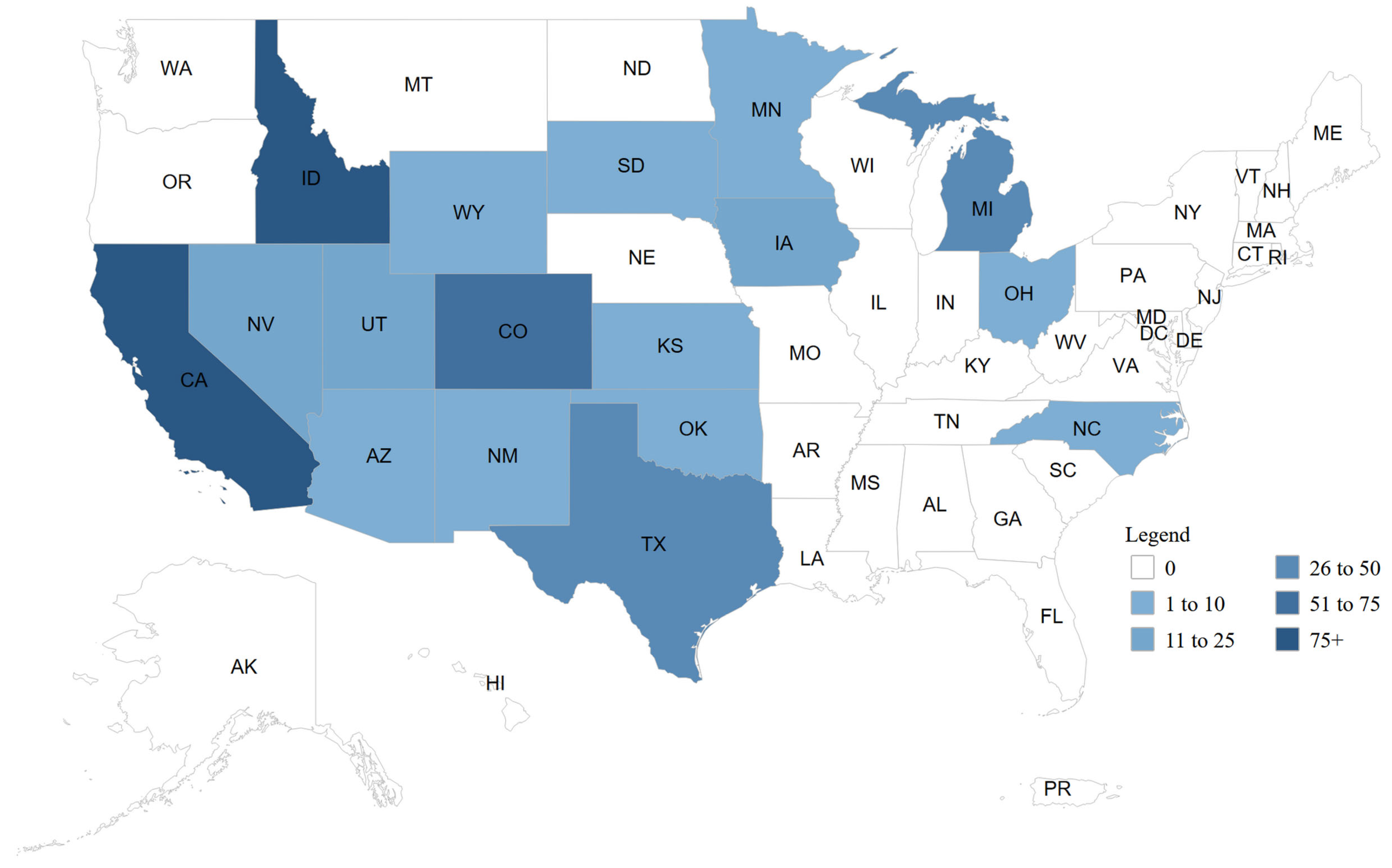
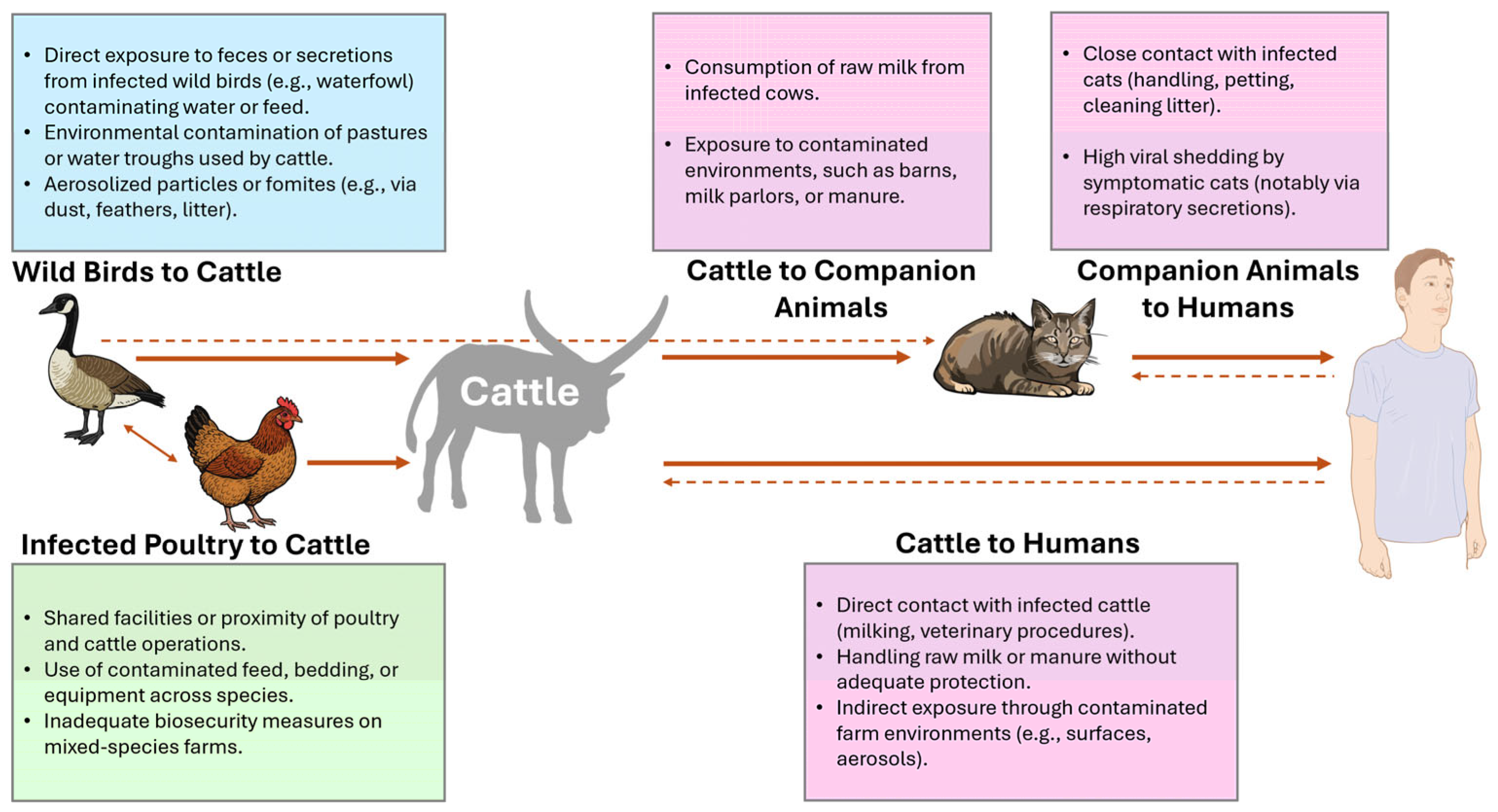
5. H5N1 in Cattle
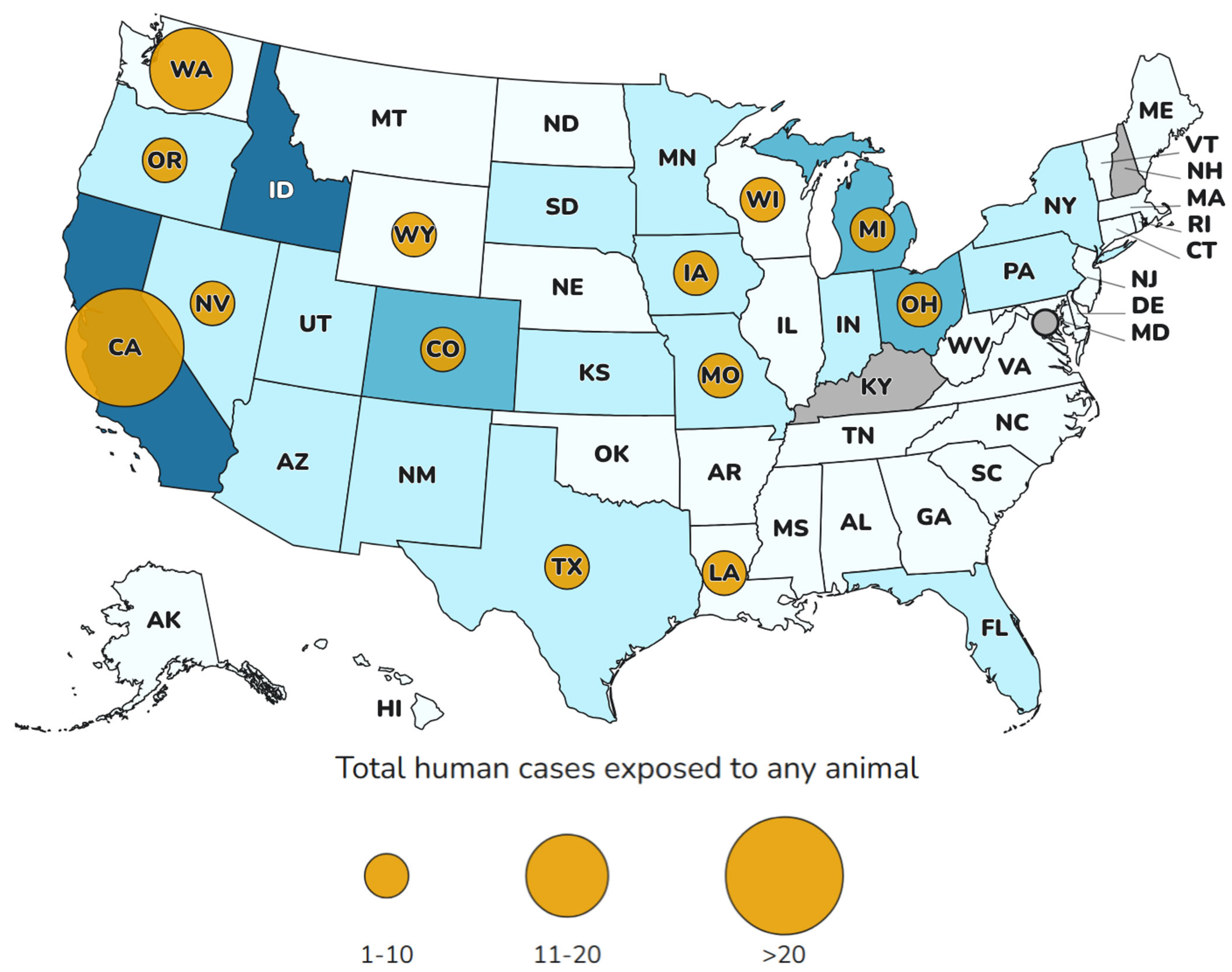
6. H5N1 in Humans
7. Limitations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hutchinson, E.C. Influenza Virus. Trends Microbiol. 2018, 26, 809–810. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Jayaswal, N.; Kumar, S.; Rao, G.; Budha, R.R.; Mohanty, A.; Mehta, R.; Apostolopoulos, V.; Sah, S.; Bonilla-Aldana, D.K.; et al. Targeting H3N2 influenza: Advancements in treatment and vaccine strategies. Expert. Rev. Anti Infect. Ther. 2025, 23, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Ryu, Y. Transmissibility and severity of influenza virus by subtype. Infect. Genet. Evol. 2018, 65, 288–292. [Google Scholar] [CrossRef]
- Rodriguez-Morales, A.J.; Hui, D.S.; Ippolito, G.; Traore, T.; Satta, G.; Everett, D.B.; Zumla, A. Avian Influenza—The next travel-associated pandemic? Proactive One Health surveillance is required to reduce the risk of the spread. Travel Med. Infect. Dis. 2025, 65, 102829. [Google Scholar] [CrossRef]
- Chaudhary, R.K.; L, A.; Patil, P.; Mateti, U.V.; Sah, S.; Mohanty, A.; Rath, R.S.; Padhi, B.K.; Malik, S.; Jassim, K.H.; et al. System Biology Approach to Identify the Hub Genes and Pathways Associated with Human H5N1 Infection. Vaccines 2023, 11, 1269. [Google Scholar] [CrossRef]
- Priyanka; Khandia, R.; Chopra, H.; Choudhary, O.P.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. The re-emergence of H3N2 influenza: An update on the risk and containment. New Microbes New Infect. 2023, 53, 101147. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Moncla, L.; Dudas, G.; VanInsberghe, D.; Sukhova, K.; Lloyd-Smith, J.O.; Worobey, M.; Lowen, A.C.; Nelson, M.I. The global H5N1 influenza panzootic in mammals. Nature 2025, 637, 304–313. [Google Scholar] [CrossRef]
- Kilbourne, E.D. Influenza pandemics of the 20th century. Emerg. Infect. Dis. 2006, 12, 9–14. [Google Scholar] [CrossRef]
- Ankomah, A.A.; Moa, A.; Chughtai, A.A. The long road of pandemic vaccine development to rollout: A systematic review on the lessons learnt from the 2009 H1N1 influenza pandemic. Am. J. Infect. Control 2022, 50, 735–742. [Google Scholar] [CrossRef]
- Charostad, J.; Rezaei Zadeh Rukerd, M.; Mahmoudvand, S.; Bashash, D.; Hashemi, S.M.A.; Nakhaie, M.; Zandi, K. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: An imminent threat at doorstep. Travel Med. Infect. Dis. 2023, 55, 102638. [Google Scholar] [CrossRef]
- Guan, Y.; Smith, G.J. The emergence and diversification of panzootic H5N1 influenza viruses. Virus Res. 2013, 178, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Chavda, V.P.; Mehta, R.; Rodriguez-Morales, A.J.; Henao-MartÍnez, A.F.; Sah, R. Alert and surveillance on H5N1 influenza virus: Risks to agriculture and public health. Ther. Adv. Infect. Dis. 2024, 11, 20499361241266521. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morales, A.J.; Bonilla-Aldana, D.K.; Paniz-Mondolfi, A.E. Concerns about influenza H5N8 outbreaks in humans and birds: Facing the next airborne pandemic? Travel Med. Infect. Dis. 2021, 41, 102054. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Aguirre-Florez, M.; Villamizar-Peña, R.; Gutiérrez-Ocampo, E.; Henao-Martínez, J.F.; Cvetkovic-Vega, A.; Dhama, K.; Rabaan, A.; Sah, R.; Rodriguez-Morales, A.J.; et al. After SARS-CoV-2, will H5N6 and other influenza viruses follow the pandemic path? Infez. Med. 2020, 28, 475–485. [Google Scholar]
- Cruz, C.D.; Icochea, M.E.; Espejo, V.; Troncos, G.; Castro-Sanguinetti, G.R.; Schilling, M.A.; Tinoco, Y. Highly Pathogenic Avian Influenza A(H5N1) from Wild Birds, Poultry, and Mammals, Peru. Emerg. Infect. Dis. 2023, 29, 2572–2576. [Google Scholar] [CrossRef]
- Neumann, G.; Kawaoka, Y. Highly pathogenic H5N1 avian influenza virus outbreak in cattle: The knowns and unknowns. Nat. Rev. Microbiol. 2024, 22, 525–526. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Saxena, A.; Magstadt, D.R.; Gauger, P.C.; Burrough, E.R.; Zhang, J.; Siepker, C.; Mainenti, M.; Gorden, P.J.; Plummer, P.J.; et al. Genomic characterization of highly pathogenic avian influenza A H5N1 virus newly emerged in dairy cattle. Emerg. Microbes Infect. 2024, 13, 2380421. [Google Scholar] [CrossRef]
- Cui, P.; Shi, J.; Wang, C.; Zhang, Y.; Xing, X.; Kong, H.; Yan, C.; Zeng, X.; Liu, L.; Tian, G.; et al. Global dissemination of H5N1 influenza viruses bearing the clade 2.3.4.4b HA gene and biologic analysis of the ones detected in China. Emerg. Microbes Infect. 2022, 11, 1693–1704. [Google Scholar] [CrossRef]
- Sreenivasan, C.C.; Li, F.; Wang, D. Emerging Threats of Highly Pathogenic Avian Influenza A (H5N1) in US Dairy Cattle: Understanding Cross-Species Transmission Dynamics in Mammalian Hosts. Viruses 2024, 16, 1703. [Google Scholar] [CrossRef]
- Ruiz-Saenz, J.; Martinez-Gutierrez, M.; Pujol, F.H. Multiple introductions of highly pathogenic avian influenza H5N1 clade 2.3.4.4b into South America. Travel Med. Infect. Dis. 2023, 53, 102591. [Google Scholar] [CrossRef]
- Rimondi, A.; Vanstreels, R.E.T.; Olivera, V.; Donini, A.; Lauriente, M.M.; Uhart, M.M. Highly Pathogenic Avian Influenza A(H5N1) Viruses from Multispecies Outbreak, Argentina, August 2023. Emerg. Infect. Dis. 2024, 30, 812–814. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, A.C.; Silva, L.M.N.; Cho, A.Y.; Repenning, M.; Amgarten, D.; de Moraes, A.P.; Malta, F.; Miller, M.; Dorlass, E.G.; Palameta, S.; et al. Incursion of Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus, Brazil, 2023. Emerg. Infect. Dis. 2024, 30, 619–621. [Google Scholar] [CrossRef]
- de Carvalho Araujo, A.; Cho, A.Y.; Silva, L.M.N.; Corrêa, T.C.; de Souza, G.C.; Albuquerque, A.S.; Macagnan, E.; Kolesnikvoas, C.K.M.; Meurer, R.; Vieira, J.V.; et al. Mortality in sea lions is associated with the introduction of the H5N1 clade 2.3.4.4b virus in Brazil October 2023: Whole genome sequencing and phylogenetic analysis. BMC Vet. Res. 2024, 20, 285. [Google Scholar] [CrossRef]
- Ahmad, F.; Haque, S.; Tawil, S.; Husni, R.; Bonilla-Aldana, D.K.; Montenegro-Idrogo, J.J.; Rodriguez-Morales, A.J. Avian influenza spillover to humans: Are we prepared to deal with another potential pandemic? Travel Med. Infect. Dis. 2023, 55, 102634. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yeom, M.; Vu, T.T.H.; Do, H.Q.; Na, W.; Lee, M.; Jeong, D.G.; Cheon, D.S.; Song, D. Characterization of highly pathogenic avian influenza A (H5N1) viruses isolated from cats in South Korea, 2023. Emerg. Microbes Infect. 2024, 13, 2290835. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, A.; Naguib, M.M.; Nogales, A.; Barre, R.S.; Stewart, J.P.; García-Sastre, A.; Martinez-Sobrido, L. Avian influenza A (H5N1) virus in dairy cattle: Origin, evolution, and cross-species transmission. mBio 2024, 15, e0254224. [Google Scholar] [CrossRef]
- Caserta, L.C.; Frye, E.A.; Butt, S.L.; Laverack, M.; Nooruzzaman, M.; Covaleda, L.M.; Thompson, A.C.; Koscielny, M.P.; Cronk, B.; Johnson, A.; et al. Spillover of highly pathogenic avian influenza H5N1 virus to dairy cattle. Nature 2024, 634, 669–676. [Google Scholar] [CrossRef]
- Burrough, E.R.; Magstadt, D.R.; Petersen, B.; Timmermans, S.J.; Gauger, P.C.; Zhang, J.; Siepker, C.; Mainenti, M.; Li, G.; Thompson, A.C.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerg. Infect. Dis. 2024, 30, 1335–1343. [Google Scholar] [CrossRef]
- Nelli, R.K.; Harm, T.A.; Siepker, C.; Groeltz-Thrush, J.M.; Jones, B.; Twu, N.C.; Nenninger, A.S.; Magstadt, D.R.; Burrough, E.R.; Piñeyro, P.E.; et al. Sialic Acid Receptor Specificity in Mammary Gland of Dairy Cattle Infected with Highly Pathogenic Avian Influenza A(H5N1) Virus. Emerg. Infect. Dis. 2024, 30, 1361–1373. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Bonilla-Aldana, J.L.; Acosta-España, J.D.; Rodriguez-Morales, A.J. Highly Pathogenic Avian Influenza H5N1 in Cats (Felis catus): A Systematic Review and Meta-Analysis. Animals 2025, 15, 1441. [Google Scholar] [CrossRef]
- Eisfeld, A.J.; Biswas, A.; Guan, L.; Gu, C.; Maemura, T.; Trifkovic, S.; Wang, T.; Babujee, L.; Dahn, R.; Halfmann, P.J.; et al. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature 2024, 633, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Honein, M.A.; Olsen, S.J.; Jernigan, D.B.; Daskalakis, D.C. Challenges and Opportunities for Wastewater Monitoring of Influenza Viruses During the Multistate Outbreak of Highly Pathogenic Avian Influenza A(H5N1) Virus in Dairy Cattle and Poultry. Am. J. Public Health 2024, 114, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; King, L.R.; Manischewitz, J.; Posadas, O.; Mishra, A.K.; Liu, D.; Beigel, J.H.; Rappuoli, R.; Tsang, J.S.; Golding, H. Licensed H5N1 vaccines generate cross-neutralizing antibodies against highly pathogenic H5N1 clade 2.3.4.4b influenza virus. Nat. Med. 2024, 30, 2771–2776. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Reed, C.; Davis, C.T.; Uyeki, T.M.; Behravesh, C.B.; Kniss, K.; Budd, A.; Biggerstaff, M.; Adjemian, J.; Barnes, J.R.; et al. Outbreak of Highly Pathogenic Avian Influenza A(H5N1) Viruses in U.S. Dairy Cattle and Detection of Two Human Cases—United States, 2024. MMWR Morb. Mortal. Wkly. Rep. 2024, 73, 501–505. [Google Scholar] [CrossRef]
- Sah, R.; Srivastava, S.; Kumar, S.; Mehta, R.; Donovan, S.; Sierra-Carrero, L.; Luna, C.; Woc-Colburn, L.; Cardona-Ospina, J.A.; Hinestroza-Jordan, M.; et al. Concerns on H5N1 avian influenza given the outbreak in U.S. dairy cattle. Lancet Reg. Health Am. 2024, 35, 100785. [Google Scholar] [CrossRef]
- Lowen, A.C.; Baker, A.L.; Bowman, A.S.; García-Sastre, A.; Hensley, S.E.; Lakdawala, S.S.; Moncla, L.H.; Nelson, M.I.; Pekosz, A.; Poulson, R.L.; et al. Pandemic risk stemming from the bovine H5N1 outbreak: An account of the knowns and unknowns. J. Virol. 2025, 99, e0005225. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M. Evolution and mutational landscape of highly pathogenic avian influenza strain A(H5N1) in the current outbreak in the USA and global landscape. Virology 2024, 600, 110246. [Google Scholar] [CrossRef]
- Kristensen, C.; Jensen, H.E.; Trebbien, R.; Webby, R.J.; Larsen, L.E. Avian and Human Influenza A Virus Receptors in Bovine Mammary Gland. Emerg. Infect. Dis. 2024, 30, 1907–1911. [Google Scholar] [CrossRef]
- Oguzie, J.U.; Marushchak, L.V.; Shittu, I.; Lednicky, J.A.; Miller, A.L.; Hao, H.; Nelson, M.I.; Gray, G.C. Avian Influenza A(H5N1) Virus among Dairy Cattle, Texas, USA. Emerg. Infect. Dis. 2024, 30, 1425–1429. [Google Scholar] [CrossRef]
- Cohen, J. Worries about bird flu in U.S. cattle intensify. Science 2024, 384, 12–13. [Google Scholar] [CrossRef]
- Kalthoff, D.; Hoffmann, B.; Harder, T.; Durban, M.; Beer, M. Experimental infection of cattle with highly pathogenic avian influenza virus (H5N1). Emerg. Infect. Dis. 2008, 14, 1132–1134. [Google Scholar] [CrossRef] [PubMed]
- Halwe, N.J.; Cool, K.; Breithaupt, A.; Schön, J.; Trujillo, J.D.; Nooruzzaman, M.; Kwon, T.; Ahrens, A.K.; Britzke, T.; McDowell, C.D.; et al. H5N1 clade 2.3.4.4b dynamics in experimentally infected calves and cows. Nature 2025, 637, 903–912. [Google Scholar] [CrossRef]
- Perez-Acle, T.; Ravello, C.; Rosemblatt, M. Are we cultivating the perfect storm for a human avian influenza pandemic? Biol. Res. 2024, 57, 96. [Google Scholar] [CrossRef]
- Gu, C.; Maemura, T.; Guan, L.; Eisfeld, A.J.; Biswas, A.; Kiso, M.; Uraki, R.; Ito, M.; Trifkovic, S.; Wang, T.; et al. A human isolate of bovine H5N1 is transmissible and lethal in animal models. Nature 2024, 636, 711–718. [Google Scholar] [CrossRef] [PubMed]
- WHO. Cumulative Number of Confirmed Human Cases for Avian Influenza A(H5N1) Reported to WHO, 2003–2024, 12 December 2024. Available online: https://www.who.int/publications/m/item/cumulative-number-of-confirmed-human-cases-for-avian-influenza-a(h5n1)-reported-to-who--2003-2024--20-december-2024 (accessed on 7 March 2025).
- CDC. First H5 Bird Flu Death Reported in United States. Available online: https://www.cdc.gov/media/releases/2025/m0106-h5-birdflu-death.html (accessed on 7 March 2025).
- Plaza, P.I.; Gamarra-Toledo, V.; Euguí, J.R.; Lambertucci, S.A. Recent Changes in Patterns of Mammal Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus Worldwide. Emerg. Infect. Dis. 2024, 30, 444–452. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: Rapid Testing for the Avian Influenza A(H5N1) Virus is Urgently Required as Infections in Poultry and Dairy Cows are on the Rise, and so is Transmission to Humans. Med. Sci. Monit. 2025, 31, e949109. [Google Scholar] [CrossRef]
- Guan, L.; Eisfeld, A.J.; Pattinson, D.; Gu, C.; Biswas, A.; Maemura, T.; Trifkovic, S.; Babujee, L.; Presler, R., Jr.; Dahn, R.; et al. Cow’s Milk Containing Avian Influenza A(H5N1) Virus—Heat Inactivation and Infectivity in Mice. N. Engl. J. Med. 2024, 391, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Louis, S.; Mark-Carew, M.; Biggerstaff, M.; Yoder, J.; Boehm, A.B.; Wolfe, M.K.; Flood, M.; Peters, S.; Stobierski, M.G.; Coyle, J.; et al. Wastewater Surveillance for Influenza A Virus and H5 Subtype Concurrent with the Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in Cattle and Poultry and Associated Human Cases—United States, 12 May 12–13 July 2024. MMWR Morb. Mortal. Wkly. Rep. 2024, 73, 804–809. [Google Scholar] [CrossRef]
- Koopmans, M.P.G.; Barton Behravesh, C.; Cunningham, A.A.; Adisasmito, W.B.; Almuhairi, S.; Bilivogui, P.; Bukachi, S.A.; Casas, N.; Cediel Becerra, N.; Charron, D.F.; et al. The panzootic spread of highly pathogenic avian influenza H5N1 sublineage 2.3.4.4b: A critical appraisal of One Health preparedness and prevention. Lancet Infect. Dis. 2024, 24, e774–e781. [Google Scholar] [CrossRef]
- Spackman, E.; Jones, D.R.; McCoig, A.M.; Colonius, T.J.; Goraichuk, I.V.; Suarez, D.L. Characterization of highly pathogenic avian influenza virus in retail dairy products in the US. J. Virol. 2024, 98, e0088124. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; McVeigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Kash, J.C. Influenza virus evolution, host adaptation, and pandemic formation. Cell Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Sheng, Z.; Huang, C.; Wang, D.; Li, F. Influenza D virus. Curr. Opin. Virol. 2020, 44, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017, 8, 1580–1591. [Google Scholar] [CrossRef]
- Sonnberg, S.; Webby, R.J.; Webster, R.G. Natural history of highly pathogenic avian influenza H5N1. Virus Res. 2013, 178, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef]
- Carter, T.; Iqbal, M. The Influenza A Virus Replication Cycle: A Comprehensive Review. Viruses 2024, 16, 316. [Google Scholar] [CrossRef]
- Te Velthuis, A.J.; Fodor, E. Influenza virus RNA polymerase: Insights into the mechanisms of viral RNA synthesis. Nat. Rev. Microbiol. 2016, 14, 479–493. [Google Scholar] [CrossRef]
- Lakadamyali, M.; Rust, M.J.; Zhuang, X. Endocytosis of influenza viruses. Microbes Infect. 2004, 6, 929–936. [Google Scholar] [CrossRef]
- Kash, J.C.; Goodman, A.G.; Korth, M.J.; Katze, M.G. Hijacking of the host-cell response and translational control during influenza virus infection. Virus Res. 2006, 119, 111–120. [Google Scholar] [CrossRef]
- Wagner, R.; Matrosovich, M.; Klenk, H.D. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev. Med. Virol. 2002, 12, 159–166. [Google Scholar] [CrossRef]
- Nayak, D.P.; Hui, E.K.; Barman, S. Assembly and budding of influenza virus. Virus Res. 2004, 106, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Luo, M. Influenza virus entry. Adv. Exp. Med. Biol. 2012, 726, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, S.; Blijleven, J.S.; Roos, W.H.; Onck, P.R.; van der Giessen, E.; van Oijen, A.M. Hemagglutinin-Mediated Membrane Fusion: A Biophysical Perspective. Annu. Rev. Biophys. 2018, 47, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Dadonaite, B.; Gilbertson, B.; Knight, M.L.; Trifkovic, S.; Rockman, S.; Laederach, A.; Brown, L.E.; Fodor, E.; Bauer, D.L.V. The structure of the influenza A virus genome. Nat. Microbiol. 2019, 4, 1781–1789. [Google Scholar] [CrossRef]
- Pleschka, S. Overview of influenza viruses. Curr. Top. Microbiol. Immunol. 2013, 370, 1–20. [Google Scholar] [CrossRef]
- PAHO. La Muerte Púrpura: La Gran Gripe de 1918. Available online: https://www.paho.org/es/quienes-somos/historia-ops/muerte-purpura-gran-gripe-1918 (accessed on 11 March 2025).
- WHO. Number of COVID-19 Deaths Reported to WHO (Cumulative Total). Available online: https://data.who.int/dashboards/covid19/deaths (accessed on 11 March 2025).
- Bett, B.; Henning, J.; Abdu, P.; Okike, I.; Poole, J.; Young, J.; Randolph, T.F.; Perry, B.D. Transmission rate and reproductive number of the H5N1 highly pathogenic avian influenza virus during the December 2005–July 2008 epidemic in Nigeria. Transbound. Emerg. Dis. 2014, 61, 60–68. [Google Scholar] [CrossRef]
- Ward, M.P.; Maftei, D.; Apostu, C.; Suru, A. Estimation of the basic reproductive number (R0) for epidemic, highly pathogenic avian influenza subtype H5N1 spread. Epidemiol. Infect. 2009, 137, 219–226. [Google Scholar] [CrossRef]
- Hermann, E.; Krammer, F. Clade 2.3.4.4b H5N1 neuraminidase has a long stalk, which is in contrast to most highly pathogenic H5N1 viruses circulating between 2002 and 2020. mBio 2025, 16, e0398924. [Google Scholar] [CrossRef]
- Bennett-Laso, B.; Berazay, B.; Muñoz, G.; Ariyama, N.; Enciso, N.; Braun, C.; Krüger, L.; Barták, M.; González-Aravena, M.; Neira, V. Confirmation of highly pathogenic avian influenza H5N1 in skuas, Antarctica 2024. Front. Vet. Sci. 2024, 11, 1423404. [Google Scholar] [CrossRef]
- Banyard, A.C.; Bennison, A.; Byrne, A.M.P.; Reid, S.M.; Lynton-Jenkins, J.G.; Mollett, B.; De Silva, D.; Peers-Dent, J.; Finlayson, K.; Hall, R.; et al. Detection and spread of high pathogenicity avian influenza virus H5N1 in the Antarctic Region. Nat. Commun. 2024, 15, 7433. [Google Scholar] [CrossRef] [PubMed]
- Lisovski, S.; Günther, A.; Dewar, M.; Ainley, D.; Aldunate, F.; Arce, R.; Ballard, G.; Bauer, S.; Belliure, J.; Banyard, A.C.; et al. Unexpected Delayed Incursion of Highly Pathogenic Avian Influenza H5N1 (Clade 2.3.4.4b) Into the Antarctic Region. Influenza Other Respir. Viruses 2024, 18, e70010. [Google Scholar] [CrossRef]
- Fasanmi, O.G.; Odetokun, I.A.; Balogun, F.A.; Fasina, F.O. Public health concerns of highly pathogenic avian influenza H5N1 endemicity in Africa. Vet. World 2017, 10, 1194–1204. [Google Scholar] [CrossRef]
- Tassoni, L.; Fusaro, A.; Milani, A.; Lemey, P.; Awuni, J.A.; Sedor, V.B.; Dogbey, O.; Commey, A.N.; Meseko, C.; Joannis, T.; et al. Genetically Different Highly Pathogenic Avian Influenza A(H5N1) Viruses in West Africa, 2015. Emerg. Infect. Dis. 2016, 22, 2132–2136. [Google Scholar] [CrossRef] [PubMed]
- Abolnik, C.; Phiri, T.; Peyrot, B.; de Beer, R.; Snyman, A.; Roberts, D.; Ludynia, K.; Jordaan, F.; Maartens, M.; Ismail, Z.; et al. The Molecular Epidemiology of Clade 2.3.4.4B H5N1 High Pathogenicity Avian Influenza in Southern Africa, 2021–2022. Viruses 2023, 15, 1383. [Google Scholar] [CrossRef]
- Abolnik, C.; Roberts, L.C.; Strydom, C.; Snyman, A.; Roberts, D.G. Outbreaks of H5N1 High Pathogenicity Avian Influenza in South Africa in 2023 Were Caused by Two Distinct Sub-Genotypes of Clade 2.3.4.4b Viruses. Viruses 2024, 16, 896. [Google Scholar] [CrossRef] [PubMed]
- Leguia, M.; Garcia-Glaessner, A.; Muñoz-Saavedra, B.; Juarez, D.; Barrera, P.; Calvo-Mac, C.; Jara, J.; Silva, W.; Ploog, K.; Amaro, L.; et al. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat. Commun. 2023, 14, 5489. [Google Scholar] [CrossRef]
- Bruno, A.; Alfaro-Núñez, A.; de Mora, D.; Armas, R.; Olmedo, M.; Garcés, J.; Vaca, M.S.; De la Torre, E.; Jarrin, D.; Burbano, L.; et al. Phylogenetic analysis reveals that the H5N1 avian influenza A outbreak in poultry in Ecuador in November 2022 is associated with the highly pathogenic clade 2.3.4.4b. Int. J. Infect. Dis. 2023, 133, 27–30. [Google Scholar] [CrossRef]
- Paternina, D.; Herazo, R.; Oviedo, M.; Mattar, S. Dramatic re-emergence of avian influenza in Colombia and Latin America. Travel Med. Infect. Dis. 2024, 59, 102711. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Brock, N.; Belser, J.A.; Sun, X.; Pappas, C.; Kieran, T.J.; Basu Thakur, P.; Zeng, H.; Cui, D.; Frederick, J.; et al. Highly pathogenic avian influenza A(H5N1) virus of clade 2.3.4.4b isolated from a human case in Chile causes fatal disease and transmits between co-housed ferrets. Emerg. Microbes Infect. 2024, 13, 2332667. [Google Scholar] [CrossRef]
- Kandeil, A.; Patton, C.; Jones, J.C.; Jeevan, T.; Harrington, W.N.; Trifkovic, S.; Seiler, J.P.; Fabrizio, T.; Woodard, K.; Turner, J.C.; et al. Rapid evolution of A(H5N1) influenza viruses after intercontinental spread to North America. Nat. Commun. 2023, 14, 3082. [Google Scholar] [CrossRef] [PubMed]
- Ly, H. Highly pathogenic avian influenza H5N1 virus infections of dairy cattle and livestock handlers in the United States of America. Virulence 2024, 15, 2343931. [Google Scholar] [CrossRef] [PubMed]
- Tawidian, P.; Torchetti, M.K.; Killian, M.L.; Lantz, K.; Dilione, K.E.; Ringenberg, J.M.; Bevins, S.N.; Lenoch, J.B.; Ip, H.S. Genotypic Clustering of H5N1 Avian Influenza Viruses in North America Evaluated by Ordination Analysis. Viruses 2024, 16, 1818. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Tellier, R.; Wong, S.; Li, Y.; Bastien, N.; Tang, J.W.; Drews, S.J.; Jang, Y.; Davis, C.T.; Fonseca, K.; et al. Full-genome analysis of avian influenza A(H5N1) virus from a human, North America, 2013. Emerg. Infect. Dis. 2014, 20, 887–891. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild-Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Engelsma, M.; Heutink, R.; Harders, F.; Germeraad, E.A.; Beerens, N. Multiple Introductions of Reassorted Highly Pathogenic Avian Influenza H5Nx Viruses Clade 2.3.4.4b Causing Outbreaks in Wild Birds and Poultry in The Netherlands, 2020–2021. Microbiol. Spectr. 2022, 10, e0249921. [Google Scholar] [CrossRef]
- Muzyka, D.; Rula, O.; Tkachenko, S.; Muzyka, N.; Köthe, S.; Pishchanskyi, O.; Stegniy, B.; Pantin-Jackwood, M.; Beer, M. Highly Pathogenic and Low Pathogenic Avian Influenza H5 Subtype Viruses in Wild Birds in Ukraine. Avian Dis. 2019, 63, 219–229. [Google Scholar] [CrossRef]
- King, J.; Harder, T.; Conraths, F.J.; Beer, M.; Pohlmann, A. The genetics of highly pathogenic avian influenza viruses of subtype H5 in Germany, 2006–2020. Transbound. Emerg. Dis. 2021, 68, 1136–1150. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Calle-Hernández, D.M.; Ulloque-Badaracco, J.R.; Alarcón-Braga, E.A.; Hernández-Bustamante, E.A.; Cabrera-Guzmán, J.C.; Quispe-Vasquez, S.M.; Huayta-Cortez, M.A.; Benites-Zapata, V.A.; Rodriguez-Morales, A.J. Highly pathogenic avian influenza A(H5N1) in animals: A systematic review and meta-analysis. New Microbes New Infect. 2024, 60–61, 101439. [Google Scholar] [CrossRef]
- Ariyama, N.; Pardo-Roa, C.; Muñoz, G.; Aguayo, C.; Ávila, C.; Mathieu, C.; Almonacid, L.I.; Medina, R.A.; Brito, B.; Johow, M.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus in Wild Birds, Chile. Emerg. Infect. Dis. 2023, 29, 1842–1845. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Siegers, J.Y.; Tan, S.; Sharp, B.; Freiden, P.; Johow, M.; Orozco, K.; Ruiz, S.; Baumberger, C.; Galdames, P.; et al. Detection and phylogenetic analysis of highly pathogenic A/H5N1 avian influenza clade 2.3.4.4b virus in Chile, 2022. Emerg. Microbes Infect. 2023, 12, 2220569. [Google Scholar] [CrossRef] [PubMed]
- Kutkat, O.; Gomaa, M.; Moatasim, Y.; El Taweel, A.; Kamel, M.N.; El Sayes, M.; GabAllah, M.; Kandeil, A.; McKenzie, P.P.; Webby, R.J.; et al. Highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b in wild rats in Egypt during 2023. Emerg. Microbes Infect. 2024, 13, 2396874. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: Concerns as Highly Pathogenic Avian Influenza (HPAI) Virus of the H5N1 Subtype is Identified in Dairy Cows and Other Mammals. Med. Sci. Monit. 2024, 30, e945315. [Google Scholar] [CrossRef]
- Szaluś-Jordanow, O.; Golke, A.; Dzieciątkowski, T.; Chrobak-Chmiel, D.; Rzewuska, M.; Czopowicz, M.; Sapierzyński, R.; Kardas, M.; Biernacka, K.; Mickiewicz, M.; et al. A Fatal A/H5N1 Avian Influenza Virus Infection in a Cat in Poland. Microorganisms 2023, 11, 2263. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.X.; Souchaud, F.; Pierre, I.; Beven, V.; Hirchaud, E.; Hérault, F.; Planel, R.; Rigaudeau, A.; Bernard-Stoecklin, S.; Van der Werf, S.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus in Domestic Cat, France, 2022. Emerg. Infect. Dis. 2023, 29, 1696–1698. [Google Scholar] [CrossRef] [PubMed]
- Chothe, S.K.; Srinivas, S.; Misra, S.; Nallipogu, N.C.; Gilbride, E.; LaBella, L.; Mukherjee, S.; Gauthier, C.H.; Pecoraro, H.L.; Webb, B.T.; et al. Marked neurotropism and potential adaptation of H5N1 clade 2.3.4.4.b virus in naturally infected domestic cats. Emerg. Microbes Infect. 2025, 14, 2440498. [Google Scholar] [CrossRef]
- Schmidt, P.H.; White, B.J.; Finley, A.; Bortoluzzi, E.M.; Depenbusch, B.E.; Mancke, M.; Brown, R.E.; Jensen, M.; Lancaster, P.A.; Larson, R.L. Determining Frequency of Common Pulmonary Gross and Histopathological Findings in Feedyard Fatalities. Vet. Sci. 2023, 10, 228. [Google Scholar] [CrossRef]
- Sillman, S.J.; Drozd, M.; Loy, D.; Harris, S.P. Naturally occurring highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b infection in three domestic cats in North America during 2023. J. Comp. Pathol. 2023, 205, 17–23. [Google Scholar] [CrossRef]
- Naraharisetti, R.; Weinberg, M.; Stoddard, B.; Stobierski, M.G.; Dodd, K.; Wineland, N.; Beal, M.; Morse, J.; Hatter, S.; Sledge, D.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection of Indoor Domestic Cats Within Dairy Industry Worker Households—Michigan, May 2024. MMWR. Morb. Mortal. Wkly. Rep. 2025, 74, 61–65. [Google Scholar] [CrossRef]
- Brown, J.D.; Black, A.; Haman, K.H.; Diel, D.G.; Ramirez, V.E.; Ziejka, R.S.; Fenelon, H.T.; Rabinowitz, P.M.; Stevens, L.; Poulson, R.; et al. Antibodies to Influenza A(H5N1) Virus in Hunting Dogs Retrieving Wild Fowl, Washington, USA. Emerg. Infect. Dis. 2024, 30, 1271–1274. [Google Scholar] [CrossRef]
- Maas, R.; Tacken, M.; Ruuls, L.; Koch, G.; van Rooij, E.; Stockhofe-Zurwieden, N. Avian influenza (H5N1) susceptibility and receptors in dogs. Emerg. Infect. Dis. 2007, 13, 1219–1221. [Google Scholar] [CrossRef]
- Chen, Y.; Zhong, G.; Wang, G.; Deng, G.; Li, Y.; Shi, J.; Zhang, Z.; Guan, Y.; Jiang, Y.; Bu, Z.; et al. Dogs are highly susceptible to H5N1 avian influenza virus. Virology 2010, 405, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Trujillo, J.D.; Carossino, M.; Lyoo, E.L.; McDowell, C.D.; Cool, K.; Matias-Ferreyra, F.S.; Jeevan, T.; Morozov, I.; Gaudreault, N.N.; et al. Pigs are highly susceptible to but do not transmit mink-derived highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b. Emerg. Microbes Infect. 2024, 13, 2353292. [Google Scholar] [CrossRef]
- Imai, M.; Herfst, S.; Sorrell, E.M.; Schrauwen, E.J.; Linster, M.; De Graaf, M.; Fouchier, R.A.; Kawaoka, Y. Transmission of influenza A/H5N1 viruses in mammals. Virus Res. 2013, 178, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Li, T.; Wen, Z.; Wu, S.; Wang, Z.; Zheng, J.; Chen, M.; Chen, F.; Wei, W.K.; Zhai, S.L.; et al. Identification of D/Yama2019 Lineage-Like Influenza D Virus in Chinese Cattle. Front. Vet. Sci. 2022, 9, 939456. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.; Thomas, M.; Sheng, Z.; Hause, B.M.; Collin, E.A.; Knudsen, D.E.; Pillatzki, A.; Nelson, E.; Wang, D.; Kaushik, R.S.; et al. Replication and Transmission of the Novel Bovine Influenza D Virus in a Guinea Pig Model. J. Virol. 2015, 89, 11990–12001. [Google Scholar] [CrossRef]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza D virus infection in Mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef]
- Flynn, O.; Gallagher, C.; Mooney, J.; Irvine, C.; Ducatez, M.; Hause, B.; McGrath, G.; Ryan, E. Influenza D Virus in Cattle, Ireland. Emerg. Infect. Dis. 2018, 24, 389–391. [Google Scholar] [CrossRef]
- Collin, E.A.; Sheng, Z.; Lang, Y.; Ma, W.; Hause, B.M.; Li, F. Cocirculation of two distinct genetic and antigenic lineages of proposed influenza D virus in cattle. J. Virol. 2015, 89, 1036–1042. [Google Scholar] [CrossRef]
- Silveira, S.; Falkenberg, S.M.; Kaplan, B.S.; Crossley, B.; Ridpath, J.F.; Bauermann, F.B.; Fossler, C.P.; Dargatz, D.A.; Dassanayake, R.P.; Vincent, A.L.; et al. Serosurvey for Influenza D Virus Exposure in Cattle, United States, 2014–2015. Emerg. Infect. Dis. 2019, 25, 2074–2080. [Google Scholar] [CrossRef]
- Quast, M.; Sreenivasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza D virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Pelletier, C.; Meyer, G. Influenza D virus in cattle, France, 2011–2014. Emerg. Infect. Dis. 2015, 21, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, C.J.; Oliva, J.; Pauly, M.; Losch, S.; Wildschutz, F.; Muller, C.P.; Hübschen, J.M.; Ducatez, M.F. Influenza D Virus Circulation in Cattle and Swine, Luxembourg, 2012–2016. Emerg. Infect. Dis. 2018, 24, 1388–1389. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Mosena, A.C.S.; Baumbach, L.; Demoliner, M.; Gularte, J.S.; Pavarini, S.P.; Driemeier, D.; Weber, M.N.; Spilki, F.R.; Canal, C.W. Cattle influenza D virus in Brazil is divergent from established lineages. Arch. Virol. 2022, 167, 1181–1184. [Google Scholar] [CrossRef]
- CDC. Situación Actual: Influenza Aviar en Vacas Lecheras. Available online: https://espanol.cdc.gov/bird-flu/situation-summary/mammals.html#:~:text=El%201%20de%20abril%2C%20los,(H5N1)%20de%20la%20HPAI (accessed on 13 March 2025).
- Goujgoulova, G.; Koev, K. Risk Assessment of Spread of the Influenza A Virus in Cows in South Bulgaria. Viruses 2025, 17, 246. [Google Scholar] [CrossRef]
- Rubin, E.J.; Baden, L.R.; Goldstein, R.; Shuford, J.A.; Morrissey, S. NEJM Outbreaks Update—H5N1: A View from the States. N. Engl. J. Med. 2025, 392, e37. [Google Scholar] [CrossRef]
- Hawman, D.W.; Tipih, T.; Hodge, E.; Stone, E.T.; Warner, N.; McCarthy, N.; Granger, B.; Meade-White, K.; Leventhal, S.; Hatzakis, K.; et al. Clade 2.3.4.4b but not historical clade 1 HA replicating RNA vaccine protects against bovine H5N1 challenge in mice. Nat. Commun. 2025, 16, 655. [Google Scholar] [CrossRef]
- Alexakis, L.; Buczkowski, H.; Ducatez, M.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Ståhl, K.; Staubach, C.; Svartström, O.; Terregino, C.; et al. Avian influenza overview September-December 2024. Efsa J. 2025, 23, e9204. [Google Scholar] [CrossRef]
- Mostafa, A.; Barre, R.S.; Allué-Guardia, A.; Escobedo, R.A.; Shivanna, V.; Rothan, H.; Castro, E.M.; Ma, Y.; Cupic, A.; Jackson, N.; et al. Replication kinetics, pathogenicity and virus-induced cellular responses of cattle-origin influenza A(H5N1) isolates from Texas, United States. Emerg. Microbes Infect. 2025, 14, 2447614. [Google Scholar] [CrossRef]
- Nguyen, T.-Q.; Hutter, C.R.; Markin, A.; Thomas, M.; Lantz, K.; Killian, M.L.; Janzen, G.M.; Vijendran, S.; Wagle, S.; Inderski, B.; et al. Emergence and interstate spread of highly pathogenic avian influenza A(H5N1) in dairy cattle in the United States. Science 2025, 388, eadq0900. [Google Scholar] [CrossRef]
- Song, H.; Hao, T.; Han, P.; Wang, H.; Zhang, X.; Li, X.; Wang, Y.; Chen, J.; Li, Y.; Jin, X.; et al. Receptor binding, structure, and tissue tropism of cattle-infecting H5N1 avian influenza virus hemagglutinin. Cell 2025, 188, 919–929.e919. [Google Scholar] [CrossRef]
- Imai, M.; Ueki, H.; Ito, M.; Iwatsuki-Horimoto, K.; Kiso, M.; Biswas, A.; Trifkovic, S.; Cook, N.; Halfmann, P.J.; Neumann, G.; et al. Highly pathogenic avian H5N1 influenza A virus replication in ex vivo cultures of bovine mammary gland and teat tissues. Emerg. Microbes Infect. 2025, 14, 2450029. [Google Scholar] [CrossRef] [PubMed]
- Crossley, B.M.; Miramontes, C.C.; Rejmanek, D.; Gallardo, R.; Pereira, R. In-laboratory inactivation of H5N1 in raw whole milk through milk acidification: Results from a pilot study. J. Dairy. Sci. 2025, 108, 2264–2275. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, F.; Cardenas, S.; Yinda, K.C.; Mukesh, R.K.; Ochwoto, M.; Gallogly, S.; Wickenhagen, A.; Bibby, K.; de Wit, E.; Morris, D.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Stability in Irradiated Raw Milk and Wastewater and on Surfaces, United States. Emerg. Infect. Dis. 2025, 31, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Le Sage, V.; Campbell, A.J.; Reed, D.S.; Duprex, W.P.; Lakdawala, S.S. Persistence of Influenza H5N1 and H1N1 Viruses in Unpasteurized Milk on Milking Unit Surfaces. Emerg. Infect. Dis. 2024, 30, 1721–1723. [Google Scholar] [CrossRef]
- Schafers, J.; Warren, C.J.; Yang, J.; Zhang, J.; Cole, S.J.; Cooper, J.; Drewek, K.; Kolli, B.R.; McGinn, N.; Qureshi, M.; et al. Pasteurisation temperatures effectively inactivate influenza A viruses in milk. Nat. Commun. 2025, 16, 1173. [Google Scholar] [CrossRef]
- Falender, R.; Radniecki, T.S.; Kelly, C.; Cieslak, P.; Mickle, D.; Hall, H.; Scholz, R.; Sutton, M. Avian Influenza A(H5) Subtype in Wastewater—Oregon, September 15, 2021–July 11, 2024. MMWR Morb. Mortal. Wkly. Rep. 2025, 74, 102–106. [Google Scholar] [CrossRef]
- Verma, A.K.; Kumar, M.; Murugkar, H.V.; Nagarajan, S.; Tosh, C.; Namdeo, P.; Singh, R.; Mishra, S.; Senthilkumar, D.; Singh, V.P.; et al. Highly pathogenic avian influenza (H5N1) infection in crows through ingestion of infected crow carcasses. Microb. Pathog. 2023, 183, 106330. [Google Scholar] [CrossRef]
- Hall, J.S.; Franson, J.C.; Gill, R.E.; Meteyer, C.U.; TeSlaa, J.L.; Nashold, S.; Dusek, R.J.; Ip, H.S. Experimental challenge and pathology of highly pathogenic avian influenza virus H5N1 in dunlin (Calidris alpina), an intercontinental migrant shorebird species. Influenza Other Respir. Viruses 2011, 5, 365–372. [Google Scholar] [CrossRef]
- Takadate, Y.; Tsunekuni, R.; Kumagai, A.; Mine, J.; Kikutani, Y.; Sakuma, S.; Miyazawa, K.; Uchida, Y. Different Infectivity and Transmissibility of H5N8 and H5N1 High Pathogenicity Avian Influenza Viruses Isolated from Chickens in Japan in the 2021/2022 Season. Viruses 2023, 15, 265. [Google Scholar] [CrossRef]
- Vahlenkamp, T.W.; Teifke, J.P.; Harder, T.C.; Beer, M.; Mettenleiter, T.C. Systemic influenza virus H5N1 infection in cats after gastrointestinal exposure. Influenza Other Respir. Viruses 2010, 4, 379–386. [Google Scholar] [CrossRef]
- Arruda, B.; Baker, A.L.V.; Buckley, A.; Anderson, T.K.; Torchetti, M.; Bergeson, N.H.; Killian, M.L.; Lantz, K. Divergent Pathogenesis and Transmission of Highly Pathogenic Avian Influenza A(H5N1) in Swine. Emerg. Infect. Dis. 2024, 30, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Graaf, A.; Piesche, R.; Sehl-Ewert, J.; Grund, C.; Pohlmann, A.; Beer, M.; Harder, T. Low Susceptibility of Pigs against Experimental Infection with HPAI Virus H5N1 Clade 2.3.4.4b. Emerg. Infect. Dis. 2023, 29, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Abente, E.J.; Kitikoon, P.; Lager, K.M.; Gauger, P.C.; Anderson, T.K.; Vincent, A.L. A highly pathogenic avian-derived influenza virus H5N1 with 2009 pandemic H1N1 internal genes demonstrates increased replication and transmission in pigs. J. Gen. Virol. 2017, 98, 18–30. [Google Scholar] [CrossRef]
- Donatelli, I.; Castrucci, M.R.; De Marco, M.A.; Delogu, M.; Webster, R.G. Human-Animal Interface: The Case for Influenza Interspecies Transmission. Adv. Exp. Med. Biol. 2017, 972, 17–33. [Google Scholar] [CrossRef]
- Meseko, C.; Globig, A.; Ijomanta, J.; Joannis, T.; Nwosuh, C.; Shamaki, D.; Harder, T.; Hoffman, D.; Pohlmann, A.; Beer, M.; et al. Evidence of exposure of domestic pigs to Highly Pathogenic Avian Influenza H5N1 in Nigeria. Sci. Rep. 2018, 8, 5900. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Dong, L.; Xin, L.; Lan, Y.; Chen, Y.; Yang, L.; Shu, Y. Human avian influenza A (H5N1) virus infection in China. Sci. China C Life Sci. 2009, 52, 407–411. [Google Scholar] [CrossRef]
- Galli, M.; Giacomelli, A.; Lai, A.; Zehender, G. H5N1 influenza A virus: Lessons from past outbreaks and emerging threats. Infez. Med. 2025, 33, 76–89. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Milton, S.; Abdul Hamid, C.; Reinoso Webb, C.; Presley, S.M.; Shetty, V.; Rollo, S.N.; Martinez, D.L.; Rai, S.; Gonzales, E.R.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection in a Dairy Farm Worker. N. Engl. J. Med. 2024, 390, 2028–2029. [Google Scholar] [CrossRef]
- Le Sage, V.; Werner, B.D.; Merrbach, G.A.; Petnuch, S.E.; O’Connell, A.K.; Simmons, H.C.; McCarthy, K.R.; Reed, D.S.; Moncla, L.H.; Bhavsar, D.; et al. Influenza A(H5N1) Immune Response among Ferrets with Influenza A(H1N1)pdm09 Immunity. Emerg. Infect. Dis. 2025, 31, 477–487. [Google Scholar] [CrossRef]
- Leonard, J.; Harker, E.J.; Szablewski, C.M.; Margrey, S.F.; Fred Gingrich, K., II; Crossley, K.; Fletcher, E.; McCreavy, C.J.; Weis-Torres, S.; Wang, D.; et al. Seroprevalence of Highly Pathogenic Avian Influenza A(H5) Virus Infections Among Bovine Veterinary Practitioners—United States, September 2024. MMWR Morb. Mortal. Wkly. Rep. 2025, 74, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Bird flu: US reports first human death in person infected with H5N1. BMJ 2025, 388, r28. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.; Coyle, J.; Mikesell, L.; Stoddard, B.; Eckel, S.; Weinberg, M.; Kuo, J.; Riner, D.; Margulieux, K.; Stricklen, J.; et al. Influenza A(H5N1) Virus Infection in Two Dairy Farm Workers in Michigan. N. Engl. J. Med. 2024, 391, 963–964. [Google Scholar] [CrossRef] [PubMed]
- Pulit-Penaloza, J.A.; Belser, J.A.; Brock, N.; Kieran, T.J.; Sun, X.; Pappas, C.; Zeng, H.; Carney, P.; Chang, J.; Bradley-Ferrell, B.; et al. Transmission of a human isolate of clade 2.3.4.4b A(H5N1) virus in ferrets. Nature 2024, 636, 705–710. [Google Scholar] [CrossRef]
- Teitelbaum, C.S.; Masto, N.M.; Sullivan, J.D.; Keever, A.C.; Poulson, R.L.; Carter, D.L.; Blake-Bradshaw, A.G.; Highway, C.J.; Feddersen, J.C.; Hagy, H.M.; et al. North American wintering mallards infected with highly pathogenic avian influenza show few signs of altered local or migratory movements. Sci. Rep. 2023, 13, 14473. [Google Scholar] [CrossRef]
- Kheirallah, K.A.; Al-Mistarehi, A.H.; Alsawalha, L.; Hijazeen, Z.; Mahrous, H.; Sheikali, S.; Al-Ramini, S.; Maayeh, M.; Dodeen, R.; Farajeh, M.; et al. Prioritizing zoonotic diseases utilizing the One Health approach: Jordan’s experience. One Health 2021, 13, 100262. [Google Scholar] [CrossRef]
- Enkirch, T.; Gervelmeyer, A.; Hallmaier-Wacker, L.; Melidou, A.; Willgert, K. Coordinated One Health investigation and management of outbreaks in humans and animals caused by zoonotic avian influenza viruses. Efsa J. 2025, 23, e9183. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | 2003–2009 | 2010–2014 | 2015–2019 | 2020 | 2021 | 2022 | 2023 | 2024 | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | Cases | Deaths | |
| Australia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 |
| Azerbaijan | 8 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 5 |
| Bangladesh | 1 | 0 | 6 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 1 |
| Cambodia | 9 | 7 | 47 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 4 | 10 | 2 | 72 | 43 |
| Canada | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 1 |
| Chile | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| China | 38 | 25 | 9 | 5 | 6 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 56 | 32 |
| Djibouti | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Ecuador | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Egypt | 90 | 27 | 120 | 50 | 149 | 43 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 359 | 120 |
| India | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Indonesia | 162 | 134 | 35 | 31 | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 200 | 168 |
| Iraq | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 2 |
| Lao | 2 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 2 |
| Myanmar | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Nepal | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Nigeria | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| Pakistan | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 |
| Spain | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Thailand | 25 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 17 |
| Turkey | 12 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 4 |
| UK | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 5 | 0 |
| USA | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 58 | 0 | 59 | 0 |
| Vietnam | 112 | 57 | 15 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 129 | 65 |
| Total | 468 | 282 | 233 | 125 | 160 | 48 | 1 | 0 | 2 | 1 | 6 | 1 | 12 | 4 | 72 | 3 | 954 | 464 |
| Domain | Challenges | Strategies |
|---|---|---|
| Surveillance |
|
|
| Diagnostics |
|
|
| Control |
|
|
| Prevention |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Rojas, I.C.; Bonilla-Aldana, D.K.; Solarte-Jimenez, C.L.; Bonilla-Aldana, J.L.; Acosta-España, J.D.; Rodriguez-Morales, A.J. Highly Pathogenic Avian Influenza (H5N1) Clade 2.3.4.4b in Cattle: A Rising One Health Concern. Animals 2025, 15, 1963. https://doi.org/10.3390/ani15131963
Sanchez-Rojas IC, Bonilla-Aldana DK, Solarte-Jimenez CL, Bonilla-Aldana JL, Acosta-España JD, Rodriguez-Morales AJ. Highly Pathogenic Avian Influenza (H5N1) Clade 2.3.4.4b in Cattle: A Rising One Health Concern. Animals. 2025; 15(13):1963. https://doi.org/10.3390/ani15131963
Chicago/Turabian StyleSanchez-Rojas, Ivan Camilo, D. Katterine Bonilla-Aldana, Catherin Lorena Solarte-Jimenez, Jorge Luis Bonilla-Aldana, Jaime David Acosta-España, and Alfonso J. Rodriguez-Morales. 2025. "Highly Pathogenic Avian Influenza (H5N1) Clade 2.3.4.4b in Cattle: A Rising One Health Concern" Animals 15, no. 13: 1963. https://doi.org/10.3390/ani15131963
APA StyleSanchez-Rojas, I. C., Bonilla-Aldana, D. K., Solarte-Jimenez, C. L., Bonilla-Aldana, J. L., Acosta-España, J. D., & Rodriguez-Morales, A. J. (2025). Highly Pathogenic Avian Influenza (H5N1) Clade 2.3.4.4b in Cattle: A Rising One Health Concern. Animals, 15(13), 1963. https://doi.org/10.3390/ani15131963