
Serum Concentrations of Imidazole Dipeptides and Serum Amyloid A in a Bottlenose Dolphin (Tursiops truncatus) with Rhabdomyolysis: Potential Biomarkers for Muscular Damage

 , and
, and {kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Case Description
2.2. Blood Sample Collection
2.3. Hematological and Biochemical Analyses
2.4. SAA Analysis
2.5. IDPs Analysis
2.6. Statistical Analysis
3. Results
3.1. Changes in IDPs, CK, and AST Levels
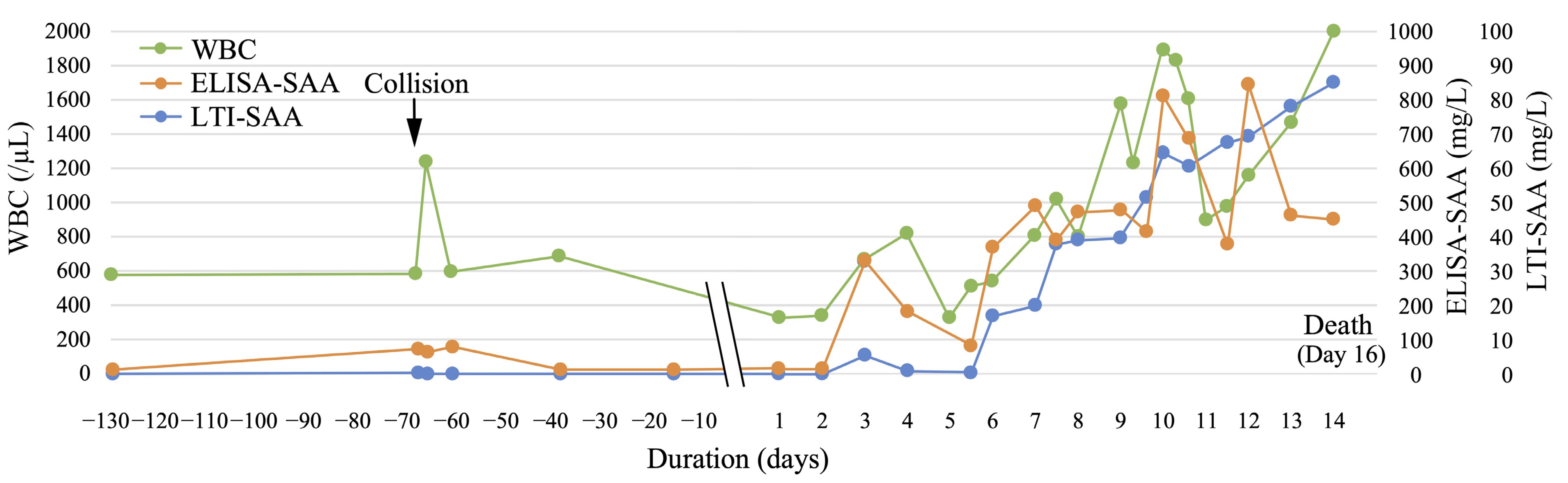
3.2. Changes in SAA and WBC Levels
3.3. Relationship Between IDPs and Other Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CK | creatinine kinase |
| AST | aspartate aminotransferase |
| AKI | acute kidney injury |
| IDP | imidazole dipeptide |
| SAA | serum amyloid A |
| WBC | white blood cell |
| ELISA | enzyme-linked immunosorbent assay |
| LTI | latex turbidimetric immunoassay |
References
- Torres, P.A.; Helmstetter, J.A.; Kaye, A.M.; Kaye, A.D. Rhabdomyolysis: Pathogenesis, diagnosis, and treatment. Ochsner J. 2015, 15, 58–69. [Google Scholar]
- Cabral, B.M.I.; Edding, S.N.; Portocarrero, J.P.; Lerma, E.V. Rhabdomyolysis. Dis. Mon. 2020, 66, 101015. [Google Scholar] [CrossRef]
- Yang, B.F.; Li, D.; Liu, C.L.; Luo, Y.; Shi, J.; Guo, X.Q.; Fan, H.J.; Lv, Q. Advances in rhabdomyolysis: A review of pathogenesis, diagnosis, and treatment. Chin. J. Traumatol. 2025; in press. [Google Scholar] [CrossRef]
- Castellini, M.A.; Somero, G.N. Buffering capacity of vertebrate muscle: Correlations with potentials for anaerobic function. J. Comp. Physiol. 1981, 143, 191–198. [Google Scholar] [CrossRef]
- Mirceta, S.; Signore, A.V.; Burns, J.M.; Cossins, A.R.; Campbell, K.L.; Berenbrink, M. Evolution of mammalian diving capacity traced by myoglobin net surface charge. Science 2013, 340, 1234192. [Google Scholar] [CrossRef]
- Noren, S.R.; Williams, T.M. Body size and skeletal muscle myoglobin of cetaceans: Adaptations for maximizing dive duration. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2000, 126, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Zager, R.A.; Burkhart, K. Myoglobin toxicity in proximal human kidney cells: Roles of Fe, Ca2+, H2O2, and terminal mitochondrial electron transport. Kidney Int. 1997, 51, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Herráez, P.; Sierra, E.; Arbelo, M.; Jaber, J.R.; de los Monteros, A.E.; Fernández, A. Rhabdomyolysis and myoglobinuric nephrosis (capture myopathy) in a striped dolphin. J. Wildl. Dis. 2007, 43, 770–774. [Google Scholar] [CrossRef]
- Herráez, P.; Espinosa de los Monteros, A.; Fernández, A.; Edwards, J.F.; Sacchini, S.; Sierra, E. Capture myopathy in live-stranded cetaceans. Vet. J. 2013, 196, 181–188. [Google Scholar] [CrossRef]
- Bonsembiante, F.; Centelleghe, C.; Rossi, G.; Giglio, S.; Madeo, E.; Gelain, M.E.; Mazzariol, S. Clinico-pathological findings in a striped dolphin (Stenella coeruleoalba) affected by rhabdomyolysis and myoglobinuric nephrosis (capture myopathy). J. Vet. Med. Sci. 2017, 79, 1013–1018. [Google Scholar] [CrossRef]
- Câmara, N.; Sierra, E.; Fernández, A.; Suárez-Santana, C.M.; Puig-Lozano, R.; Arbelo, M.; Herráez, P. Skeletal and cardiac rhabdomyolysis in a live-stranded neonatal Bryde’s whale with fetal distress. Front. Vet. Sci. 2019, 6, 476. [Google Scholar]
- Carvalho, V.L.; Viana, D.A.; Damasceno, A.A.P.; Monteiro, M.V.B.; Monteiro, F.O.B. Capture myopathy in a wild Clymene dolphin (Stenella clymene) stranded alive on the coast of Ceara State, Brazil. Braz. J. Vet. Med. 2025, 47, e000625. [Google Scholar] [CrossRef] [PubMed]
- Nueangphuet, P.; Hamano, T.; Hirai, T.; Sakaguchi, Y.; Sonoda, H.; Otsuka, M.; Yamato, O.; Hobo, S.; Ikeda, M.; Yamaguchi, R. Rhabdomyolysis, myoglobinuric nephrosis, and crystalline nephropathy in a captive bottlenose dolphin. J. Vet. Diagn. Investig. 2022, 34, 668–673. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Berg, L.C.; Thomsen, P.D.; Andersen, P.H.; Jensen, H.E.; Jacobsen, S. Serum amyloid A is expressed in histologically normal tissues from horses and cattle. Vet. Immunol. Immunopathol. 2011, 144, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Sack, G.H. Serum amyloid A (SAA) proteins. In Vertebrate and Invertebrate Respiratory Proteins, Lipoproteins and Other Body Fluid Proteins; Hoeger, U., Harris, J.R., Eds.; Subcellular Biochemistry; Springer International Publishing: Cham, Switzerland, 2020; pp. 421–436. [Google Scholar]
- Flower, J.E.; Langan, J.N.; Wells, R.S.; Cray, C.; Arheart, K.; Chinnadurai, S.K.; Adkesson, M.J. Serum acute-phase proteins in bottlenose dolphins (Tursiops truncatus) and correlation with commonly utilized inflammatory indices. J. Zoo Wildl. Med. 2020, 51, 657–662. [Google Scholar] [CrossRef]
- Anderson, R.; Waayers, R.; Knight, A. Orca behavior and subsequent aggression associated with oceanarium confinement. Animals 2016, 6, 49. [Google Scholar] [CrossRef]
- Dudzinski, K.D.; Hill, H.M.M. Stakeholder perspectives regarding captive marine mammals—How can we compromise? Zoo Biol. 2025, 44, 211–220. [Google Scholar] [CrossRef]
- Yamadate, S.; Nakayama, T. The future topics of discussion on JSCC recommended methods. Rinsho Byori 2016, 64, 544–549. [Google Scholar]
- Cray, C.; Arheart, K.L.; Hunt, M.; Clauss, T.; Leppert, L.L.; Roberts, K.; McCulloch, S.D.; Goldstein, J.D.; Gonzalez, C.; Sweeney, J.; et al. Acute phase protein quantitation in serum samples from healthy Atlantic bottlenose dolphins (Tursiops truncatus). J. Vet. Diagn. Investig. 2013, 25, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Kiemle, J.; Hindenberg, S.; Bauer, N.; Roecken, M. Comparison of a point-of-care serum amyloid A analyzer frequently used in equine practice with 2 turbidimetric immunoassays used in human and veterinary medicine. J. Vet. Diagn. Investig. 2022, 34, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, M.; Kato, S.; Arakawa, N.; Otsuka, M.; Hamano, T.; Kashiwagi, N.; Yabuki, A.; Yamato, O. Quantification of histidine-containing dipeptides in dolphin serum using a reversed-phase ion-pair high-performance liquid chromatography method. Separations 2021, 8, 128. [Google Scholar] [CrossRef]
- Ostrowski, P.; Bonczar, M.; Avram, A.E.; Lippi, G.; Henry, B.M. Safety monitoring of drug-induced muscle injury and rhabdomyolysis: A biomarker-guided approach for clinical practice and drug trials. Clin. Chem. Lab. Med. 2023, 61, 1688–1699. [Google Scholar] [CrossRef]
- Zhang, M.H. Rhabdomyolosis and its pathogenesis. World J. Emerg. Med. 2012, 3, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Dunnett, M.; Harris, R.C.; Dunnett, C.E.; Harris, P.A. Plasma carnosine concentration: Diurnal variation and effects of age, exercise and muscle damage. Equine Vet. J. 2002, 34, 283–287. [Google Scholar] [CrossRef]
- Bazzano, M.; Marchegiani, A.; Troisi, A.; McLean, A.; Laus, F. Serum amyloid A as a promising biomarker in domestic animals’ reproduction: Current knowledge and future perspective. Animals 2022, 12, 589. [Google Scholar] [CrossRef]
- Gelain, M.E.; Bonsembiante, F. Acute phase proteins in marine mammals: State of art, perspectives and challenges. Front. Immunol. 2019, 10, 1220. [Google Scholar] [CrossRef]
- Christense, M.; Jacobsen, S.; Ichiyanagi, T.; Kjelgaard-Hansen, M. Evaluation of an automated assay based on monoclonal anti-human serum amyloid A (SAA) antibodies for measurement of canine, feline, and equine SAA. Vet. J. 2012, 194, 332–337. [Google Scholar] [CrossRef]
- Yuki, M.; Inden, T.; Hirano, T.; Naito, E.; Taira, H.; Yokota, S.; Narita, M. Comparison of polyclonal and monoclonal antibody assays for serum amyloid A in cats: A study based on an automated turbidimetric immunoassay in a primary care veterinary hospital. Am. J. Vet. Res. 2024, 85. [Google Scholar] [CrossRef]
- Cray, C.; Knibb, R.I.; Knibb, J.R. Serum amyloid A and plasma protein electrophoresis fractions in farmed white-tailed deer. J. Vet. Diagn. Investig. 2019, 31, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nango, H.; Ushijima, M.; Takashima, M.; Nakamoto, M.; Matsutomo, T.; Jikihara, H.; Arakawa, N.; Maki, S.; Yabuki, A.; et al. Therapeutic effect of aged garlic extract on gingivitis in dogs. Front. Vet. Sci. 2023, 10, 1277272. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.N.; Petersen, M.B.; Capion, N.; Lundsgaard, J.F.H.; Jensen, A.L. Performance of an automated immunoturbidimetric assay for bovine serum amyloid A. Vet. Clin. Pathol. 2024, 53, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Shinya, U.; Yamato, O.; Iwamura, Y.; Kato, T.; Hamada, Y.; Widodo, O.S.; Taniguchi, M.; Takag, M. Serum amyloid A concentrations in young Japanese Black cattle: Relationship with colostrum intake and establishing cut-off concentrations. Animals 2025, 15, 1239. [Google Scholar] [CrossRef] [PubMed]
- Ijiri, M.; Akioka, K.; Kitano, T.; Miura, H.; Ono, H.K.; Terashima, R.; Fujimoto, Y.; Matsuo, T.; Yamato, O.; Kawaguchi, H. Acupuncture treatment improves transport stress in microminipigs through the acupoint in ears. In Vivo 2023, 37, 2100–2104. [Google Scholar] [CrossRef]
- Lennox, A.; Asahi, Y.; Arheart, K.; Ichiyanagi, T.; Cray, C. Preliminary evaluation of an immunoturbidimetric assay and lateral flow device for the measurement of serum amyloid A in rabbits. J. Exotic Pet Med. 2020, 33, 54–56. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arakawa, N.; Otsuka, M.; Hamano, T.; Kumagai, M.; Kato, S.; Hirai, T.; Yabuki, A.; Yamato, O. Serum Concentrations of Imidazole Dipeptides and Serum Amyloid A in a Bottlenose Dolphin (Tursiops truncatus) with Rhabdomyolysis: Potential Biomarkers for Muscular Damage. Animals 2025, 15, 1950. https://doi.org/10.3390/ani15131950
Arakawa N, Otsuka M, Hamano T, Kumagai M, Kato S, Hirai T, Yabuki A, Yamato O. Serum Concentrations of Imidazole Dipeptides and Serum Amyloid A in a Bottlenose Dolphin (Tursiops truncatus) with Rhabdomyolysis: Potential Biomarkers for Muscular Damage. Animals. 2025; 15(13):1950. https://doi.org/10.3390/ani15131950
Chicago/Turabian StyleArakawa, Nanami, Mika Otsuka, Takahisa Hamano, Momochika Kumagai, Sanae Kato, Takuya Hirai, Akira Yabuki, and Osamu Yamato. 2025. "Serum Concentrations of Imidazole Dipeptides and Serum Amyloid A in a Bottlenose Dolphin (Tursiops truncatus) with Rhabdomyolysis: Potential Biomarkers for Muscular Damage" Animals 15, no. 13: 1950. https://doi.org/10.3390/ani15131950
APA StyleArakawa, N., Otsuka, M., Hamano, T., Kumagai, M., Kato, S., Hirai, T., Yabuki, A., & Yamato, O. (2025). Serum Concentrations of Imidazole Dipeptides and Serum Amyloid A in a Bottlenose Dolphin (Tursiops truncatus) with Rhabdomyolysis: Potential Biomarkers for Muscular Damage. Animals, 15(13), 1950. https://doi.org/10.3390/ani15131950