
The Oral Microbiome in Queensland Free-Ranging Koalas (Phascolarctos cinereus) and Its Association with Age and Periodontal Disease

 ,
,  , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animal Selection and Age Determination
2.3. Clinical Examination and Sample Collection
2.4. DNA Extraction and Verification
2.5. 16S rRNA Gene Amplification, Library Preparation and Sequencing
2.6. Taxonomic Assignment and OTU Clustering
2.7. Statistical Analysis
3. Results
3.1. General and Oral Health
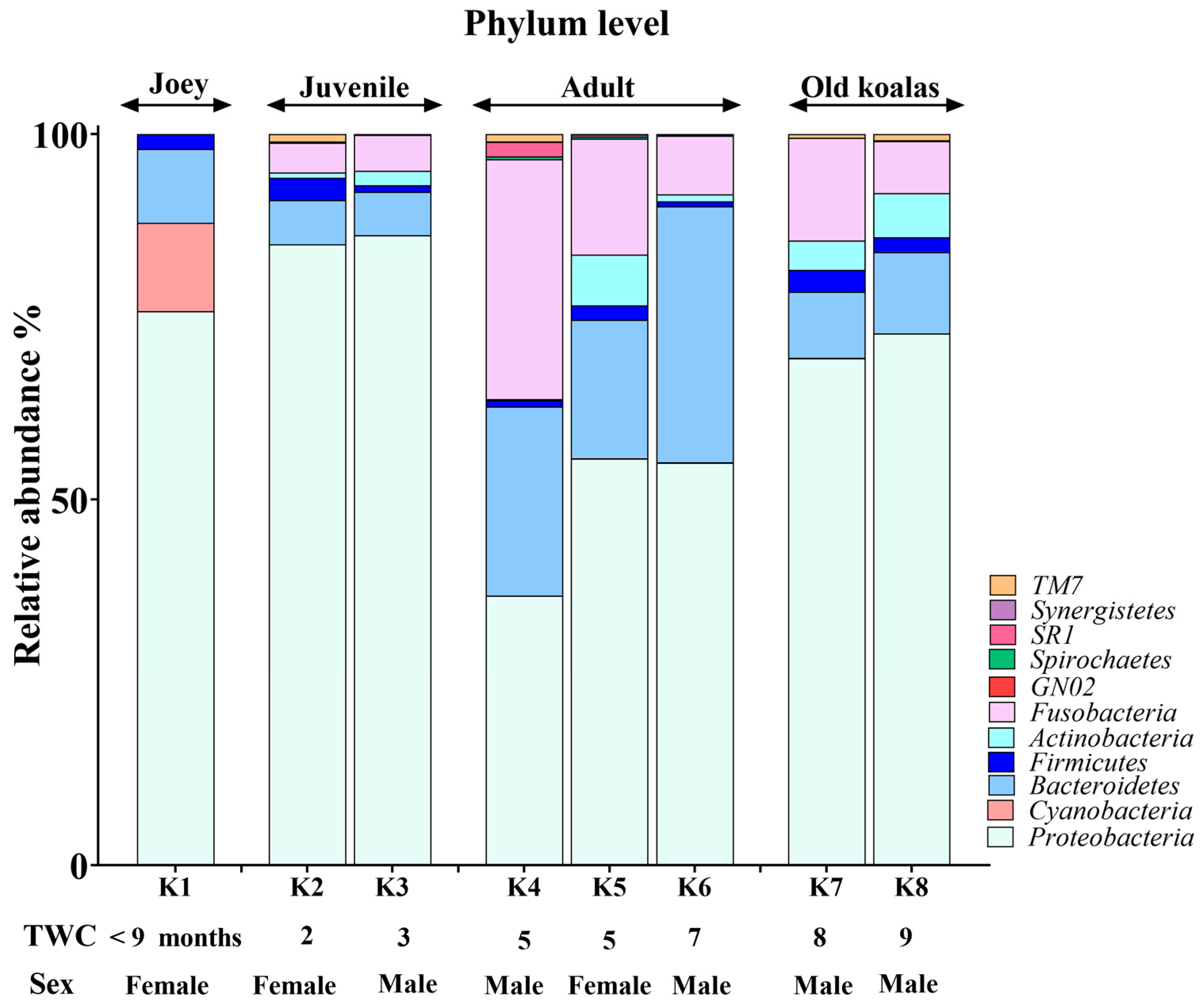
3.2. Phylum-Level Oral Microbiome Composition
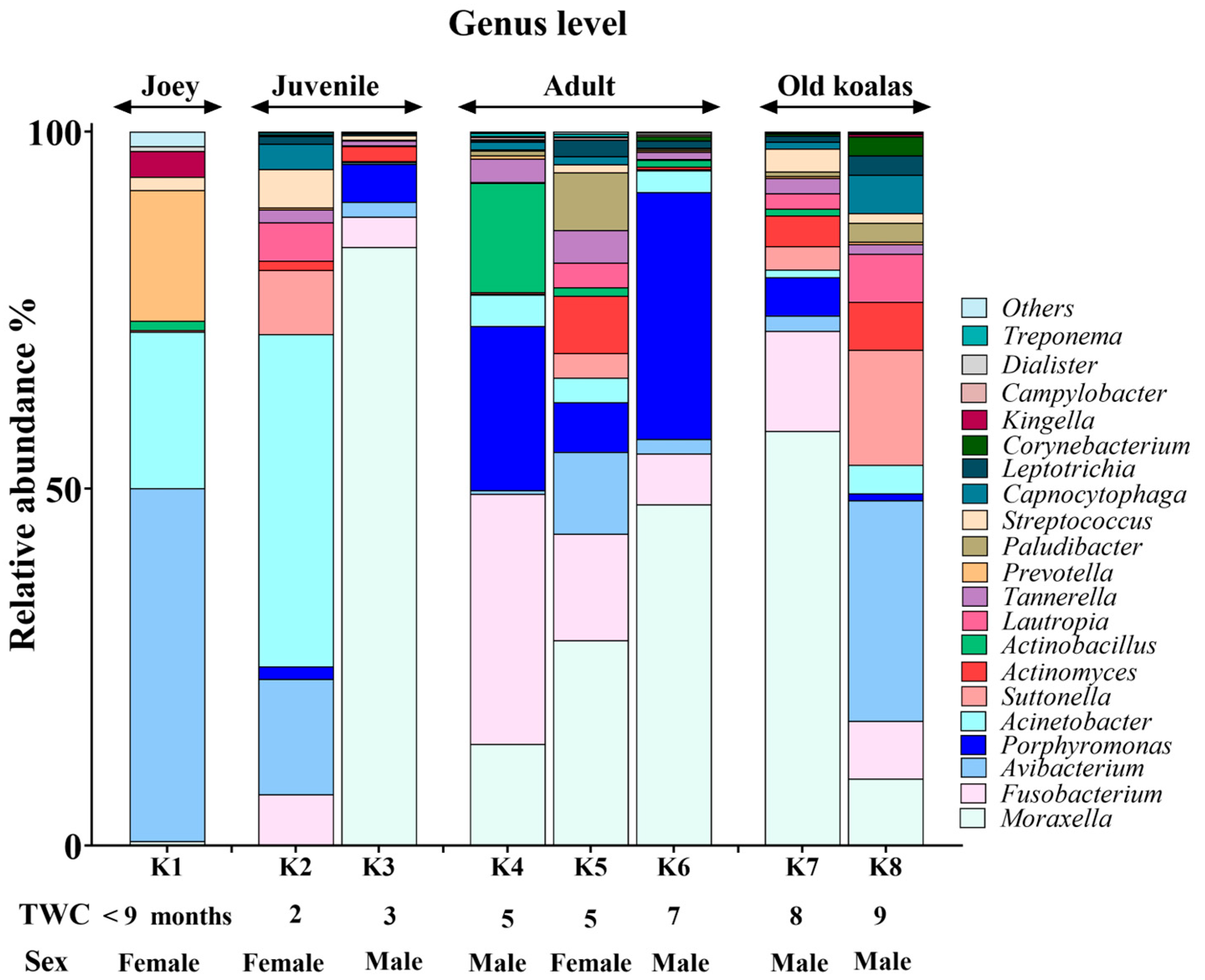
3.3. Genus-Level Oral Microbiome Composition
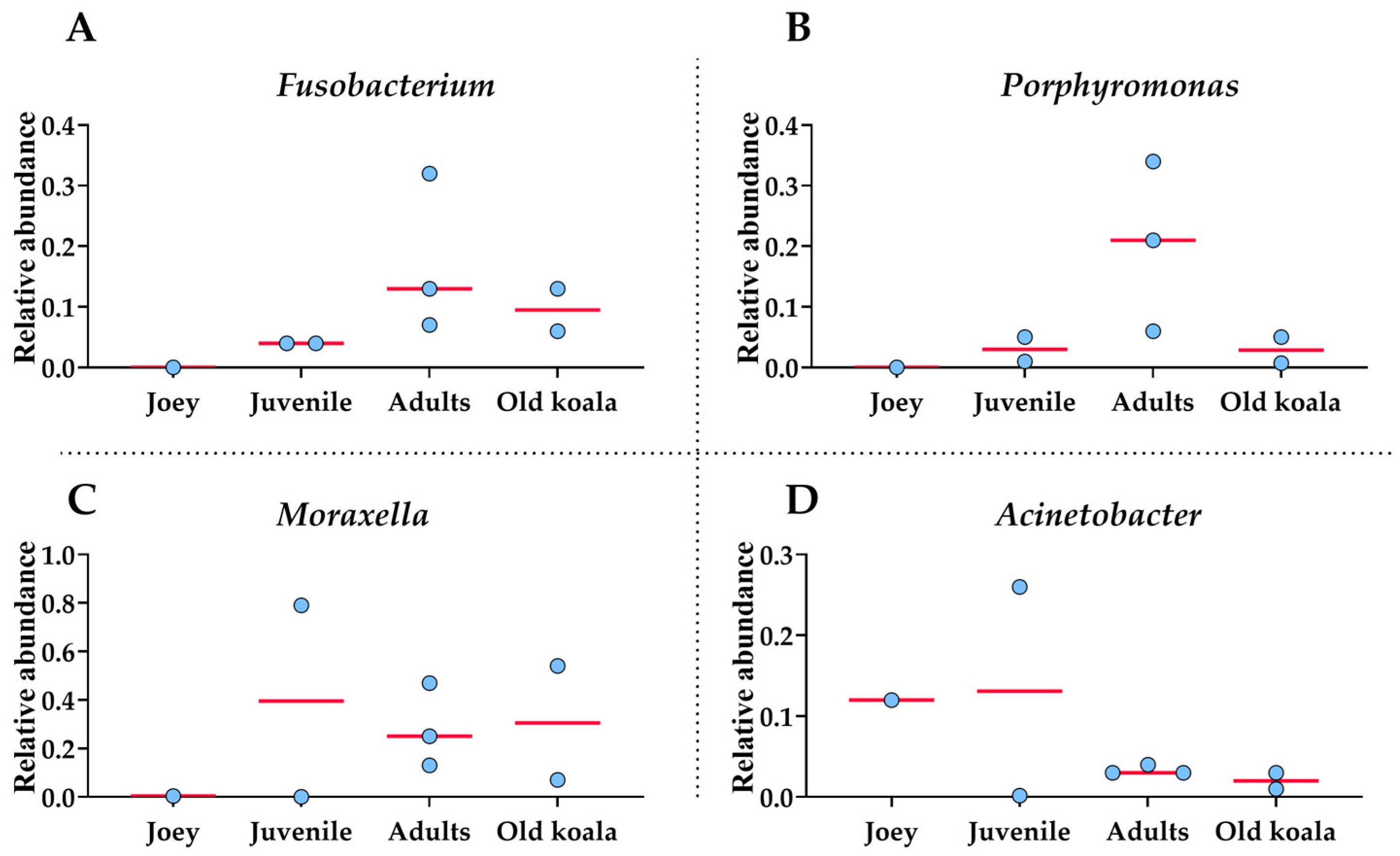
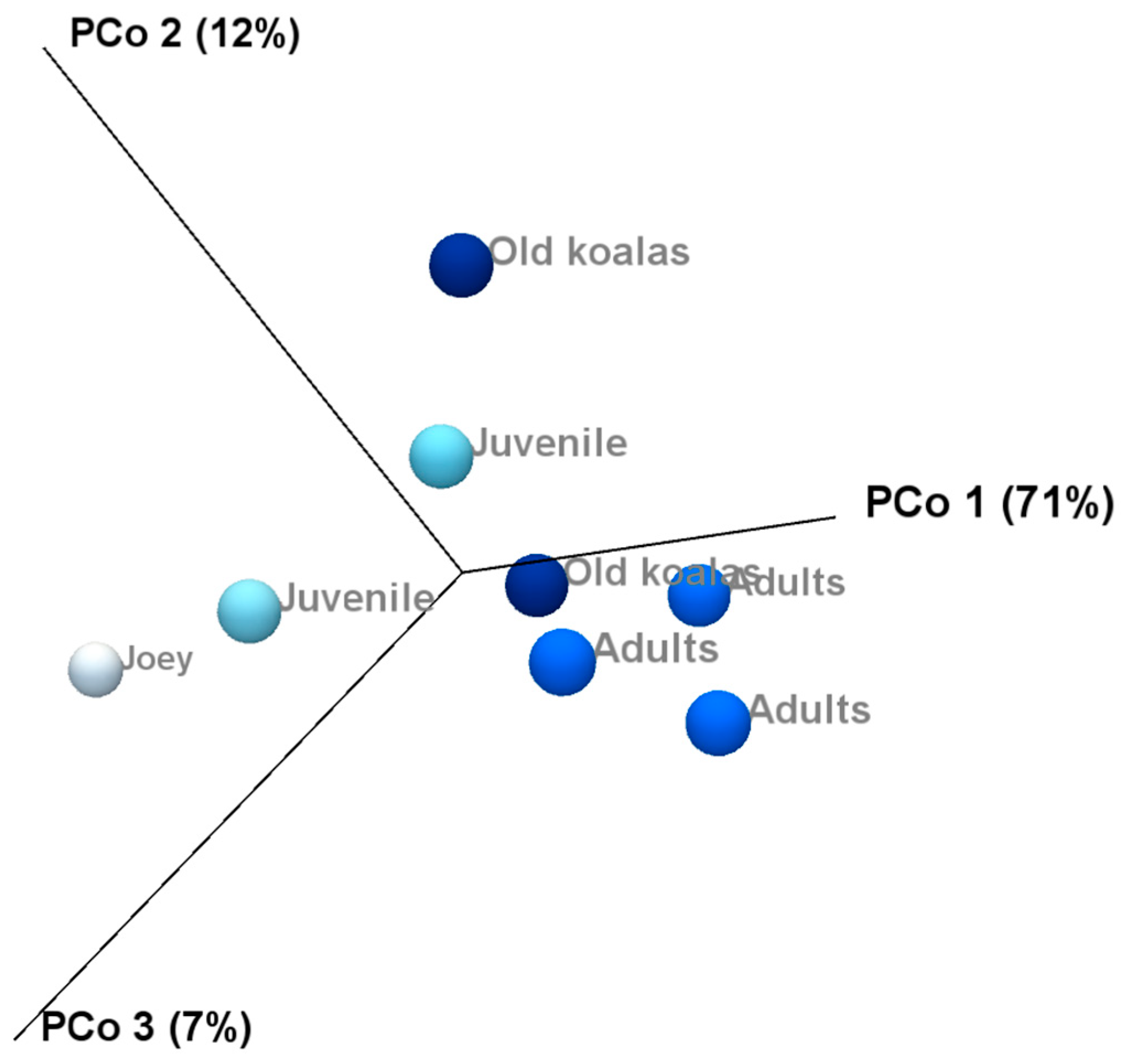
3.4. Age-Related Shifts in the Oral Microbiome
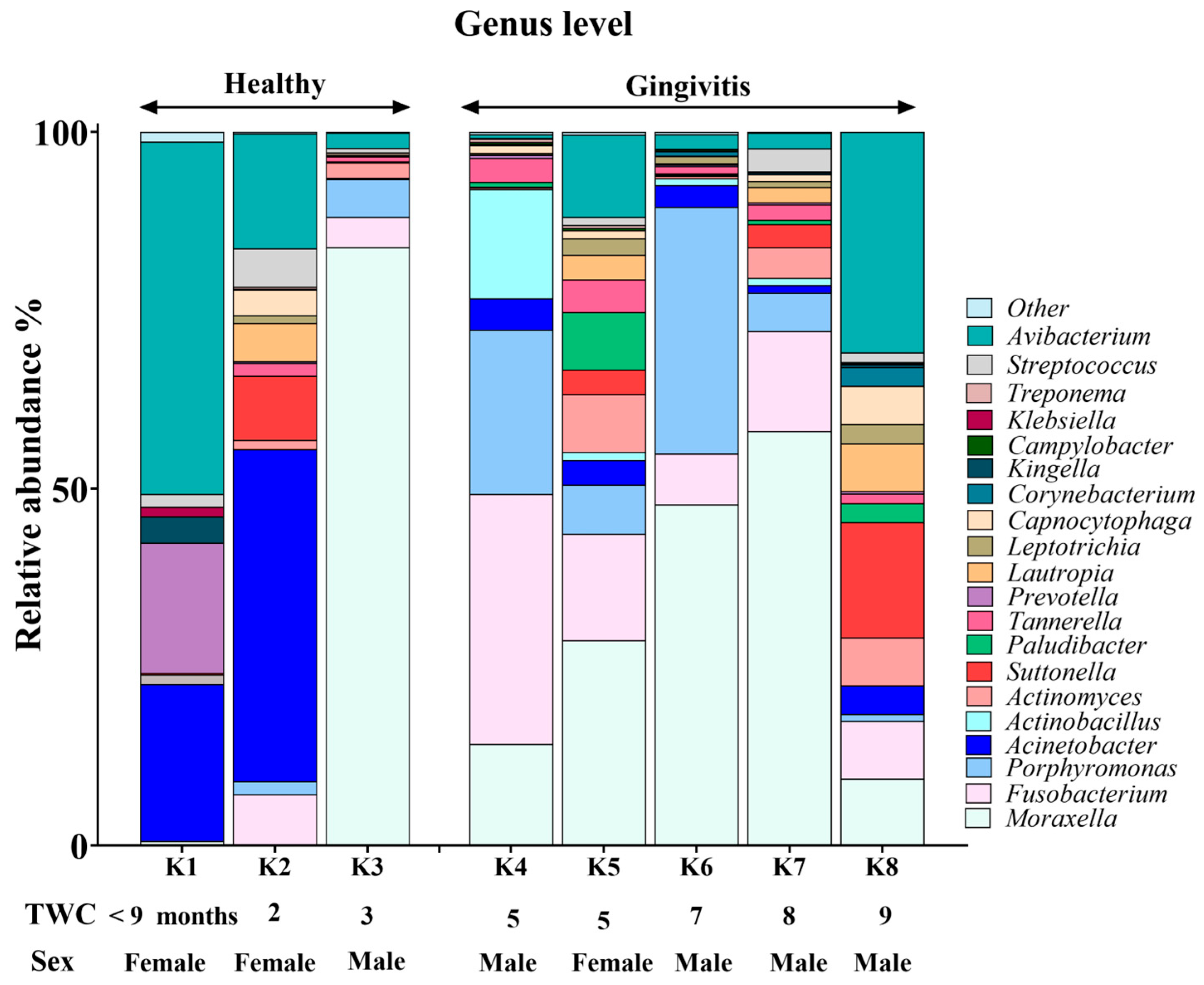
3.5. Oral Microbiome Composition in Koalas with Gingivitis and Periodontitis
3.6. Bacterial Diversity in Oral Koala Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quigley, B.L.; Timms, P. Helping koalas battle disease—Recent advances in Chlamydia and koala retrovirus (KoRV) disease understanding and treatment in koalas. FEMS Microbiol. Rev. 2020, 44, 583–605. [Google Scholar] [CrossRef]
- Zheng, H.; Pan, Y.; Tang, S.; Pye, G.W.; Stadler, C.K.; Vogelnest, L.; Herrin, K.V.; Rideout, B.A.; Switzer, W.M. Koala retrovirus diversity, transmissibility, and disease associations. Retrovirology 2020, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.F.; Varanasi, S.; Pettett, L.M.; Bird, P.S.; Symons, A.L. Loss of tooth-supporting bone in the koala (Phascolarctos cinereus) with age. Aust. J. Zool. 2011, 59, 49–53. [Google Scholar] [CrossRef]
- Pettett, L. Oral Health in South East Queensland Koalas: Prevalence of Periodontal Disease and Other Pathologies. Ph.D Thesis, University of Queensland, St Lucia, QLD, Australia, 2016. [Google Scholar]
- Pettett, L.M.; McKinnon, A.J.; Wilson, G.J.; Carrick, F.N.; Sly, L.I.; Bird, P.S. The development of an oral health charting system for koalas (Phascolarctos cinereus). J. Vet. Dent. 2012, 29, 232–241. [Google Scholar] [CrossRef]
- Butcher, R.; Pettett, L.; Fabijan, J.; Ebrahimie, E.; Mohammadi-Dehcheshmeh, M.; Speight, K.; Boardman, W.; Bird, P.; Trott, D. Periodontal disease in free-ranging koalas (Phascolarctos cinereus) from the Mount Lofty Ranges, South Australia, and its association with koala retrovirus infection. Aust. Vet. J. 2020, 98, 200–206. [Google Scholar] [CrossRef]
- Pettett, L.M.; Wilson, G.J.; Bird, P.S. An Oral Health Survey of Free-Ranging and Captive Koalas from Southeast Queensland, Australia. J. Vet. Dent. 2025, 8. [Google Scholar] [CrossRef]
- Bryce, A.J.; Milne, M.E.; Tyrrell, D.; Bodley, K. Case series: Computed tomography (CT) demonstrates lacrimal canal involvement in koalas (Phascolarctos cinereus) with maxillary incisor dental disease. Aust. Vet. J. 2022, 100, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef]
- Mikkelsen, D.; Milinovich, G.J.; Burrell, P.C.; Huynh, S.C.; Pettett, L.M.; Blackall, L.L.; Trott, D.J.; Bird, P.S. Phylogenetic analysis of Porphyromonas species isolated from the oral cavity of Australian marsupials. Environ. Microbiol. 2008, 10, 2425–2432. [Google Scholar] [CrossRef]
- Pettett, L.M.; Wilson, G.J.; Nicolson, V.; Boardman, W.; Speight, N.; Fabijan, J.; Trott, D.J.; Bird, P.S. Malocclusions in the koala (Phascolarctos cinereus). Aust. Vet. J. 2019, 97, 473–481. [Google Scholar] [CrossRef]
- Maki, K.A.; Kazmi, N.; Barb, J.J.; Ames, N. The Oral and Gut Bacterial Microbiomes: Similarities, Differences, and Connections. Biol. Res. Nurs. 2021, 23, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.T.; Taylor, G.W.; Scannapieco, F.; Kinane, D.F.; Curtis, M.; Beck, J.D.; Kogon, S. Periodontal health and systemic disorders. J. Can. Dent. Assoc. 2002, 68, 188–192. [Google Scholar]
- Caselli, E.; Fabbri, C.; D’Accolti, M.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the oral microbiome by whole-genome sequencing and resistome analysis: The complexity of the healthy picture. BMC Microbiol. 2020, 20, 120. [Google Scholar] [CrossRef]
- Gao, W.; Chan, Y.; You, M.; Lacap-Bugler, D.C.; Leung, W.K.; Watt, R.M. In-depth snapshot of the equine subgingival microbiome. Microb. Pathog. 2016, 94, 76–89. [Google Scholar] [CrossRef]
- Cheng, Y.; Siddle, H.V.; Belov, K. The Marsupial Major Histocompatibility Complex. In Marsupial Genetics and Genomics; Springer: Dordrecht, The Netherlands, 2010; pp. 339–356. [Google Scholar]
- Chhour, K.L.; Hinds, L.A.; Jacques, N.A.; Deane, E.M. An observational study of the microbiome of the maternal pouch and saliva of the tammar wallaby, Macropus eugenii, and of the gastrointestinal tract of the pouch young. Microbiol. (Read.) 2010, 156, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Alfano, N.; Courtiol, A.; Vielgrader, H.; Timms, P.; Roca, A.L.; Greenwood, A.D. Variation in koala microbiomes within and between individuals: Effect of body region and captivity status. Sci. Rep. 2015, 5, 10189. [Google Scholar] [CrossRef] [PubMed]
- Yip, S.; Dehcheshmeh, M.M.; McLelland, D.J.; Boardman, W.S.J.; Saputra, S.; Ebrahimie, E.; Weyrich, L.S.; Bird, P.S.; Trott, D.J. Porphyromonas spp., Fusobacterium spp., and Bacteroides spp. dominate microbiota in the course of macropod progressive periodontal disease. Sci. Rep. 2021, 11, 17775. [Google Scholar] [CrossRef] [PubMed]
- Bird, P.S. Oral disease in Animals: The Australian perspective. Isolation and Characterisation of Black-Pigmented bacteria from the Oral Cavity of Marsupials. Anaerobe 2002, 8, 79–87. [Google Scholar] [CrossRef]
- Bird, P.S.; Trott, D.J.; Mikkelsen, D.; Milinovich, G.J.; Hillman, K.M.; Burrell, P.C.; Blackall, L.L. Porphyromonas loveana sp. nov. a novel Porphyromonas species isolated from the oral cavity of Australian marsupials. Int. J. Syst. Evol. Microbiol. 2016, 66, 3771–3778. [Google Scholar] [CrossRef]
- Mandlik, J.S.; Patil, A.S.; Singh, S. Next-Generation Sequencing (NGS): Platforms and Applications. J. Pharm. Bioallied Sci. 2024, 16, S41–S45. [Google Scholar] [CrossRef]
- Weinstock, G.M. Genomic approaches to studying the human microbiota. Nature 2012, 489, 250–256. [Google Scholar] [CrossRef]
- Avila, M.; Ojcius, D.M.; Yilmaz, O. The oral microbiota: Living with a permanent guest. DNA Cell Biol. 2009, 28, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S. Australian Mammals: Biology and Captive Management; CSIRO Publishing: Collingwood, VIC, Australia, 2003; p. 524. [Google Scholar]
- Gordon, G. Estimation of the age of the koala Phascolarctos cinereus (Marsupialia: Phascolarctidae) from tooth wear and growth. Aust. Mammal. 1991, 14, 5–12. [Google Scholar] [CrossRef]
- AVDC. Veterinary Dental Nomenclature. Available online: https://avdc.org/avdc-nomenclature/ (accessed on 14 January 2011).
- Caiafa, A. Oral Examination/Dental Charting and Diagnostic Tools. In Proceedings of the WSAVA, Auckland, New Zealand, 6–8 March 2013. [Google Scholar]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef]
- McDonald, D.; Jiang, Y.; Balaban, M.; Cantrell, K.; Zhu, Q.; Gonzalez, A.; Morton, J.T.; Nicolaou, G.; Parks, D.H.; Karst, S.M. Greengenes2 unifies microbial data in a single reference tree. Nat. Biotechnol. 2024, 42, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Chen, J.; Lv, W.; Zhang, X.; Zhang, T.; Dong, J.; Wang, Z.; Liu, T.; Zhang, P.; Pyne, M.; Dong, G. Animal age affects the gut microbiota and immune system in captive koalas (Phascolarctos cinereus). Microbiol. Spectr. 2023, 11, e04101-22. [Google Scholar] [CrossRef]
- Sinclair, H.A.; Chapman, P.; Omaleki, L.; Bergh, H.; Turni, C.; Blackall, P.; Papacostas, L.; Braslins, P.; Sowden, D.; Nimmo, G.R. Identification of Lonepinella sp. in Koala Bite Wound Infections, Queensland, Australia. Emerg. Infect. Dis. 2019, 25, 153–156. [Google Scholar] [CrossRef]
- Sarkar, P.; Malik, S.; Laha, S.; Das, S.; Bunk, S.; Ray, J.G.; Chatterjee, R.; Saha, A. Dysbiosis of oral microbiota during oral squamous cell carcinoma development. Front. Oncol. 2021, 11, 614448. [Google Scholar] [CrossRef]
- Xi, M.; Ruan, Q.; Zhong, S.; Li, J.; Qi, W.; Xie, C.; Wang, X.; Abuduxiku, N.; Ni, J. Periodontal bacteria influence systemic diseases through the gut microbiota. Front. Cell. Infect. Microbiol. 2024, 14, 1478362. [Google Scholar] [CrossRef]
- Peng, X.; Cheng, L.; You, Y.; Tang, C.; Ren, B.; Li, Y.; Xu, X.; Zhou, X. Oral microbiota in human systematic diseases. Int. J. Oral Sci. 2022, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Groeger, S.; Zhou, Y.; Ruf, S.; Meyle, J. Pathogenic mechanisms of Fusobacterium nucleatum on oral epithelial cells. Front. Oral Health 2022, 3, 831607. [Google Scholar] [CrossRef]
- De Andrade, K.Q.; Almeida-da-Silva, C.L.C.; Coutinho-Silva, R. Immunological pathways triggered by Porphyromonas gingivalis and Fusobacterium nucleatum: Therapeutic possibilities? Mediat. Inflamm. 2019, 2019, 7241312. [Google Scholar] [CrossRef] [PubMed]
- Aral, K.; Milward, M.R.; Cooper, P.R. Dysregulation of inflammasomes in human dental pulp cells exposed to Porphyromonas gingivalis and Fusobacterium nucleatum. J. Endod. 2020, 46, 1265–1272. [Google Scholar] [CrossRef]
- Menghani, S.V. Carcinogenetic mechanisms employed by the oral microbiome: A narrative review. Am. J. Med. Sci. 2025, 369, 56–561. [Google Scholar] [CrossRef]
- Wu, Y.; Dong, G.; Xiao, W.; Xiao, E.; Miao, F.; Syverson, A.; Missaghian, N.; Vafa, R.; Cabrera-Ortega, A.A.; Rossa, C., Jr.; et al. Effect of Aging on Periodontal Inflammation, Microbial Colonization, and Disease Susceptibility. J. Dent. Res. 2016, 95, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Krockenberger, A.K.; Hume, I.D.; Cork, S.J. Production of Milk and Nutrition of the Dependent Young of Free-Ranging Koalas (Phascolarctos cinereus). Physiol. Zool. 1998, 71, 45–56. [Google Scholar] [CrossRef]
- Blyton, M.D.J.; Soo, R.M.; Hugenholtz, P.; Moore, B.D. Maternal inheritance of the koala gut microbiome and its compositional and functional maturation during juvenile development. Environ. Microbiol. 2022, 24, 475–493. [Google Scholar] [CrossRef]
- Liu, Z.; Cao, Q.; Wang, W.; Wang, B.; Yang, Y.; Xian, C.J.; Li, T.; Zhai, Y. The Impact of Lactobacillus reuteri on Oral and Systemic Health: A Comprehensive Review of Recent Research. Microorganisms 2024, 13, 45. [Google Scholar] [CrossRef]
- Alvarenga, D.O.; Franco, M.W.; Sivonen, K.; Fiore, M.F.; Varani, A.M. Evaluating Eucalyptus leaf colonization by Brasilonema octagenarum (Cyanobacteria, Scytonemataceae) using in planta experiments and genomics. PeerJ 2020, 8, e9158. [Google Scholar] [CrossRef]
- Littleford-Colquhoun, B.L.; Weyrich, L.S.; Hohwieler, K.; Cristescu, R.; Frère, C.H. How microbiomes can help inform conservation: Landscape characterisation of gut microbiota helps shed light on additional population structure in a specialist folivore. Anim. Microbiome 2022, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Lott, M.J.; Frankham, G.J.; Eldridge, M.D.B.; Alquezar-Planas, D.E.; Donnelly, L.; Zenger, K.R.; Leigh, K.A.; Kjeldsen, S.R.; Field, M.A.; Lemon, J.; et al. Reversing the decline of threatened koala (Phascolarctos cinereus) populations in New South Wales: Using genomics to enhance conservation outcomes. Ecol. Evol. 2024, 14, e11700. [Google Scholar] [CrossRef]
- Grant, L.; Mohammadi Dehcheshmeh, M.; Ebrahimie, E.; Khabiri, A.; Veltman, T.; Shipstone, M.; Trott, D.J. The effect of daily oral probiotic and postbiotic supplementation on the canine skin microbiota: Insights from culture-dependent and long-read 16S rRNA gene sequencing methods. Vet. Dermatol 2025. Early View. [Google Scholar] [CrossRef]
- Kondo, K.; Suzuki, M.; Amadaira, M.; Araki, C.; Watanabe, R.; Murakami, K.; Ochiai, S.; Ogura, T.; Hayakawa, T. Association of maternal genetics with the gut microbiome and eucalypt diet selection in captive koalas. PeerJ 2024, 12, e17385. [Google Scholar] [CrossRef]
- Whisson, D.A.; Ashman, K.R. When an iconic native animal is overabundant: The koala in southern Australia. Conserv. Sci. Pract. 2020, 2, e188. [Google Scholar] [CrossRef]
- Lanyon, J.; Sanson, G. Koala (Phascolarctos cinereus) dentition and nutrition. II. Implications of tooth wear in nutrition. J. Zool. 2011, 209, 169–181. [Google Scholar] [CrossRef]
- Martínez-García, M.; Hernández-Lemus, E. Periodontal Inflammation and Systemic Diseases: An Overview. Front. Physiol. 2021, 12, 709438. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.; Meers, J.; Young, P. Biology and evolution of the endogenous koala retrovirus. Cell. Mol. Life Sci. 2008, 65, 3413–3421. [Google Scholar] [CrossRef]
- Lee, Y.H.; Chung, S.W.; Auh, Q.S.; Hong, S.J.; Lee, Y.A.; Jung, J.; Lee, G.J.; Park, H.J.; Shin, S.I.; Hong, J.Y. Progress in Oral Microbiome Related to Oral and Systemic Diseases: An Update. Diagnostics 2021, 11, 1283. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Koala | Sex | TWC | Conjunctivitis Severity | Cystitis Severity | Outcome | |
|---|---|---|---|---|---|---|
| Left Eye | Right Eye | |||||
| 1 | F | 9 months | 0 | 0 | 0 | no treatment/released |
| 2 | F | 2 | 3 | 3 | 0 | treated/released |
| 3 | M | 3 | 1, injured eye | 0 | 0 | treated/released |
| 4 | M | 5 | 3 | 3 | 0 | treated/released |
| 5 | F | 5 | 3 | 3 | 0 | treated/released |
| 6 | M | 7 | 3 | 3 | 0 | treated/released |
| 7 | M | 8 | blind/damaged eye | 0 | 0 | treated/released |
| 8 | M | 9 | 1 | 1 | 0 | treated/released |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pettett, L.; Ebrahimie, E.; Chinkangsadarn, T.; Mohammadi Dehcheshmeh, M.; Trott, D.J.; Bird, P.S. The Oral Microbiome in Queensland Free-Ranging Koalas (Phascolarctos cinereus) and Its Association with Age and Periodontal Disease. Animals 2025, 15, 1834. https://doi.org/10.3390/ani15131834
Pettett L, Ebrahimie E, Chinkangsadarn T, Mohammadi Dehcheshmeh M, Trott DJ, Bird PS. The Oral Microbiome in Queensland Free-Ranging Koalas (Phascolarctos cinereus) and Its Association with Age and Periodontal Disease. Animals. 2025; 15(13):1834. https://doi.org/10.3390/ani15131834
Chicago/Turabian StylePettett, Lyndall, Esmaeil Ebrahimie, Teerapol Chinkangsadarn, Manijeh Mohammadi Dehcheshmeh, Darren J. Trott, and Philip S. Bird. 2025. "The Oral Microbiome in Queensland Free-Ranging Koalas (Phascolarctos cinereus) and Its Association with Age and Periodontal Disease" Animals 15, no. 13: 1834. https://doi.org/10.3390/ani15131834
APA StylePettett, L., Ebrahimie, E., Chinkangsadarn, T., Mohammadi Dehcheshmeh, M., Trott, D. J., & Bird, P. S. (2025). The Oral Microbiome in Queensland Free-Ranging Koalas (Phascolarctos cinereus) and Its Association with Age and Periodontal Disease. Animals, 15(13), 1834. https://doi.org/10.3390/ani15131834