
Identification of Thermoneutral Zone in Sahiwal Zebu Calves in Subtropical Climate of India
 ,
,  , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Place of Study
2.2. Animals
2.3. Experimental Design
2.4. Psychrometric Chamber
2.5. Meteorological Variables
2.6. Physiological Observations
2.7. Blood Sampling
2.8. Hematological Analysis
2.9. Biochemical Analysis
2.10. Statistical Analysis
2.11. Mathematical Output
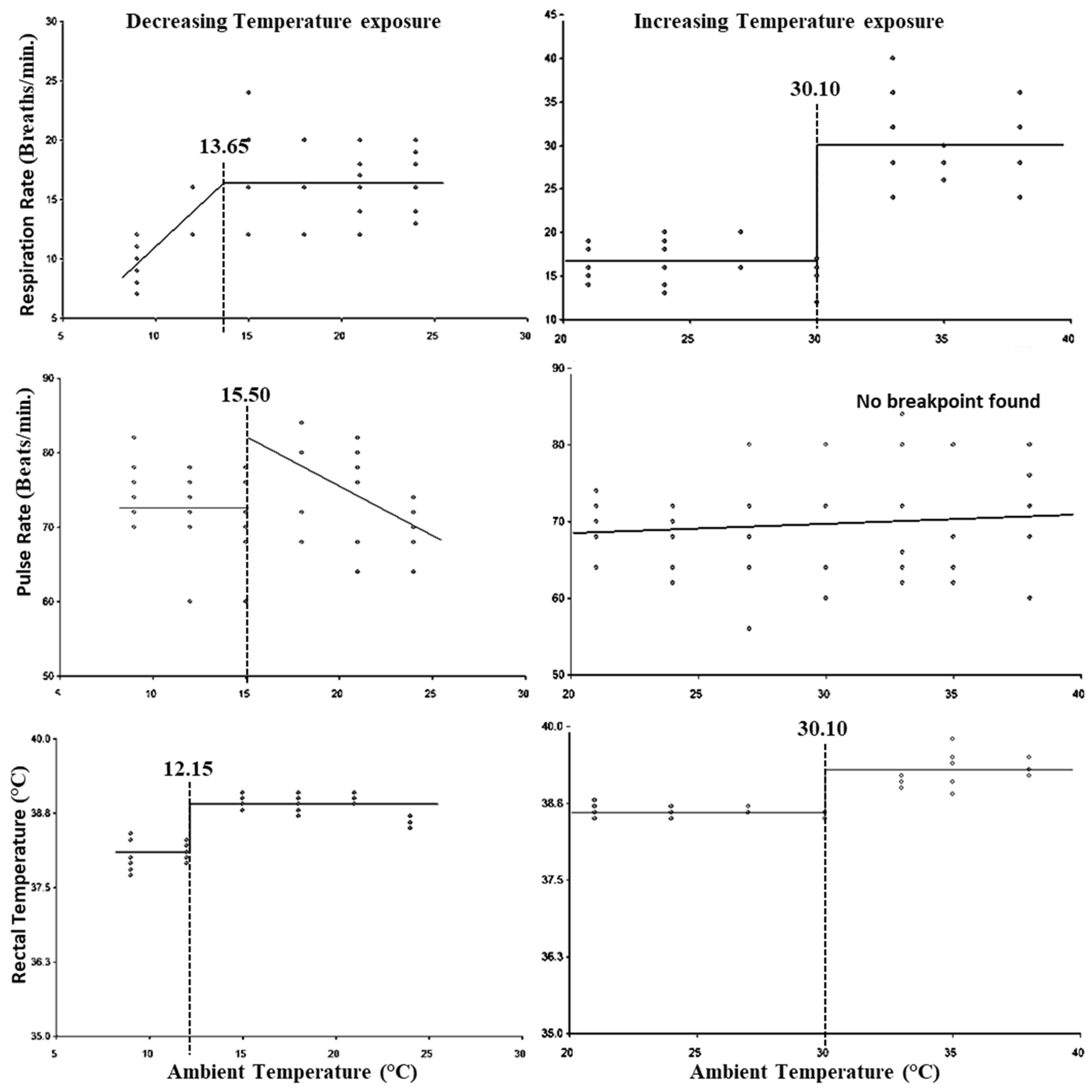
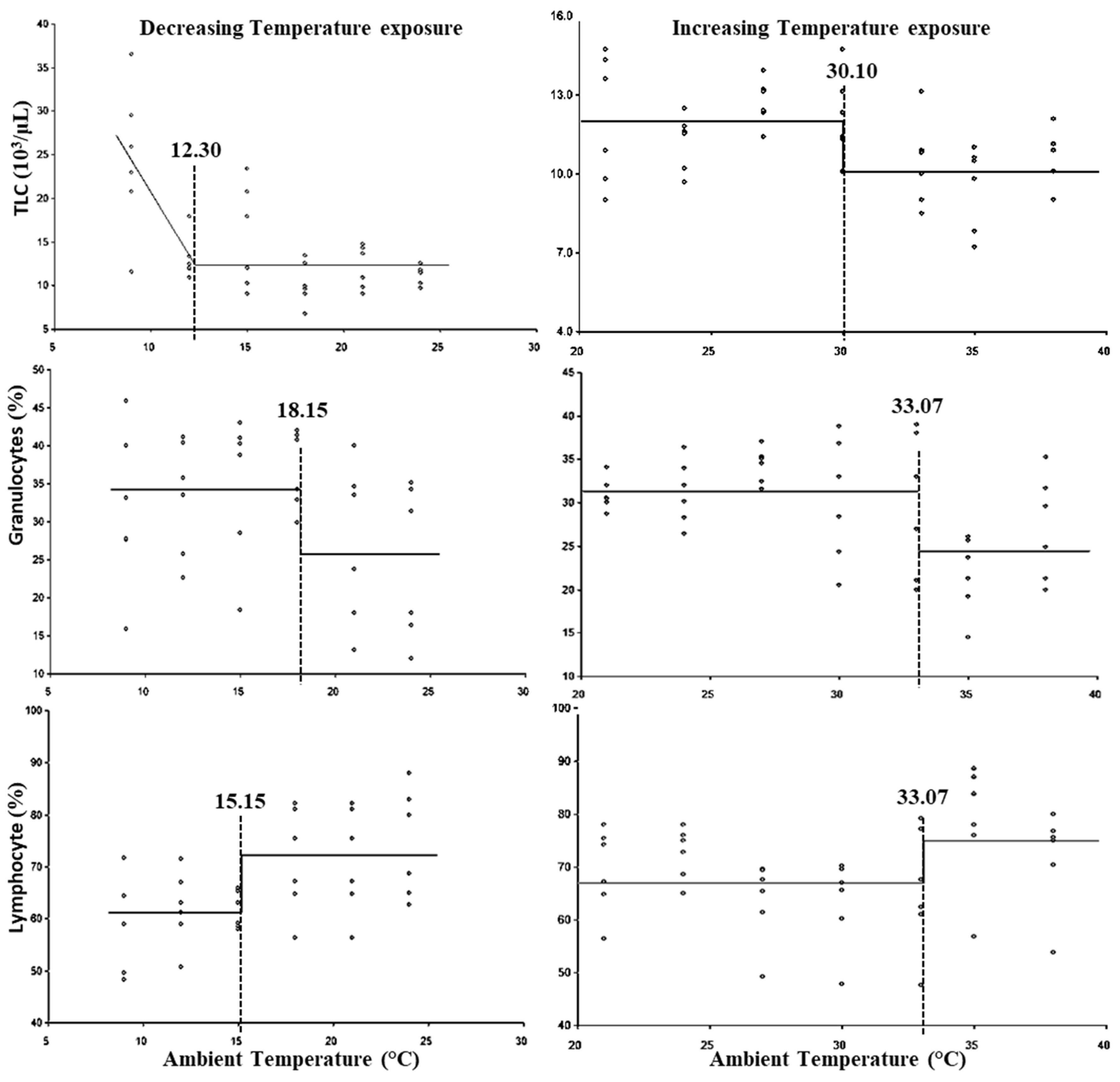
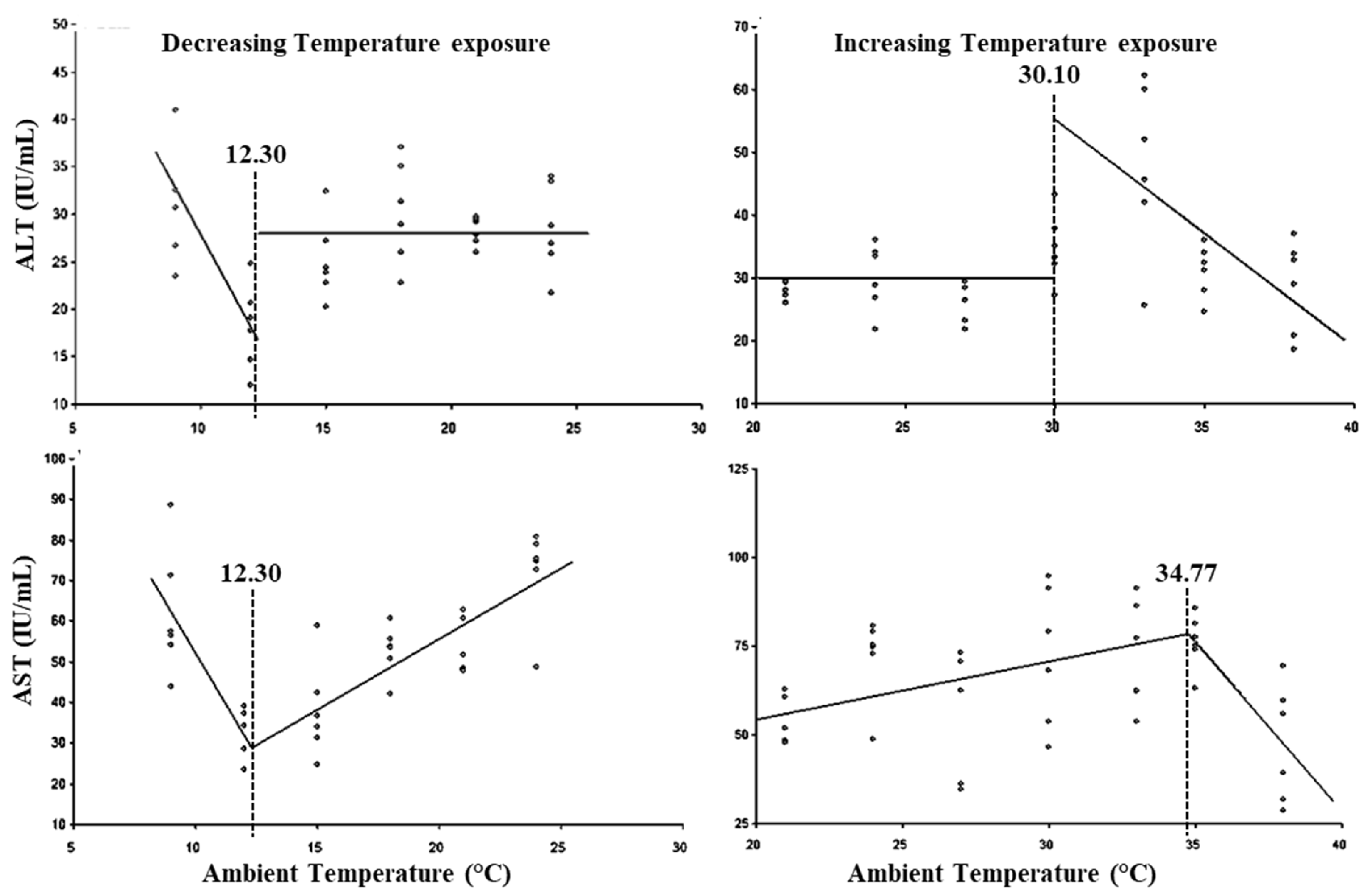
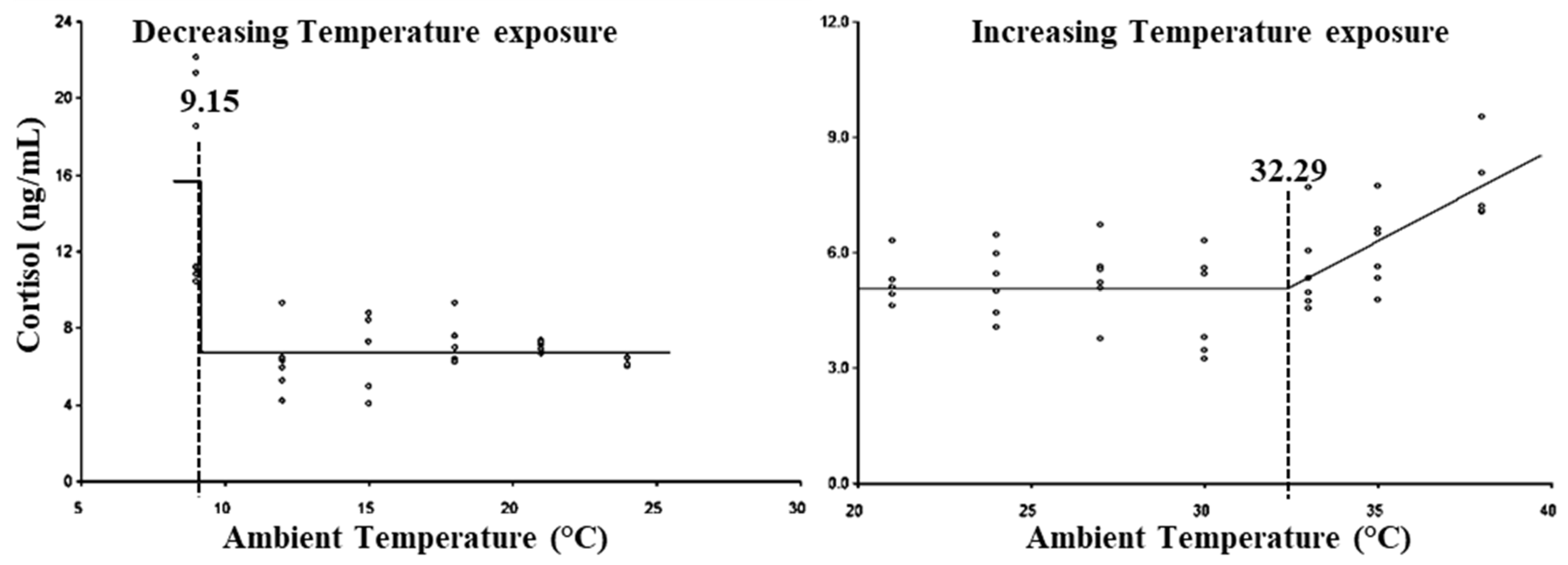
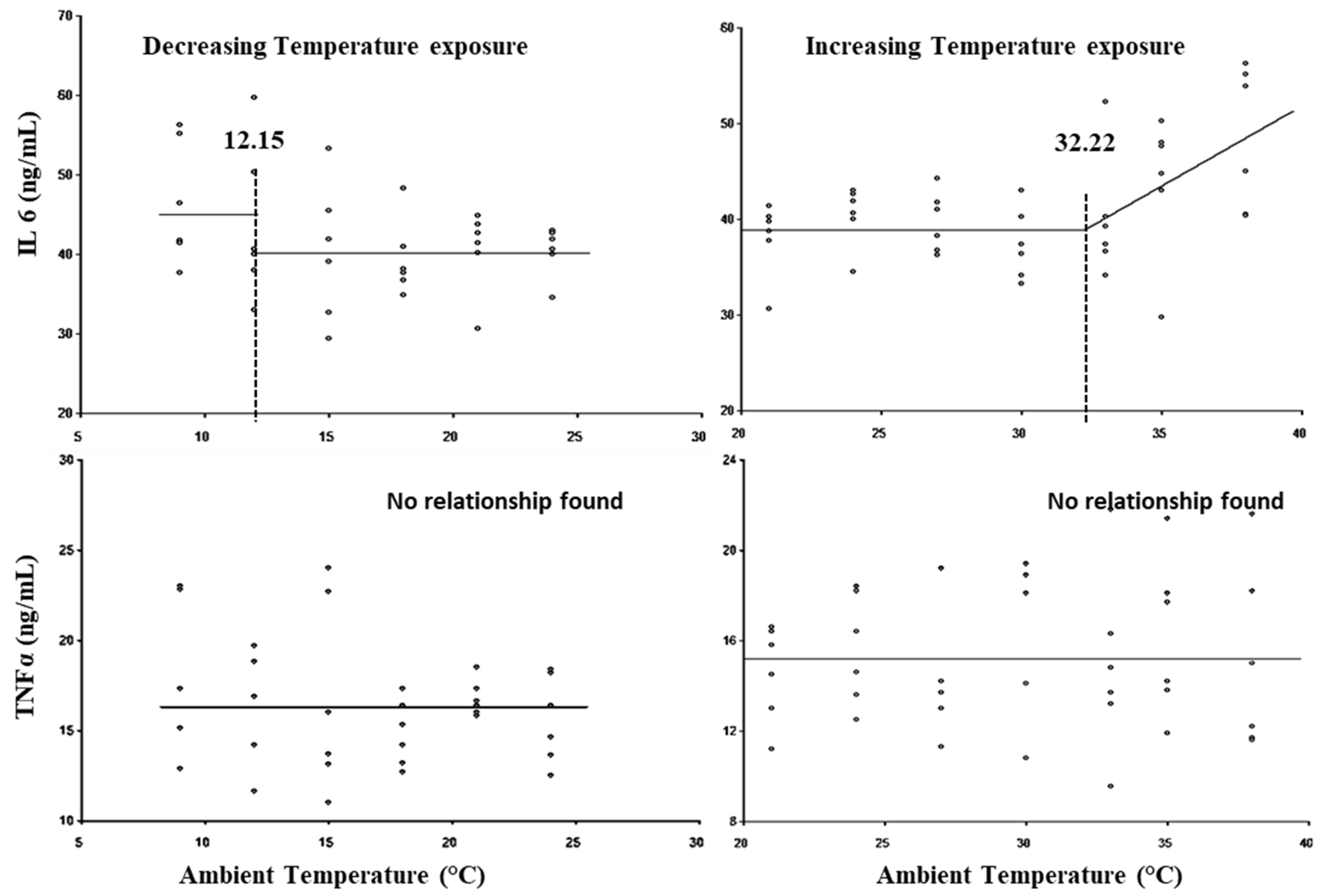
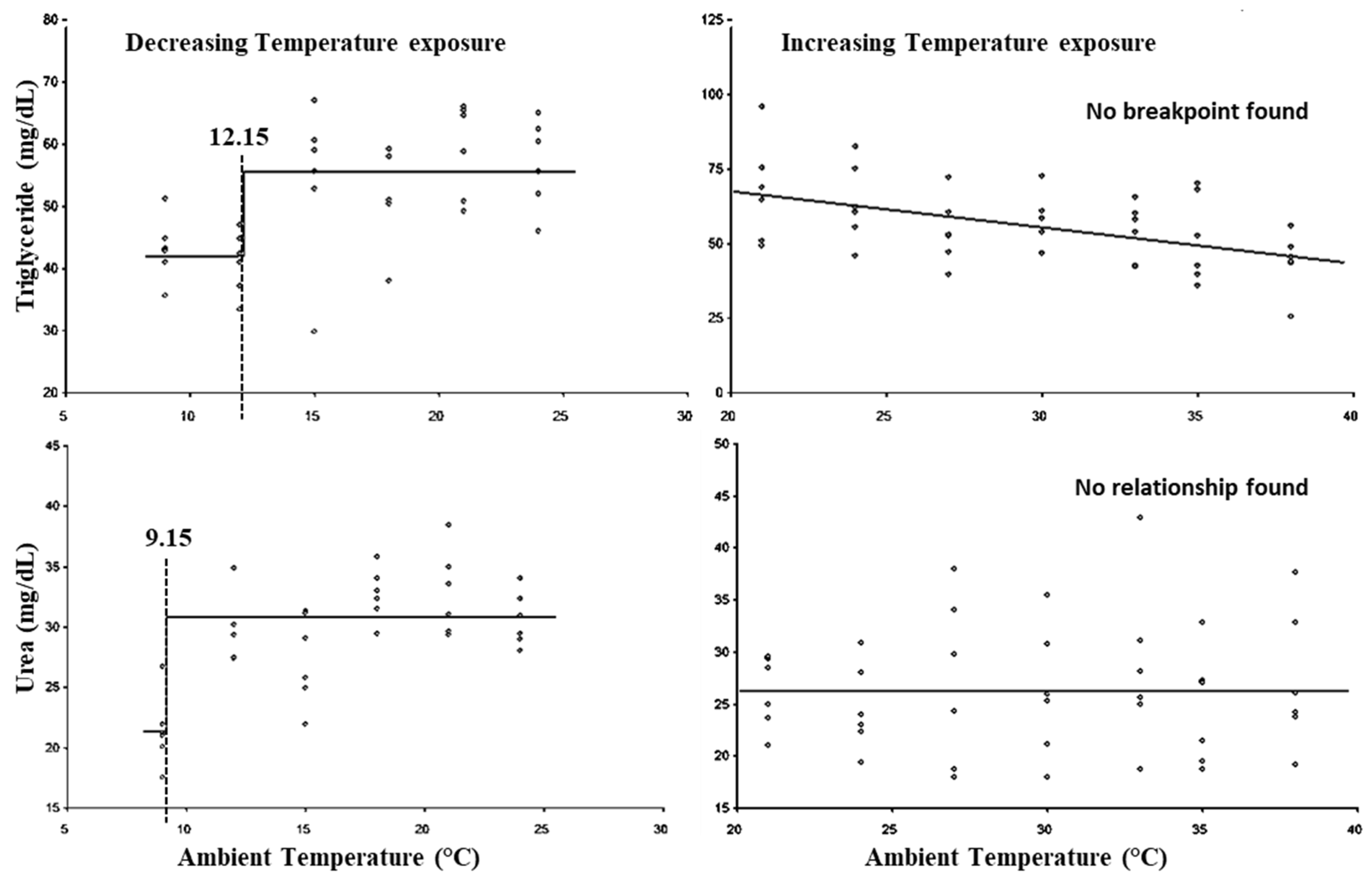
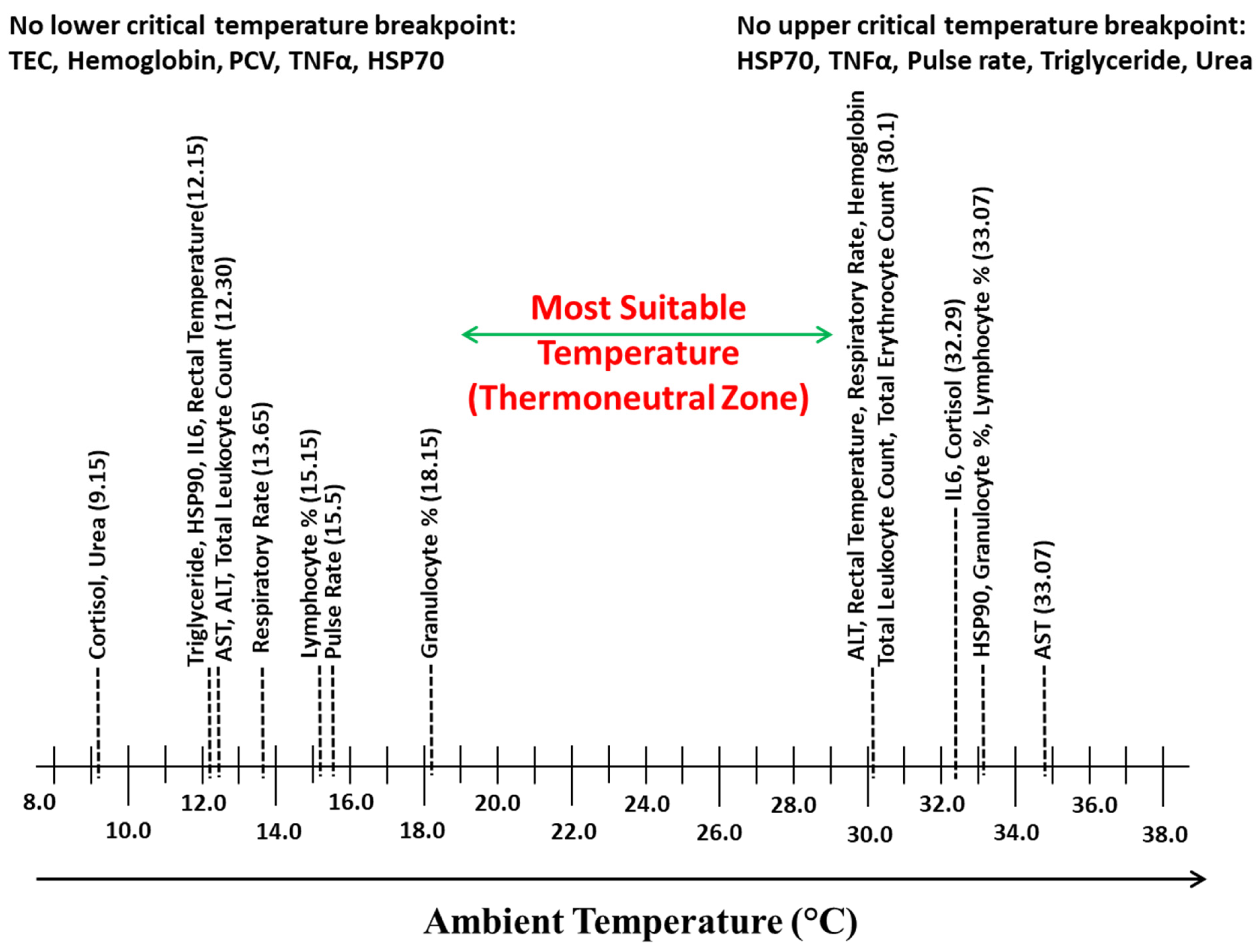
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- West, J.W. Effects of Heat-Stress on Production in Dairy Cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.; Singh, G.; Wankar, A.; Dutta, N.; Chaturvedi, V.B.; Verma, M.R. Effect of Simulated Heat Stress on Digestibility, Methane Emission and Metabolic Adaptability in Crossbred Cattle. Asian-Australas. J. Anim. Sci. 2016, 29, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Yadav, B.; Madan, A.K.; Kumar, M.; Sirohi, R.; Reddy, A.V. Dynamics of Heat-Shock Proteins, Metabolic and Endocrine Responses during Increasing Temperature Humidity Index (THI) in Lactating Hariana (Zebu) Cattle. Biol. Rhythm Res. 2019, 51, 934–950. [Google Scholar] [CrossRef]
- Butt, J.H.; Konwar, D.; Brahma, B.; Khan, A.; Chakraborty, D. Assessment of Cold Stress in Crossbred Cattle by Wind Chill Temperature Index. Indian J. Anim. Sci. 2022, 91, 1109–1111. [Google Scholar] [CrossRef]
- Brody, S. Climatic Physiology of Cattle. J. Dairy Sci. 1956, 39, 715–725. [Google Scholar] [CrossRef]
- Bianca, W. Review of the progress of dairy science: Cattle in hot environment. J. Dairy Sci. 1965, 32, 291–308. [Google Scholar]
- Berman, A.; Meltzer, A. Critical Temperatures in Lactating Dairy Cattle: A New Approach to an Old Problem. Int. J. Biometeorol. 1973, 17, 167–176. [Google Scholar] [CrossRef]
- El-Tarabany, M.S.; El-Tarabany, A.A.; Atta, M.A. Physiological and Lactation Responses of Egyptian Dairy Baladi Goats to Natural Thermal Stress under Subtropical Environmental Conditions. Int. J. Biometeorol. 2016, 61, 61–68. [Google Scholar] [CrossRef]
- Jeelani, R.; Konwar, D.; Khan, A.; Kumar, D.; Chakraborty, D.; Brahma, B. Reassessment of Temperature-Humidity Index for Measuring Heat Stress in Crossbred Dairy Cattle of a Sub-Tropical Region. J. Therm. Biol. 2019, 82, 99–106. [Google Scholar] [CrossRef]
- Domínguez, R.R.L.; Peláez, C.G.V.; Padilla, E.G. Effect of heat stress and its interaction with other management and productive variables on pregnancy rate in dairy cows in Aguascalientes, Mexico. Vet. Mex. 2005, 36, 245–260. [Google Scholar]
- Dash, S.; Chakravarty, A.K.; Singh, A.; Behera, R.; Upadhyay, A.; Shivahre, P.R. Determination of Critical Heat Stress Zone for Fertility Traits Using Temperature Humidity Index in Murrah Buffaloes. Indian J. Anim. Sci. 2014, 84, 1181–1184. [Google Scholar] [CrossRef]
- Dash, S.; Chakravarty, A.K.; Sah, V.; Jamuna, V.; Behera, R.; Kashyap, N.; Deshmukh, B. Influence of Temperature and Humidity on Pregnancy Rate of Murrah Buffaloes under Subtropical Climate. Asian-Australas. J. Anim. Sci. 2015, 28, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Hillman, P.; Gebremedhin, K.G.; Willard, S.T.; Lee, C.-S.; Kennedy, A.D. Continuous Measurements of Vaginal Temperature of Female Cattle Using a Data Logger Encased in a Plastic Anchor. Appl. Eng. Agric. 2009, 25, 291–296. [Google Scholar] [CrossRef]
- Atkins, I.K.; Cook, N.B.; Mondaca, M.R.; Choi, C.Y. Continuous Respiration Rate Measurement of Heat-Stressed Dairy Cows and Relation to Environment, Body Temperature, and Lying Time. Trans. ASABE 2018, 61, 1475–1485. [Google Scholar] [CrossRef]
- Ammer, S.; Lambertz, C.; Gauly, M. Is Reticular Temperature a Useful Indicator of Heat Stress in Dairy Cattle? J. Dairy Sci. 2016, 99, 10067–10076. [Google Scholar] [CrossRef]
- Ji, B.; Banhazi, T.; Ghahramani, A.; Bowtell, L.; Wang, C.; Li, B. Modelling of Heat Stress in a Robotic Dairy Farm. Part 1: Thermal Comfort Indices as the Indicators of Production Loss. Biosyst. Eng. 2019, 199, 27–42. [Google Scholar] [CrossRef]
- Kim, N.Y.; Kim, S.J.; Oh, M.; Jang, S.Y.; Moon, S.H. Changes in Facial Surface Temperature of Laying Hens under Different Thermal Conditions. Asian-Australas. J. Anim. Sci. 2020, 34, 1235–1242. [Google Scholar] [CrossRef]
- Peng, D.; Chen, S.; Li, G.; Chen, J.; Wang, J.; Gu, X. Infrared Thermography Measured Body Surface Temperature and Its Relationship with Rectal Temperature in Dairy Cows under Different Temperature-Humidity Indexes. Int. J. Biometeorol. 2019, 63, 327–336. [Google Scholar] [CrossRef]
- Kovács, L.; Kézér, F.L.; Póti, P.; Boros, N.; Nagy, K. Short Communication: Upper Critical Temperature-Humidity Index for Dairy Calves Based on Physiological Stress Variables. J. Dairy Sci. 2020, 103, 2707–2710. [Google Scholar] [CrossRef]
- Yan, G.; Shi, Z.; Li, H. Critical Temperature-Humidity Index Thresholds Based on Surface Temperature for Lactating Dairy Cows in a Temperate Climate. Agriculture 2021, 11, 970. [Google Scholar] [CrossRef]
- Yadav, B.; Yadav, S.; Madan, A.K.; Anand, M.; Swain, D.K.; Pandey, V.; Sirohi, R. Heat stress responses to increasing temperature humidity index (THI) in lactating Murrah buffalo. Buffalo Bull. 2022, 41, 161–170. [Google Scholar] [CrossRef]
- Srivastava, A.; Yadav, P.; Mahajan, A.; Anand, M.; Yadav, S.; Madan, A.K.; Yadav, B. Appropriate THI Model and Its Threshold for Goats in Semi-Arid Regions of India. J. Therm. Biol. 2021, 96, 102845. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Yadav, B.; Swain, D.K.; Anand, M.; Yadav, S.; Madan, A.K. Differential Expression of MiRNAs and Related MRNAs during Heat Stress in Buffalo Heifers. J. Therm. Biol. 2021, 97, 102904. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.; Madan, A.K.; Yadav, S.; Pandey, V.; Sirohi, R. Effect of Cooling Strategies on Milk Production, Physiological Variables and Blood Profile during Hot-Dry and Hot-Humid Summer in Sahiwal Cattle. Indian J. Anim. Sci. 2021, 91, 845–850. [Google Scholar] [CrossRef]
- Yadav, B.; Singh, G.; Wankar, A. Acclimatization Dynamics to Extreme Heat Stress in Crossbred Cattle. Biol. Rhythm Res. 2019, 52, 524–534. [Google Scholar] [CrossRef]
- Kumar, J.; Madan, A.K.; Kumar, M.; Sirohi, R.; Yadav, B.; Reddy, A.V.; Swain, D.K. Impact of Season on Antioxidants, Nutritional Metabolic Status, Cortisol and Heat Shock Proteins in Hariana and Sahiwal Cattle. Biol. Rhythm Res. 2017, 49, 29–38. [Google Scholar] [CrossRef]
- Zhang, F.J.; Weng, X.G.; Wang, J.F.; Zhou, D.; Zhang, W.; Zhai, C.C.; Hou, Y.X.; Zhu, Y.H. Effects of Temperature–Humidity Index and Chromium Supplementation on Antioxidant Capacity, Heat Shock Protein 72, and Cytokine Responses of Lactating Cows1. J. Anim. Sci. 2014, 92, 3026–3034. [Google Scholar] [CrossRef]
- Collier, R.J.; Beede, D.K.; Thatcher, W.W.; Israel, L.A.; Wilcox, C.J. Influences of Environment and Its Modification on Dairy Animal Health and Production. J. Dairy Sci. 1982, 65, 2213–2227. [Google Scholar] [CrossRef]
- Roland, L.; Drillich, M.; Klein-Jöbstl, D.; Iwersen, M. Invited review: Influence of climatic conditions on the development, performance, and health of calves. J. Dairy Sci. 2016, 99, 2438–2452. [Google Scholar] [CrossRef]
- Scibilia, L.S.; Muller, L.D.; Kensinger, R.S.; Sweeney, T.F.; Shellenberger, P.R. Effect of Environmental Temperature and Dietary Fat on Growth and Physiological Responses of Newborn Calves. J. Dairy Sci. 1987, 70, 1426–1433. [Google Scholar] [CrossRef]
- Kang, H.J.; Lee, I.K.; Piao, M.Y.; Gu, M.J.; Yun, C.H.; Kim, H.J.; Kim, K.H.; Baik, M. Effects of Ambient Temperature on Growth Performance, Blood Metabolites, and Immune Cell Populations in Korean Cattle Steers. Asian-Australas. J. Anim. Sci. 2016, 29, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.P.; Papasian, C.J.; Ritter, R.C. The effects of cold stress on neonatal calves. II. Absorption of colostral immunoglobulins. Can. J. Comp. Med. 1980, 44, 19. [Google Scholar] [PubMed]
- Nakajima, N.; Doi, K.; Tamiya, S.; Yayota, M. Effects of Direct Exposure to Cold Weather under Grazing in Winter on the Physiological, Immunological, and Behavioral Conditions of Japanese Black Beef Cattle in Central Japan. Anim. Sci. J. 2019, 90, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Reiten, M.; Rousing, T.; Thomsen, P.; Otten, N.; Forkman, B.; Houe, H.; Sørensen, J.; Kirchner, M.K. Mortality, Diarrhea and Respiratory Disease in Danish Dairy Heifer Calves: Effect of Production System and Season. Prev. Vet. Med. 2018, 155, 21–26. [Google Scholar] [CrossRef]
- Webster, A.J.F. Heat loss from cattle with particular emphasis on the effects of cold. In Heat Loss from Animals and Man. Assessment and Control; Elsevier: Amsterdam, The Netherlands, 1974; pp. 205–231. [Google Scholar] [CrossRef]
- Webster, F.; Gordon, J.G.; McGregor, R. The Cold Tolerance of Beef and Dairy Type Calves in the First Weeks of Life. Anim. Sci. 1978, 26, 85–92. [Google Scholar] [CrossRef]
- National Research Council; Committee on Animal Nutrition; Subcommittee on Dairy Cattle Nutrition. Nutrient Requirements of Dairy Cattle: 2001; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Mader, T.L.; Davis, M.S.; Brown-Brandl, T. Environmental Factors Influencing Heat Stress in Feedlot Cattle1,2. J. Anim. Sci. 2006, 84, 712–719. [Google Scholar] [CrossRef]
- Oosterbaan, R.J. Statistical Significance of Segmented Linear Regression with Break-Point Using Variance Analysis (ANOVA) and F-Tests. 2017, pp. 1–13, Unpublished. Available online: https://www.waterlog.info/segreg.htm (accessed on 11 March 2022).
- Pinto, S.; Hoffmann, G.; Ammon, C.; Amon, T. Critical THI Thresholds Based on the Physiological Parameters of Lactating Dairy Cows. J. Therm. Biol. 2020, 88, 102523. [Google Scholar] [CrossRef]
- Yadav, B.; Singh, G.; Wankar, A. Adaptive Capability as Indicated by Redox Status and Endocrine Responses in Crossbred Cattle Exposed to Thermal Stress. J. Anim. Res. 2015, 5, 67. [Google Scholar] [CrossRef]
- Wang, S.; Li, Q.; Peng, J.; Niu, H. Effects of Long-Term Cold Stress on Growth Performance, Behavior, Physiological Parameters, and Energy Metabolism in Growing Beef Cattle. Animals 2023, 13, 1619. [Google Scholar] [CrossRef]
- Berian, S.; Gupta, S.K.; Ali, S.; Dua, S.; Ganaie, I.; Kumar, A. Effect of Cold Stress on Milk Yield, Physiological and Hemato-Biochemical Profile of Cross Bred Dairy Cattle. J. Anim. Res. 2019, 9, 335–338. [Google Scholar] [CrossRef]
- Li, P.; Li, Q.; Chen, D.; He, H.; Ji, X.; Gao, D.; Gao, A. Comparative study on physiological, serum biochemical indexes and immune performance of Dorper sheep, Suffolk sheep and Mongolian sheep under low temperature environment. China Anim. Husb. Vet. Med. 2021, 48, 109–116. [Google Scholar]
- Yadav, B.; Singh, G.; Wankar, A. The Use of Infrared Skin Temperature Measurements for Monitoring Heat Stress and Welfare of Crossbred Cattle. Indian J. Dairy Sci. 2017, 70, 127–131. [Google Scholar]
- Bharati, J.; Dangi, S.S.; Mishra, S.R.; Chouhan, V.S.; Verma, V.; Shankar, O.; Bharti, M.K.; Paul, A.; Mahato, D.K.; Rajesh, G.; et al. Expression Analysis of Toll like Receptors and Interleukins in Tharparkar Cattle during Acclimation to Heat Stress Exposure. J. Therm. Biol. 2017, 65, 48–56. [Google Scholar] [CrossRef] [PubMed]
- 47. Jose, B.; Samad, H.A.; Bharati, J.; Tejaswi, V.; Konda, P.; Sharun, K.; Tripathi, M.K.; Kumar, S.; Punetha, M.; Mohan, D.; et al. Evaluation of Thermo-Adaptability between Tharparkar (Bos indicus) and Crossbred (Bos indicus X Bos taurus) Calves in a Controlled Environment. J. Therm. Biol. 2022, 110, 103381. [Google Scholar] [CrossRef]
- Ouellet, V.; Toledo, I.M.; Dado-Senn, B.; Dahl, G.E.; Laporta, J. Critical Temperature-Humidity Index Thresholds for Dry Cows in a Subtropical Climate. Front. Anim. Sci. 2021, 2, 706636. [Google Scholar] [CrossRef]
- Schalm, O.W.; Feldman, B.F.; Zinkl, J.G.; Jain, N.C. Schalm’s Veterinary Hematology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Kumar, M.; Kaur, H.; Mani, V.; Deka, R.S.; Tyagi, A.K.; Chandra, G.; Dang, A.K.; Kushwaha, R. Supplemental Chromium in Cold-Stressed Buffalo Calves (Bubalus bubalis): Effects on Growth Performance, Nutrient Utilization and Cell Mediated and Humoral Immune Response. Vet. Arh. 2017, 87, 441–456. [Google Scholar] [CrossRef]
- Broucek, J.; Kisac, P.; Uhrincat, M. Effect of Hot Temperatures on the Hematological Parameters, Health and Performance of Calves. Int. J. Biometeorol. 2009, 53, 201–208. [Google Scholar] [CrossRef]
- Tej, J.N.K.; Varma, G.G.; Abhina, M.; Adarsh, N.; Karthiayini, K.; Gleeja, V.L. Impact of Season on Haematological and Biochemical Parameters of Crossbred Female Calves in Hot and Humid Tropics. Biol. Rhythm Res. 2017, 49, 267–275. [Google Scholar] [CrossRef]
- Park, D.S.; Gu, B.-H.; Park, Y.J.; Joo, S.S.; Lee, S.-S.; Kim, S.-H.; Kim, E.T.; Kim, D.H.; Lee, S.S.; Lee, S.J.; et al. Dynamic Changes in Blood Immune Cell Composition and Function in Holstein and Jersey Steers in Response to Heat Stress. Cell Stress Chaperones 2021, 26, 705–720. [Google Scholar] [CrossRef]
- Velayudhan, S.M.; Brügemann, K.; Alam, S.; Yin, T.; Devaraj, C.; Sejian, V.; Schlecht, E.; König, S. Molecular, Physiological and Hematological Responses of Crossbred Dairy Cattle in a Tropical Savanna Climate. Biology 2022, 12, 26. [Google Scholar] [CrossRef]
- Abera, M.; Mummed, Y.Y.; Eshetu, M.; Pilla, F.; Wondifraw, Z. Physiological, Biochemical, and Growth Parameters of Fogera Cattle Calves to Heat Stress during Different Seasons in Sub-Humid Part of Ethiopia. Animals 2021, 11, 1062. [Google Scholar] [CrossRef] [PubMed]
- Lacetera, N.; Bernabucci, U.; Scalia, D.; Basiricò, L.; Morera, P.; Nardone, A. Heat Stress Elicits Different Responses in Peripheral Blood Mononuclear Cells from Brown Swiss and Holstein Cows. J. Dairy Sci. 2006, 89, 4606–4612. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Hooda, O.K.; Kumar, S. Impact of Heat Stress and Hypercapnia on Physiological, Hematological, and Behavioral Profile of Tharparkar and Karan Fries Heifers. Vet. World 2017, 10, 1149–1155. [Google Scholar] [CrossRef]
- Bagath, M.; Krishnan, G.; Devaraj, C.; Rashamol, V.P.; Pragna, P.; Lees, A.M.; Sejian, V. The Impact of Heat Stress on the Immune System in Dairy Cattle: A Review. Res. Vet. Sci. 2019, 126, 94–102. [Google Scholar] [CrossRef]
- Wankar, A.K.; Singh, G.; Yadav, B. Thermoregulatory and Adaptive Responses of Adult Buffaloes (Bubalus bubalis) During Hyperthermia: Physiological, Behavioral, and Metabolic Approach. Vet. World 2014, 7, 825–830. [Google Scholar] [CrossRef]
- Kim, W.S.; Peng, D.Q.; Jo, Y.H.; Nejad, J.G.; Lee, H.G. Responses of Beef Calves to Long-Term Heat Stress Exposure by Evaluating Growth Performance, Physiological, Blood and Behavioral Parameters. J. Therm. Biol. 2021, 100, 103033. [Google Scholar] [CrossRef]
- Kim, W.-S.; Lee, H.G.; Lee, H.-G. Impact of Cold Stress on Physiological, Endocrinological, Immunological, Metabolic, and Behavioral Changes of Beef Cattle at Different Stages of Growth. Animals 2023, 13, 1073. [Google Scholar] [CrossRef]
- Mishra, S.R. Significance of Molecular Chaperones and Micro RNAs in Acquisition of Thermo-Tolerance in Dairy Cattle. Anim. Biotechnol. 2020, 33, 765–775. [Google Scholar] [CrossRef]
- Neuwirth, J.G.; Norton, J.K.; Rawlings, C.A.; Thompson, F.N.; Ware, G.O. Physiologic Responses of Dairy Calves to Environmental Heat Stress. Int. J. Biometeorol. 1979, 23, 243–254. [Google Scholar] [CrossRef]
- Lemal, P.; May, K.; König, S.; Schroyen, M.; Gengler, N. Invited Review: From Heat Stress to Disease—Immune Response and Candidate Genes Involved in Cattle Thermotolerance. J. Dairy Sci. 2023, 106, 4471–4488. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Bonner, S.L.; Loxton, I.; Mader, T.L. Effects of Chronic Heat Stress on Plasma Concentration of Secreted Heat Shock Protein 70 in Growing Feedlot Cattle1. J. Anim. Sci. 2013, 91, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Dangi, S.S.; Gupta, M.; Nagar, V.; Yadav, V.P.; Dangi, S.K.; Shankar, O.; Chouhan, V.S.; Kumar, P.; Singh, G.; Sarkar, M. Impact of Short-Term Heat Stress on Physiological Responses and Expression Profile of HSPs in Barbari Goats. Int. J. Biometeorol. 2014, 58, 2085–2093. [Google Scholar] [CrossRef] [PubMed]
- Umar, S.I.U.; Konwar, D.; Khan, A.; Bhat, M.A.; Javid, F.; Jeelani, R.; Nabi, B.; Najar, A.A.; Kumar, D.; Brahma, B. Delineation of Temperature-Humidity Index (THI) as Indicator of Heat Stress in Riverine Buffaloes (Bubalus bubalis) of a Sub-Tropical Indian Region. Cell Stress Chaperones 2021, 26, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Khalilvandi-Behroozyar, H.; Mohtashami, B.; Dehghan-Banadaky, M.; Kazemi-Bonchenari, M.; Ghaffari, M.H. Effects of Fat Source in Calf Starter on Growth Performance, Blood Fatty Acid Profiles, and Inflammatory Markers during Cold Season. Sci. Rep. 2023, 13, 18627. [Google Scholar] [CrossRef]
- Nonnecke, B.J.; Foote, M.R.; Miller, B.L.; Fowler, M.; Johnson, T.E.; Horst, R.L. Effects of Chronic Environmental Cold on Growth, Health, and Select Metabolic and Immunologic Responses of Preruminant Calves. J. Dairy Sci. 2009, 92, 6134–6143. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Rashamol, V.P.; Bagath, M.; Sejian, V.; Dunshea, F.R. Impacts of Heat Stress on Immune Responses and Oxidative Stress in Farm Animals and Nutritional Strategies for Amelioration. Int. J. Biometeorol. 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Tao, S.; Connor, E.E.; Bubolz, J.W.; Thompson, I.M.; do Amaral, B.C.; Hayen, M.J.; Dahl, G.E. Short Communication: Effect of Heat Stress during the Dry Period on Gene Expression in Mammary Tissue and Peripheral Blood Mononuclear Cells. J. Dairy Sci. 2013, 96, 378–383. [Google Scholar] [CrossRef]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited Review: Genes Involved in the Bovine Heat Stress Response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef]
- Wathes, C.M.; Jones, C.R.; Webster, A.R. Ventilation, Air Hygiene and Animal Health. Vet. Rec. 1983, 113, 554–559. [Google Scholar]
- Holmes, C.W.; Christensen, R.; McLean, N.A.; Lockyer, J. Effects of Winter Weather on the Growth Rate and Heat Production of Dairy Cattle. N. Z. J. Agric. Res. 1978, 21, 549–556. [Google Scholar] [CrossRef]
- Spain, J.N.; Spiers, D.E. Effects of Supplemental Shade on Thermoregulatory Response of Calves to Heat Challenge in a Hutch Environment. J. Dairy Sci. 1996, 79, 639–646. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attributes | Total Mixed Ration |
|---|---|
| Organic matter | 91.18 |
| Crude protein | 17.36 |
| Ether extract | 1.46 |
| Neutral detergent fiber | 66.73 |
| Acid detergent fiber | 17.03 |
| Hemicellulose | 20.31 |
| Total carbohydrates | 77.86 |
| Calcium | 0.87 |
| Phosphorus | 0.48 |
| First Phase | Second Phase | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Psychrometric Chamber | Antechamber | Psychrometric Chamber | Antechamber | ||||||||
| T (°C) | RH (%) | THI | T (°C) | Average RH (%) | T (°C) | RH (%) | THI | T (°C) | Average RH (%) | ||
| Max | Min | Max | Min | ||||||||
| 21 | 60 | 67.16 | 26.73 | 21.31 | 68.2 | 24 | 70 | 72.32 | 28.68 | 22.72 | 68.6 |
| 24 | 70 | 72.32 | 29.68 | 23.22 | 65.2 | 21 | 60 | 67.16 | 26.73 | 21.31 | 68.2 |
| 27 | 75 | 77.45 | 31.50 | 24.86 | 62.1 | 18 | 60 | 62.96 | 22.42 | 15.24 | 70.2 |
| 30 | 75 | 82.10 | 33.53 | 26.38 | 59.3 | 15 | 60 | 58.76 | 20.54 | 13.60 | 75.2 |
| 33 | 75 | 86.75 | 34.82 | 26.64 | 75.3 | 12 | 60 | 54.56 | 19.68 | 10.82 | 80.2 |
| 36 | 72 | 90.75 | 34.96 | 27.21 | 81.4 | 9 | 60 | 50.36 | 16.54 | 7.25 | 82.5 |
| 39 | 70 | 94.82 | 35.64 | 28.32 | 83.6 | -- | -- | -- | -- | -- | -- |
| S. No | Parameter | Lower Threshold THI | Upper Threshold THI |
|---|---|---|---|
| 1 | Respiratory Rate | 56.73 | 82.35 |
| 2 | Pulse Rate | 58.92 | No Breakpoint |
| 3 | Rectal Temperature | 58.92 | 82.35 |
| 4 | Total Red Blood Cells | No Breakpoint | 82.35 |
| 5 | Hemoglobin | No Breakpoint | 82.35 |
| 6 | PCV | No Breakpoint | 82.35 |
| 7 | Total Leukocytic Count | 54.97 | 82.35 |
| 8 | Granulocyte % | 63.10 | 86.94 |
| 9 | Lymphocyte % | No Breakpoint | 86.94 |
| 10 | AST | 54.75 | 88.98 |
| 11 | ALT | 54.97 | 82.35 |
| 12 | Cortisol | 54.97 | 84.29 |
| 13 | HSP70 | No Breakpoint | No Breakpoint |
| 14 | HSP90 | 58.92 | 86.94 |
| 15 | TNFα | No Breakpoint | No Breakpoint |
| 16 | IL6 | 58.92 | 85.67 |
| 17 | Triglyceride | 58.92 | No Breakpoint |
| 18 | Urea | 54.97 | No Breakpoint |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, B.; Srivastava, A.; Yadav, P.; Swain, D.K.; Anand, M.; Yadav, S.; Madan, A.K. Identification of Thermoneutral Zone in Sahiwal Zebu Calves in Subtropical Climate of India. Animals 2025, 15, 1830. https://doi.org/10.3390/ani15131830
Yadav B, Srivastava A, Yadav P, Swain DK, Anand M, Yadav S, Madan AK. Identification of Thermoneutral Zone in Sahiwal Zebu Calves in Subtropical Climate of India. Animals. 2025; 15(13):1830. https://doi.org/10.3390/ani15131830
Chicago/Turabian StyleYadav, Brijesh, Anandita Srivastava, Poonam Yadav, Dilip Kumar Swain, Mukul Anand, Sarvajeet Yadav, and Arun Kumar Madan. 2025. "Identification of Thermoneutral Zone in Sahiwal Zebu Calves in Subtropical Climate of India" Animals 15, no. 13: 1830. https://doi.org/10.3390/ani15131830
APA StyleYadav, B., Srivastava, A., Yadav, P., Swain, D. K., Anand, M., Yadav, S., & Madan, A. K. (2025). Identification of Thermoneutral Zone in Sahiwal Zebu Calves in Subtropical Climate of India. Animals, 15(13), 1830. https://doi.org/10.3390/ani15131830