
Effects of Alpha-Ketoglutarate Supplementation on Growth Performance, Diarrhea Incidence, Plasma Amino Acid, and Nutrient Digestibility in Weaned Piglets
 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Approval
2.2. Experimental Design
2.3. Growth Performance and Diarrhea Incidence
2.4. Apparent Digestibility of Nutrients
2.5. Plasma Biochemical Indices
2.6. Plasma Amino Acid Level
2.7. Statistical Analysis
3. Results
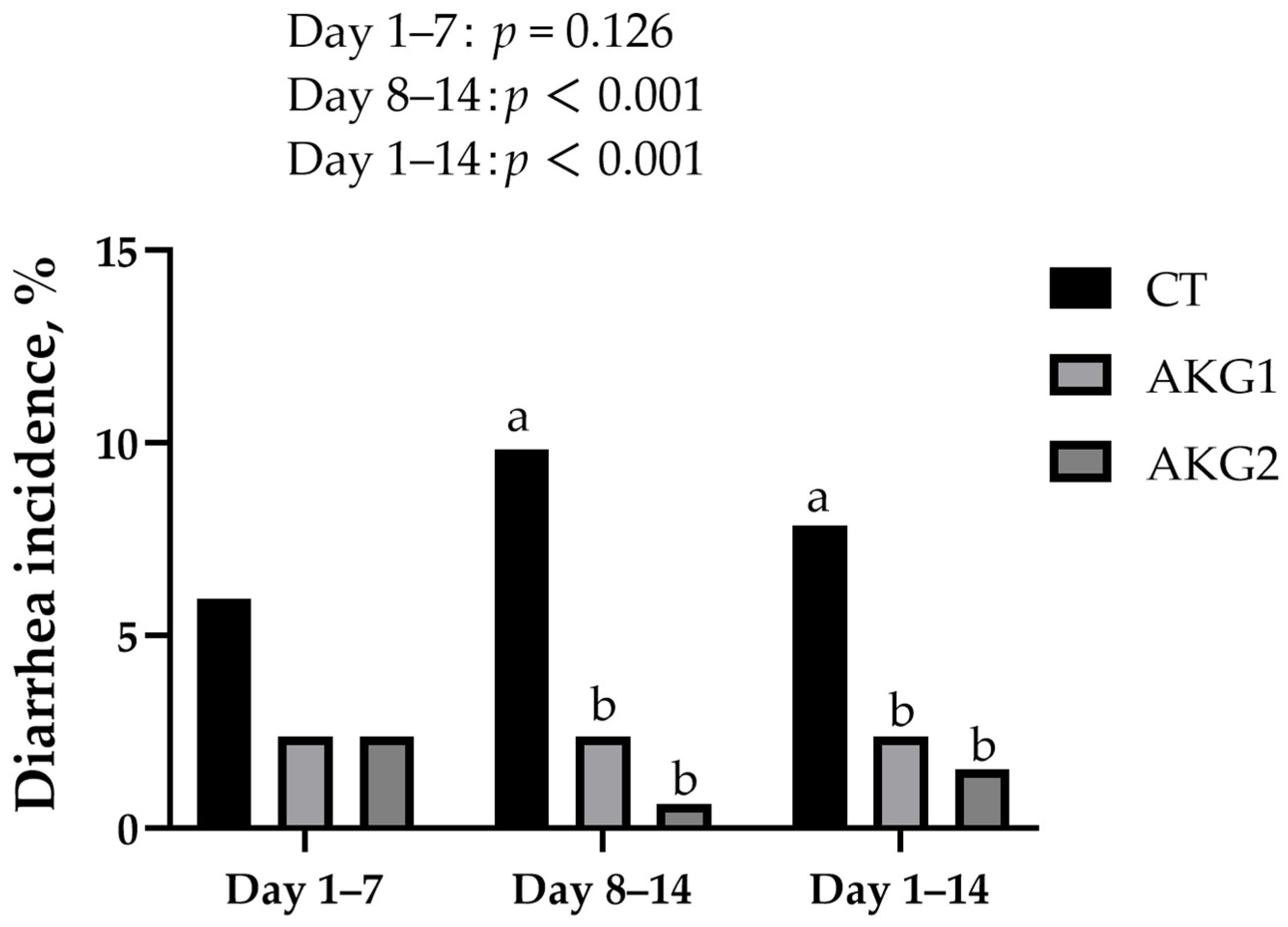
3.1. Growth Performance and Diarrhea Incidence of Weaned Piglets
3.2. Nutrient Apparent Digestibility of Weaned Piglets
3.3. Plasma Biochemistry Parameters of Weaned Piglets
3.4. Plasma Amino Acid Level of Weaned Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADFI | Average Daily Feed Intake |
| ADG | Average Daily Gain |
| AKG | Alpha-ketoglutaric acid |
| Ala | Alanine acid |
| ALB | Albumin |
| ALP | Alkaline Phosphatase |
| Arg | Arginine acid |
| Asp | Aspartic acid |
| AST | Aspartate Aminotransferase |
| FCR | Feed Conversion Ratio |
| Glu | Glutamic acid |
| GLU | Glucose |
| Gly | Glycine acid |
| HDL | High-Density Lipoprotein |
| His | Histidine acid |
| Ile | Isoleucine acid |
| LDL | Low-Density Lipoprotein |
| Leu | Leucine acid |
| Lys | Lysine acid |
| Met | Methionine acid |
| Phe | Phenylalanine acid |
| Ser | Serine acid |
| TC | Total Cholesterol |
| TG | Triglycerides |
| Thr | Threonine acid |
| TP | Total Protein |
| Trp | Tryptophan acid |
| Tyr | Tyrosine acid |
| Val | Valine acid |
References
- Baldwin, J.E.; Krebs, H. The evolution of metabolic cycles. Nature 1981, 291, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Lambert, B.D.; Filip, R.; Stoll, B.; Junghans, P.; Derno, M.; Hennig, U.; Souffrant, W.B.; Pierzynowski, S.; Burrin, D.G. First-pass metabolism limits the intestinal absorption of enteral α-ketoglutarate in young pigs. J. Nutr. 2006, 136, 2779–2784. [Google Scholar] [CrossRef] [PubMed]
- Pierzynowski, S.; Sjodin, A. Perspectives of glutamine and its derivatives as feed additives for farm animals. J. Anim. Feed. Sci. 1998, 7, 79–91. [Google Scholar] [CrossRef]
- Kristensen, N.B.; Jungvid, H.; Fernández, J.A.; Pierzynowski, S. Absorption and metabolism of α-ketoglutarate in growing pigs. J. Anim. Physiol. Anim. Nutr. 2002, 86, 239–245. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Ding, B.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Kang, P.; Yin, Y.; Wu, G. Alpha-ketoglutarate and intestinal function. Front. Biosci. 2011, 16, 1186–1196. [Google Scholar] [CrossRef]
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca, Argentina, 2021. [Google Scholar]
- Filip, R.; Wdowiak, L.; Harrison, A.P.; Pierzynowski, S.G. Dietary supplementation with phytohemagglutinin in combination with alpha-ketoglutarate limits the excretion of nitrogen via urinary tract. Ann. Agric. Environ. Med. 2008, 15, 309–315. [Google Scholar] [PubMed]
- Yao, K.; Yin, Y.; Li, X.; Xi, P.; Wang, J.; Lei, J.; Hou, Y.; Wu, G. Alpha-ketoglutarate inhibits glutamine degradation and enhances protein synthesis in intestinal porcine epithelial cells. Amino Acids 2012, 42, 2491–2500. [Google Scholar] [CrossRef]
- Żurek, A.; Mizerska-Kowalska, M.; Sławińska-Brych, A.; Kaławaj, K.; Bojarska-Junak, A.; Kandefer-Szerszeń, M.; Zdzisińska, B. Alpha ketoglutarate exerts a pro-osteogenic effect in osteoblast cell lines through activation of JNK and mTOR/S6K1/S6 signaling pathways. Toxicol. Appl. Pharmacol. 2019, 374, 53–64. [Google Scholar] [CrossRef]
- Andersen, N.; Tatara, M.; Krupski, W.; Majcher, P.; Harrison, A. The long-term effect of α-ketoglutarate, given early in postnatal life, on both growth and various bone parameters in pigs. J. Anim. Physiol. Anim. Nutr. 2008, 92, 519–528. [Google Scholar] [CrossRef]
- Tekwe, C.D.; Yao, K.; Lei, J.; Li, X.; Gupta, A.; Luan, Y.; Meininger, C.J.; Bazer, F.W.; Wu, G. Oral administration of α-ketoglutarate enhances nitric oxide synthesis by endothelial cells and whole-body insulin sensitivity in diet-induced obese rats. Exp. Biol. Med. 2019, 244, 1081–1088. [Google Scholar] [CrossRef]
- Roe, D.S.; Roe, C.R.; Brivet, M.; Sweetman, L. Evidence for a short-chain carnitine–acylcarnitine translocase in mitochondria specifically related to the metabolism of branched-chain amino acids. Mol. Genet. Metab. 2000, 69, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.J.; Kristal, B.S. Multiple roles of glutathione in the central nervous system. Biol. Chem. Hoppe Seyler. 1997, 378, 793–802. [Google Scholar]
- Wang, L.; Hou, Y.; Yi, D.; Li, Y.; Ding, B.; Zhu, H.; Liu, J.; Xiao, H.; Wu, G. Dietary supplementation with glutamate precursor α-ketoglutarate attenuates lipopolysaccharide-induced liver injury in young pigs. Amino Acids 2015, 47, 1309–1318. [Google Scholar] [CrossRef]
- Filip, R.; Pierzynowski, S.G. The role of glutamine and alfa-ketoglutarate in gut metabolism and the potential application in medicine and nutrition. J. Pre-Clin. Clin. Res. 2007, 1, 9–15. [Google Scholar]
- He, L.; Li, H.; Huang, N.; Tian, J.; Liu, Z.; Zhou, X.; Yao, K.; Li, T.; Yin, Y. Effects of alpha-ketoglutarate on glutamine metabolism in piglet enterocytes in vivo and in vitro. J. Agric. Food Chem. 2016, 64, 2668–2673. [Google Scholar] [CrossRef]
- Tian, J.; Yang, F.; Bao, X.; Jiang, Q.; Li, Y.; Yao, K.; Yin, Y. Dietary alpha-ketoglutarate supplementation improves bone growth, phosphorus digestion, and growth performance in piglets. Animals 2023, 13, 569. [Google Scholar] [CrossRef]
- He, L.; Zhou, X.; Huang, N.; Li, H.; Cui, Z.; Tian, J.; Jiang, Q.; Liu, S.; Wu, J.; Li, T. Administration of alpha-ketoglutarate improves epithelial restitution under stress injury in early-weaning piglets. Oncotarget 2017, 8, 91965. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, H.; Long, L.; Zhao, Y.; Jiang, Q.; Wu, F.; Kang, B.; Liu, S.; Adebowale, T.O.; Fu, C. The effects of dietary supplementation with α-ketoglutarate on the intestinal microbiota, metabolic profiles, and ammonia levels in growing pigs. Anim. Feed. Sci. Technol. 2017, 234, 321–328. [Google Scholar] [CrossRef]
- Zhou, B.; Magana, L.; Hong, Z.; Huang, L.S.; Chakraborty, S.; Tsukasaki, Y.; Huang, C.; Wang, L.; Di, A.; Ganesh, B. The angiocrine Rspondin3 instructs interstitial macrophage transition via metabolic–epigenetic reprogramming and resolves inflammatory injury. Nat. Immunol. 2020, 21, 1430–1443. [Google Scholar] [CrossRef]
- Liu, M.; Chen, Y.; Wang, S.; Zhou, H.; Feng, D.; Wei, J.; Shi, X.; Wu, L.; Zhang, P.; Yang, H. α-Ketoglutarate modulates macrophage polarization through regulation of PPARγ transcription and mTORC1/p70S6K pathway to ameliorate ALI/ARDS. Shock 2020, 53, 103–113. [Google Scholar] [CrossRef]
- Council, N.R.; Earth, D.o.; Studies, L.; Committee on Nutrient Requirements of Swine; Division on Earth and Life Studies; National Research Council. Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; Pub AOAC International: Rockville, MD, USA, 2005. [Google Scholar]
- Newkirk, R.; Classen, H.; Scott, T.; Edney, M. The digestibility and content of amino acids in toasted and non-toasted canola meals. Can. J. Anim. Sci. 2003, 83, 131–139. [Google Scholar] [CrossRef]
- Dai, Z.; Wu, Z.; Jia, S.; Wu, G. Analysis of amino acid composition in proteins of animal tissues and foods as pre-column o-phthaldialdehyde derivatives by HPLC with fluorescence detection. J. Chromatogr. B 2014, 964, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.; James, P.; Smith, M.; Bourne, F. Effect of weaning on the capacity of pig intestinal villi to digest and absorb nutrients. J. Agric. Sci. 1986, 107, 579–590. [Google Scholar] [CrossRef]
- Nabuurs, M.; Hoogendoorn, A.; Van Der Molen, E.; Van Osta, A. Villus height and crypt depth in weaned and unweaned pigs, reared under various circumstances in the Netherlands. Res. Vet. Sci. 1993, 55, 78–84. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Huang, N.; Li, H.; Tian, J.; Zhou, X.; Li, T.; Yao, K.; Wu, G.; Yin, Y. AMPK/α-ketoglutarate axis regulates intestinal water and ion homeostasis in young pigs. J. Agric. Food Chem. 2017, 65, 2287–2298. [Google Scholar] [CrossRef]
- Chen, J.; Wu, F.; Yang, H.; Li, F.; Jiang, Q.; Liu, S.; Kang, B.; Li, S.; Adebowale, T.; Huang, N. Growth performance, nitrogen balance, and metabolism of calcium and phosphorus in growing pigs fed diets supplemented with alpha-ketoglutarate. Anim. Feed. Sci. Technol. 2017, 226, 21–28. [Google Scholar] [CrossRef]
- Wang, L.; Fan, Z.; Wu, D.; Li, J.; Xu, Q.; Miao, L.; Ge, X.; Cao, D.; Zheng, X. Effects of dietary α-ketoglutarate on the growth performance, digestive enzymes, tor signaling pathway and intestinal microbiota of juvenile mirror carp (Cyprinus carpio) fed low phosphorus diets. Aquaculture 2023, 574, 739736. [Google Scholar] [CrossRef]
- Chen, L.; Li, P.; Wang, J.; Li, X.; Gao, H.; Yin, Y.; Hou, Y.; Wu, G. Catabolism of nutritionally essential amino acids in developing porcine enterocytes. Amino Acids 2009, 37, 143–152. [Google Scholar] [CrossRef]
- Jones, C.; Palmer, T.A.; Griffiths, R. Randomized clinical outcome study of critically ill patients given glutamine-supplemented enteral nutrition. Nutrition 1999, 15, 108–115. [Google Scholar] [CrossRef]
- Zhao, L.; Guo, H.; Sun, H. Effects of low-protein diet supplementation with alpha-ketoglutarate on growth performance, nitrogen metabolism and mTOR signalling pathway of skeletal muscle in piglets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 300–309. [Google Scholar] [CrossRef]
- Ai, F.; Wang, L.; Li, J.; Xu, Q. Effects of a-ketoglutarate (AKG) supplementation in low phosphorous diets on the growth, phosphorus metabolism and skeletal development of juvenile mirror carp (Cyprinus carpio). Aquaculture 2019, 507, 393–401. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Huang, N.; Zhou, X.; Tian, J.; Li, T.; Wu, J.; Tian, Y.; Yin, Y.; Yao, K. Alpha-ketoglutarate suppresses the NF-κB-mediated inflammatory pathway and enhances the PXR-regulated detoxification pathway. Oncotarget 2017, 8, 102974. [Google Scholar] [CrossRef] [PubMed]
- Barter, P.; Gotto, A.M.; LaRosa, J.C.; Maroni, J.; Szarek, M.; Grundy, S.M.; Kastelein, J.J.; Bittner, V.; Fruchart, J.-C. HDL cholesterol, very low levels of LDL cholesterol, and cardiovascular events. N. Engl. J. Med. 2007, 357, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Radzki, R.P.; Bieńko, M.; Pierzynowski, S.G. Effect of dietary alpha-ketoglutarate on blood lipid profile during hypercholesterolaemia in rats. Scand. J. Clin. Lab. Investig. 2009, 69, 175–180. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhu, C.; Wang, Y.; Sun, J.; Feng, J.; Ma, Z.; Li, P.; Peng, W.; Yin, C.; Xu, G. α-Ketoglutaric acid ameliorates hyperglycemia in diabetes by inhibiting hepatic gluconeogenesis via serpina1e signaling. Sci. Adv. 2022, 8, eabn2879. [Google Scholar] [CrossRef]
- Wu, G. Intestinal mucosal amino acid catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]

{kind=link}
| Pre-Conservation Period | Late Conservation Period | |||||
|---|---|---|---|---|---|---|
| CT | AKG1 | AKG2 | CT | AKG1 | AKG2 | |
| Corn | 46.46 | 46.46 | 46.46 | 60.3 | 60.3 | 60.3 |
| Soybean meal, 46% | 13.2 | 13.2 | 13.2 | 17.3 | 17.3 | 17.3 |
| Expanded soybean | 14.5 | 14.5 | 14.5 | 6.5 | 6.5 | 6.5 |
| Fish meal | 6 | 6 | 6 | 4 | 4 | 4 |
| Whey powder | 15 | 15 | 15 | 5 | 5 | 5 |
| Soybean oil | 1.1 | 1.1 | 1.1 | 1.5 | 1.5 | 1.5 |
| Bran | 0.112 | 0.112 | 0.112 | 0.02 | 0.02 | 0.02 |
| Calcium dihydrogen phosphate | 0.35 | 0.35 | 0.35 | 0 | 0 | 0 |
| Limestone | 0.77 | 0.77 | 0.77 | 0.6 | 0.6 | 0.6 |
| Salt | 0.4 | 0.4 | 0.4 | 1 | 1 | 1 |
| Choline chloride, 60% | 0.05 | 0.05 | 0.05 | 0.4 | 0.4 | 0.4 |
| L-Lysine HCL, 65% | 0.9 | 0.9 | 0.9 | 0.05 | 0.05 | 0.05 |
| DL-Met | 0.06 | 0.06 | 0.06 | 0.65 | 0.65 | 0.65 |
| Threonine | 0.24 | 0.24 | 0.24 | 0.05 | 0.05 | 0.05 |
| Tryptophan | 0.02 | 0.02 | 0.02 | 0.14 | 0.14 | 0.14 |
| Valine | 0 | 0 | 0 | 0.01 | 0.01 | 0.01 |
| Isoleucine | 0 | 0 | 0 | 0.6 | 0.6 | 0.6 |
| Phytase (10,000) | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
| Acidifier | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Sodium butyrate | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Alpha-ketoglutarate | 0 | 0.05 | 0.1 | 0 | 0.1 | 0.2 |
| Premix 1 | 0.448 | 0.448 | 0.448 | 0.268 | 0.268 | 0.268 |
| 100.00 | 100.05 | 100.10 | 100 | 100.1 | 100.2 | |
| Nutrition composition | ||||||
| Analyzed value | ||||||
| CP, % | 19.64 | 19.55 | 19.68 | 17.97 | 18.04 | 18.10 |
| Ca, % | 0.82 | 0.83 | 0.83 | 0.73 | 0.73 | 0.72 |
| p, % | 0.53 | 0.53 | 0.57 | 0.47 | 0.48 | 0.47 |
| EE, % | 4.34 | 4.65 | 4.72 | 5.92 | 6.35 | 6.45 |
| Ash, % | 5.82 | 5.84 | 5.84 | 11.20 | 11.02 | 11.18 |
| Calculated value | ||||||
| ME, kcal | 3350 | 3350 | 3350 | 3300 | 3300 | 3300 |
| Lys, % | 1.25 | 1.25 | 1.25 | 1.22 | 1.22 | 1.22 |
| Met, % | 0.36 | 0.36 | 0.36 | 0.35 | 0.35 | 0.35 |
| Thr, % | 0.74 | 0.74 | 0.74 | 0.72 | 0.72 | 0.72 |
| Trp, % | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 | 0.21 |
| AKG Level, mg/kg | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 500 | 1000 | SEM | ANOVA | Linear | Quadratic | |
| BW, kg | |||||||
| Day 0 | 7.33 | 7.33 | 7.33 | 0.51 | 1.000 | 0.998 | 0.999 |
| Day 14 | 11.75 | 11.45 | 12.03 | 0.77 | 0.862 | 0.796 | 0.635 |
| Day 28 | 19.05 | 19.13 | 19.55 | 0.87 | 0.922 | 0.703 | 0.903 |
| Day 42 | 27.81 | 27.97 | 28.48 | 1.12 | 0.835 | 0.552 | 0.991 |
| ADG, g | |||||||
| Day 0–14 | 315 | 294 | 336 | 24 | 0.469 | 0.555 | 0.282 |
| Day 14–28 | 522 | 548 | 537 | 26 | 0.780 | 0.684 | 0.568 |
| Day 28–42 | 626 | 632 | 638 | 30 | 0.675 | 0.402 | 0.783 |
| Day 0–42 | 487 | 491 | 503 | 18 | 0.710 | 0.411 | 0.989 |
| ADFI, g | |||||||
| Day 0–14 | 432 | 398 | 443 | 29 | 0.633 | 0.754 | 0.369 |
| Day 14–28 | 793 | 852 | 813 | 30 | 0.970 | 0.811 | 0.960 |
| Day 28–42 | 1104 | 1115 | 1122 | 53 | 0.776 | 0.664 | 0.575 |
| Day 0–42 | 777 | 788 | 793 | 30 | 0.913 | 0.673 | 0.963 |
| FCR | |||||||
| Day 0–14 | 1.394 | 1.372 | 1.319 | 0.052 | 0.656 | 0.398 | 0.730 |
| Day 14–28 | 1.526 | 1.569 | 1.513 | 0.039 | 0.762 | 0.664 | 0.555 |
| Day 28–42 | 1.763 | 1.770 | 1.759 | 0.030 | 0.440 | 0.291 | 0.471 |
| Day 0–42 | 1.593 | 1.607 | 1.572 | 0.024 | 0.587 | 0.307 | 0.928 |
| AKG Level, mg/kg | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 500 | 1000 | SEM | ANOVA | Linear | Quadratic | |
| Dry matter | 78.35 ab | 77.45 b | 79.70 a | 0.56 | 0.045 | 0.118 | 0.042 |
| Crude protein | 74.73 | 74.79 | 77.47 | 0.90 | 0.092 | 0.056 | 0.269 |
| Ether extract | 69.73 | 70.48 | 74.45 | 1.55 | 0.113 | 0.054 | 0.423 |
| Gross energy | 80.30 | 79.76 | 81.62 | 0.56 | 0.092 | 0.123 | 0.107 |
| AKG Level, mg/kg | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 500 | 1000 | SEM | ANOVA | Linear | Quadratic | |
| Day 14 | |||||||
| ALB, g/L | 22.13 | 21.52 | 23.24 | 1.07 | 0.537 | 0.481 | 0.393 |
| ALP, IU/L | 297 | 264 | 407 | 41 | 0.062 | 0.076 | 0.099 |
| ALT, IU/L | 49.50 | 55.26 | 51.12 | 4.27 | 0.640 | 0.797 | 0.370 |
| AST, IU/L | 87.96 | 91.15 | 82.38 | 10.45 | 0.852 | 0.726 | 0.665 |
| GLU, mmol/L | 3.42 | 3.74 | 3.53 | 0.41 | 0.862 | 0.852 | 0.614 |
| HDL, mmol/L | 0.40 | 0.51 | 0.51 | 0.05 | 0.208 | 0.116 | 0.412 |
| LDL, mmol/L | 0.90 | 1.05 | 1.00 | 0.08 | 0.494 | 0.451 | 0.363 |
| TC, mmol/L | 1.59 | 1.83 | 1.81 | 0.13 | 0.396 | 0.262 | 0.446 |
| TG, mmol/L | 0.74 | 0.68 | 0.68 | 0.09 | 0.899 | 0.702 | 0.808 |
| TP, g/L | 48.49 | 49.88 | 49.96 | 2.31 | 0.884 | 0.663 | 0.822 |
| Day 42 | |||||||
| ALB, g/L | 26.62 ab | 24.01 b | 30.79 a | 1.66 | 0.046 | 0.114 | 0.045 |
| ALP, IU/L | 200 | 199 | 249 | 21 | 0.199 | 0.123 | 0.352 |
| ALT, IU/L | 55.89 | 51.66 | 49.77 | 5.26 | 0.736 | 0.452 | 0.867 |
| AST, IU/L | 66.97 | 83.80 | 55.72 | 19.15 | 0.674 | 0.725 | 0.422 |
| GLU, mmol/L | 6.08 ab | 5.30 b | 6.39 a | 0.23 | 0.015 | 0.368 | 0.006 |
| HDL, mmol/L | 0.60 | 0.62 | 0.75 | 0.05 | 0.125 | 0.062 | 0.413 |
| LDL, mmol/L | 1.15 | 1.26 | 1.27 | 0.10 | 0.626 | 0.394 | 0.665 |
| TC, mmol/L | 2.08 | 2.24 | 2.36 | 0.12 | 0.341 | 0.151 | 0.886 |
| TG, mmol/L | 1.05 | 0.70 | 0.62 | 0.17 | 0.423 | 0.222 | 0.660 |
| TP, g/L | 62.76 | 60.04 | 63.12 | 1.76 | 0.454 | 0.892 | 0.219 |
| AKG Level, mg/kg | p-Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 500 | 1000 | SEM | ANOVA | Linear | Quadratic | |
| Day 14 | |||||||
| Asp | 1.12 | 1.13 | 1.09 | 0.11 | 0.959 | 0.840 | 0.843 |
| Glu | 12.83 | 12.07 | 12.61 | 1.31 | 0.927 | 0.913 | 0.714 |
| Ser | 8.31 | 6.87 | 7.26 | 0.67 | 0.327 | 0.291 | 0.288 |
| His | 3.01 | 2.42 | 2.81 | 0.27 | 0.350 | 0.627 | 0.177 |
| Gly | 38.16 | 35.76 | 36.70 | 3.48 | 0.890 | 0.774 | 0.704 |
| Thr | 19.97 | 18.24 | 16.55 | 2.61 | 0.666 | 0.375 | 0.995 |
| Arg | 16.16 | 14.98 | 16.66 | 1.22 | 0.623 | 0.780 | 0.359 |
| Ala | 17.01 | 15.20 | 16.41 | 0.88 | 0.373 | 0.644 | 0.190 |
| Tyr | 6.36 | 5.99 | 6.64 | 1.25 | 0.935 | 0.876 | 0.745 |
| Val | 10.74 | 8.70 | 10.77 | 1.08 | 0.350 | 0.985 | 0.154 |
| Met | 5.16 | 5.17 | 4.97 | 0.37 | 0.917 | 0.726 | 0.830 |
| Trp | 3.15 | 2.31 | 2.98 | 0.43 | 0.393 | 0.785 | 0.187 |
| Ile | 8.57 | 7.28 | 7.80 | 0.95 | 0.663 | 0.594 | 0.470 |
| Phe | 5.66 | 3.95 | 4.87 | 0.49 | 0.089 | 0.289 | 0.051 |
| Lys | 18.94 | 16.74 | 15.82 | 1.40 | 0.423 | 0.209 | 0.761 |
| Leu | 9.30 | 7.17 | 8.75 | 1.25 | 0.505 | 0.774 | 0.265 |
| Day 42 | |||||||
| Asp | 1.16 | 1.06 | 1.18 | 0.11 | 0.752 | 0.911 | 0.462 |
| Glu | 14.04 | 13.03 | 13.94 | 1.52 | 0.878 | 0.963 | 0.617 |
| Ser | 6.05 | 5.69 | 5.97 | 0.53 | 0.886 | 0.922 | 0.636 |
| His | 6.29 | 7.09 | 6.44 | 0.73 | 0.722 | 0.892 | 0.434 |
| Gly | 36.27 | 33.03 | 36.45 | 3.11 | 0.693 | 0.968 | 0.400 |
| Thr | 13.49 | 10.78 | 10.40 | 1.30 | 0.284 | 0.150 | 0.519 |
| Arg | 10.72 | 10.19 | 10.25 | 0.94 | 0.920 | 0.744 | 0.815 |
| Ala | 22.68 | 20.41 | 20.30 | 1.68 | 0.611 | 0.385 | 0.646 |
| Tyr | 6.57 | 4.55 | 5.54 | 0.59 | 0.091 | 0.244 | 0.059 |
| Val | 9.42 | 9.15 | 9.57 | 0.86 | 0.949 | 0.909 | 0.768 |
| Met | 4.59 | 3.39 | 4.03 | 0.40 | 0.150 | 0.350 | 0.086 |
| Trp | 7.00 | 6.19 | 7.04 | 0.59 | 0.528 | 0.964 | 0.267 |
| Ile | 8.41 | 8.04 | 7.93 | 0.61 | 0.859 | 0.604 | 0.871 |
| Phe | 6.95 | 7.39 | 6.36 | 0.66 | 0.615 | 0.575 | 0.423 |
| Lys | 20.00 | 17.11 | 17.62 | 1.08 | 0.174 | 0.146 | 0.226 |
| Leu | 11.43 | 10.36 | 11.59 | 0.45 | 0.154 | 0.815 | 0.059 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Han, R.; Xi, H.; Chen, W.; Li, Y.; Zhou, Q.; Li, X.; Huang, K.; Bontempo, V.; Gu, X.; et al. Effects of Alpha-Ketoglutarate Supplementation on Growth Performance, Diarrhea Incidence, Plasma Amino Acid, and Nutrient Digestibility in Weaned Piglets. Animals 2025, 15, 1723. https://doi.org/10.3390/ani15121723
Sun W, Han R, Xi H, Chen W, Li Y, Zhou Q, Li X, Huang K, Bontempo V, Gu X, et al. Effects of Alpha-Ketoglutarate Supplementation on Growth Performance, Diarrhea Incidence, Plasma Amino Acid, and Nutrient Digestibility in Weaned Piglets. Animals. 2025; 15(12):1723. https://doi.org/10.3390/ani15121723
Chicago/Turabian StyleSun, Weiyan, Ruyi Han, Hongbo Xi, Wenning Chen, Yanpin Li, Qingchao Zhou, Xilong Li, Kaikun Huang, Valentino Bontempo, Xu Gu, and et al. 2025. "Effects of Alpha-Ketoglutarate Supplementation on Growth Performance, Diarrhea Incidence, Plasma Amino Acid, and Nutrient Digestibility in Weaned Piglets" Animals 15, no. 12: 1723. https://doi.org/10.3390/ani15121723
APA StyleSun, W., Han, R., Xi, H., Chen, W., Li, Y., Zhou, Q., Li, X., Huang, K., Bontempo, V., Gu, X., & Jiang, X. (2025). Effects of Alpha-Ketoglutarate Supplementation on Growth Performance, Diarrhea Incidence, Plasma Amino Acid, and Nutrient Digestibility in Weaned Piglets. Animals, 15(12), 1723. https://doi.org/10.3390/ani15121723