
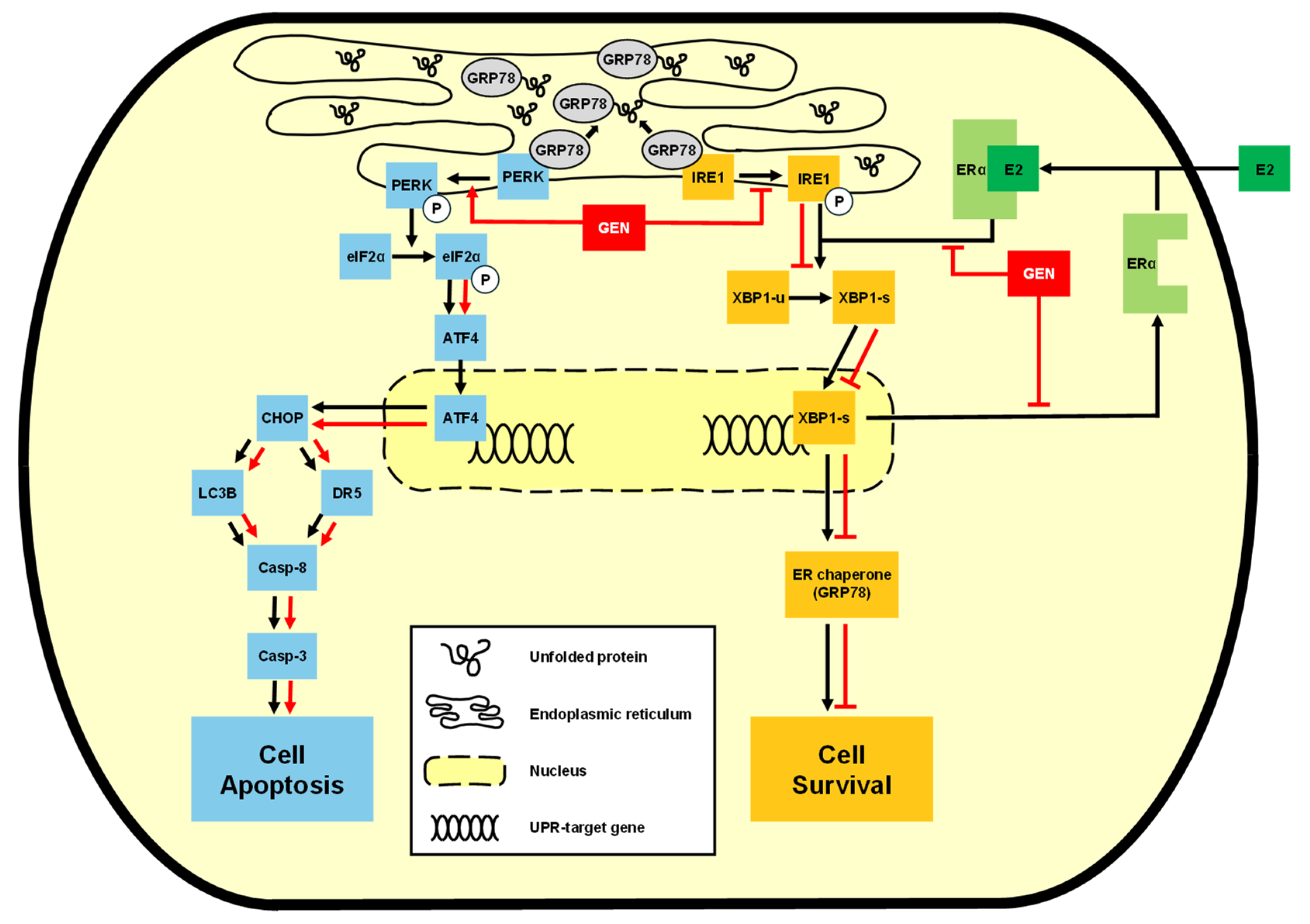
The Anticancer Effect of Genistein Through Enhancing PERK Signaling and Suppressing the IRE1α-XBP1 Axis in Canine Mammary Gland Tumor Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. 3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) Assay
2.3. Annexin v/Propidium Iodide (PI) Staining
2.4. Western Blotting
2.5. Immunocytochemistry
2.6. Statistical Analysis
3. Results
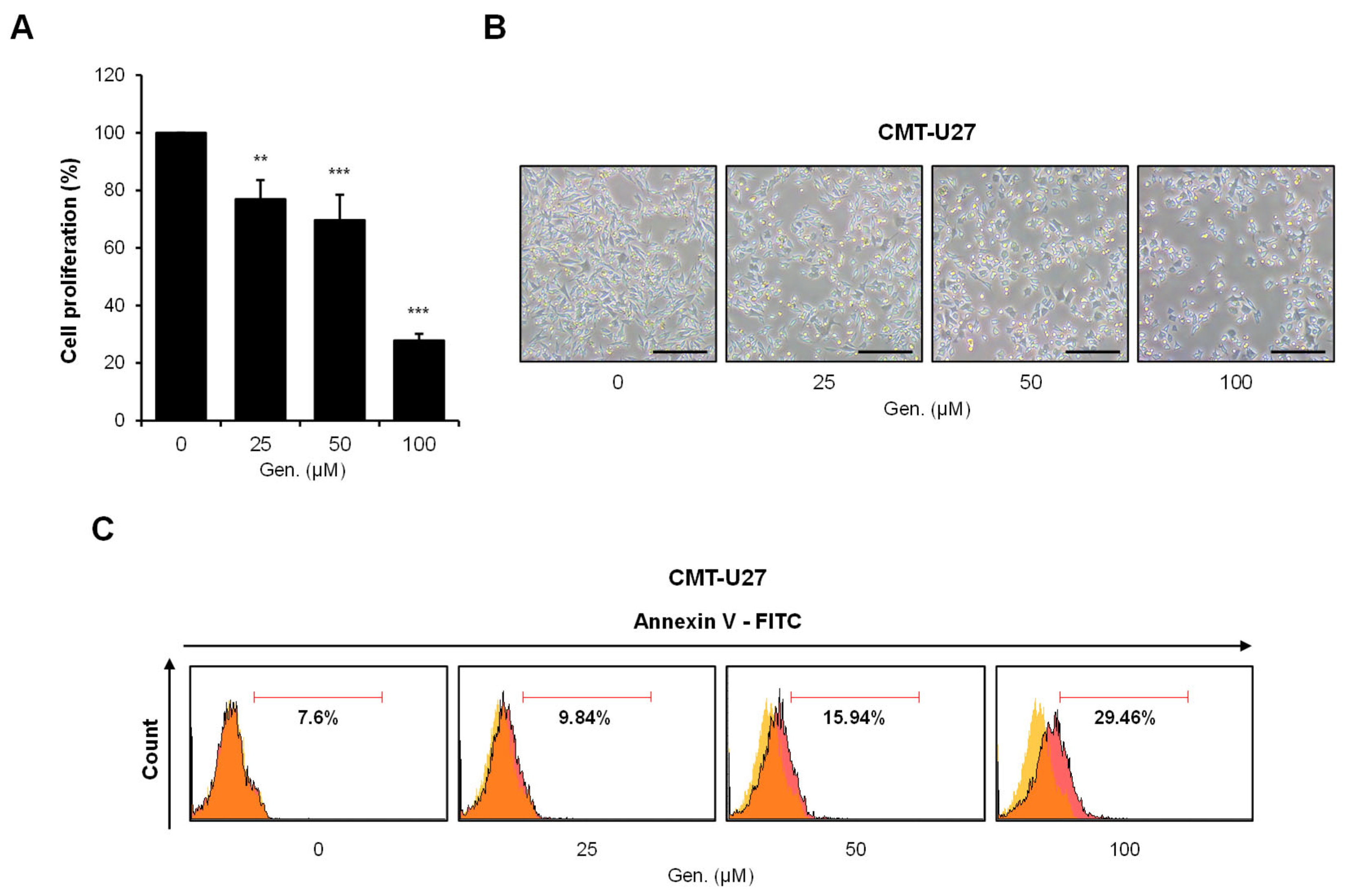
3.1. Genistein Inhibits Cell Proliferation and Induces Apoptosis in CMT-U27 Cells
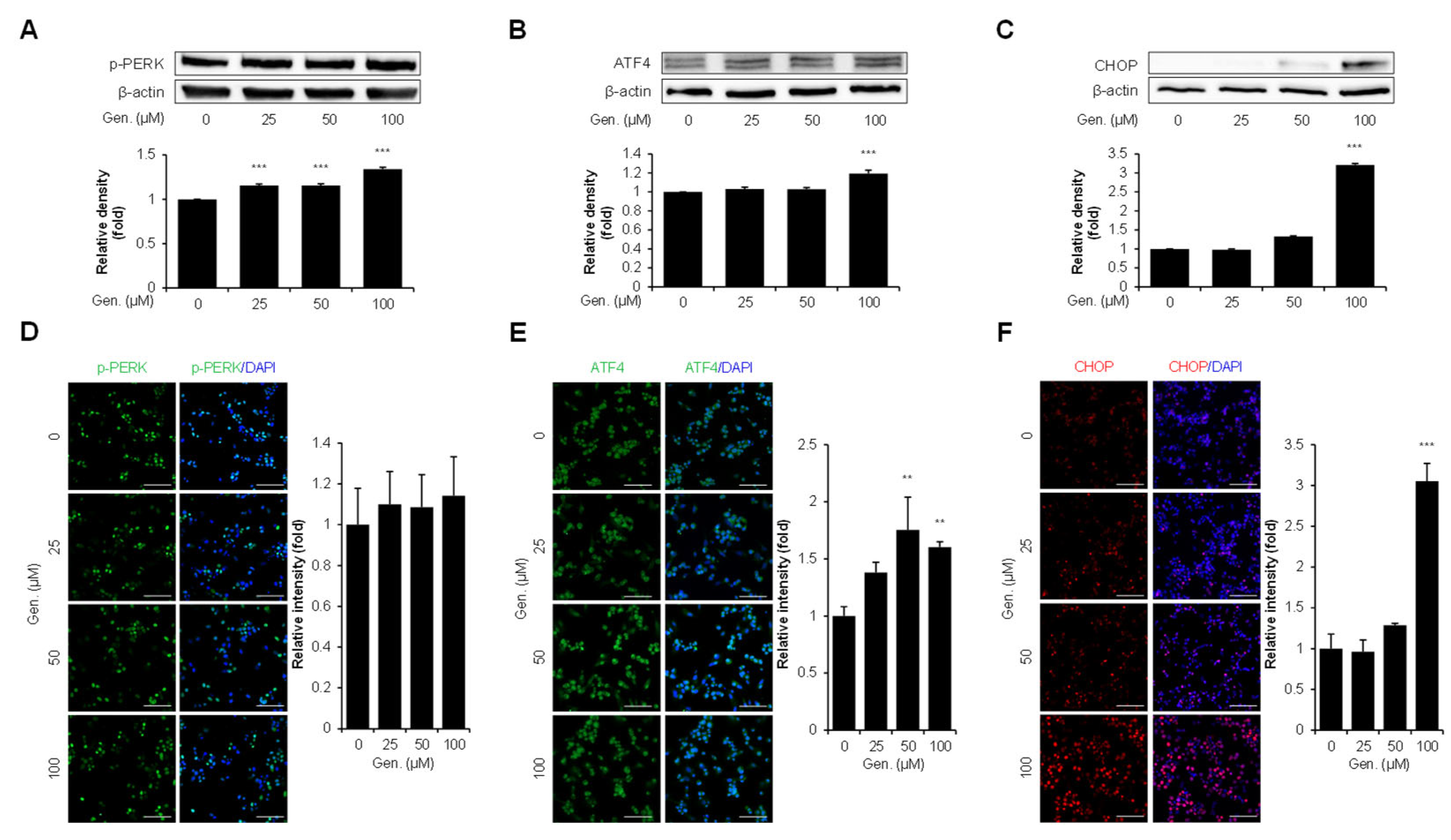
3.2. Genistein Activates the PERK–ATF4–CHOP Pathway in CMT-U27 Cells
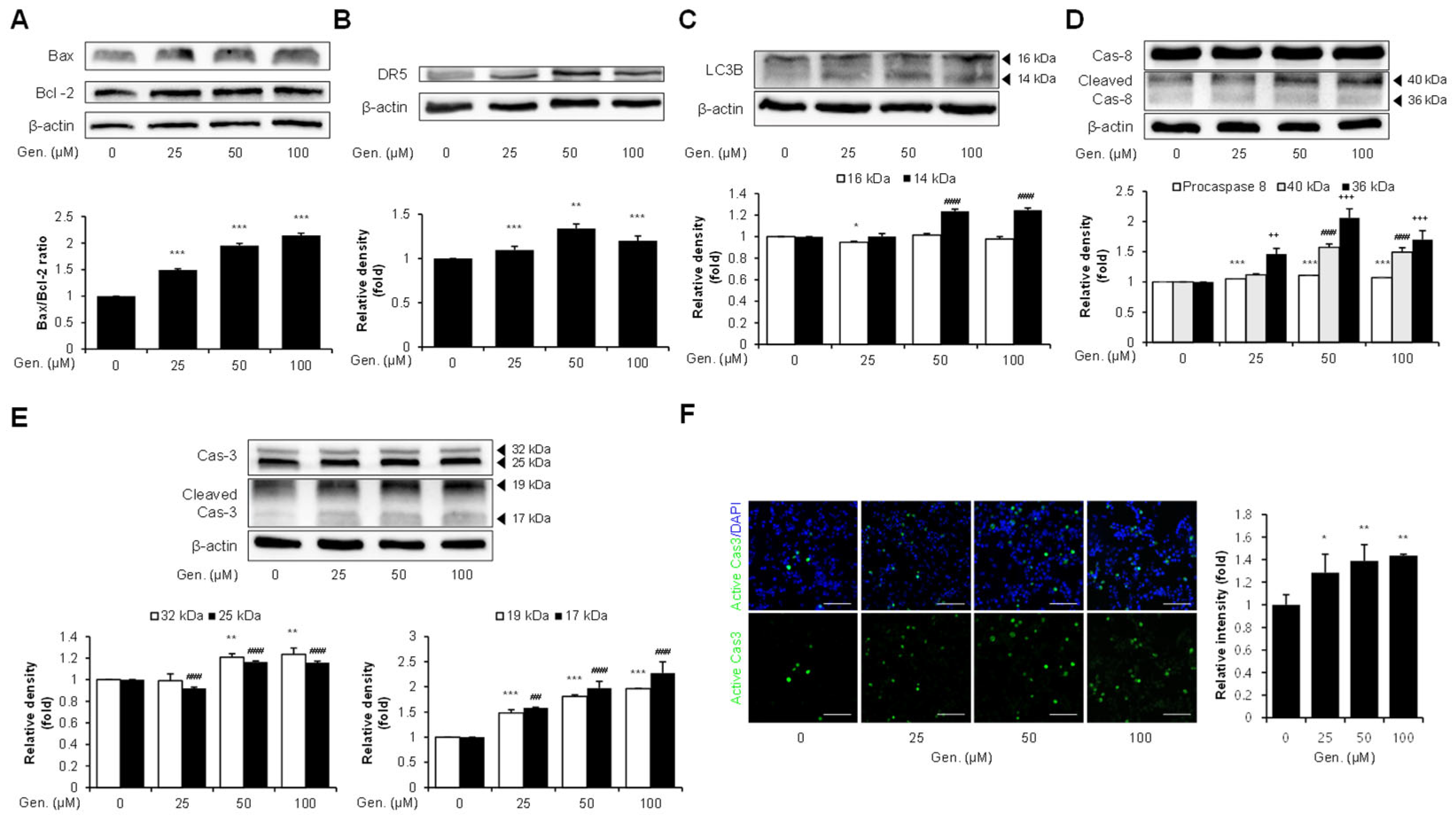
3.3. Genistein Promotes CHOP-Mediated Apoptosis via DR5, LC3B–Caspase-8–Caspase-3, and Bax/Bcl-2 Signaling in CMT-U27 Cells
3.4. Genistein Suppresses the IRE1α–XBP1s Pathway in CMT-U27 Cells
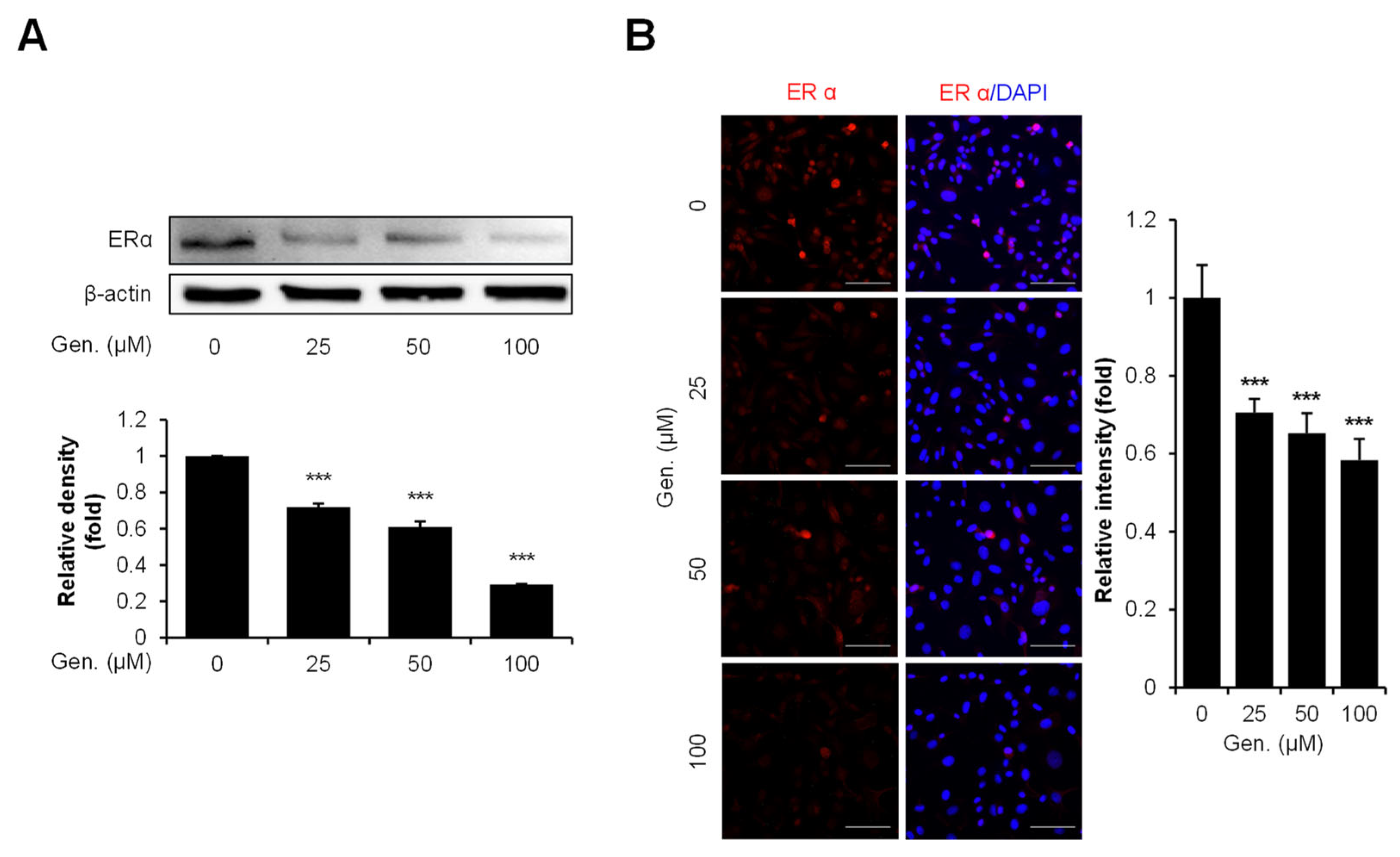
3.5. Genistein Downregulates ERα Expression in CMT-U27 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CMT | Canine mammary gland tumor |
| ER | Endoplasmic reticulum |
| PERK | Protein kinase-like ER kinase |
| IRE1α | Inositol-requiring enzyme 1 alpha |
| ATF6 | Activating transcription factor 6 |
| GRP78 | Glucose-regulated protein 78 |
| UPR | Unfolded protein response |
| XBP1 | X-box binding protein 1 |
| ESR1 | Estrogen receptor 1 |
| ATF4 | Activating transcription factor 4 |
| CHOP | C/EBP homologous protein |
| p-PERK | Phosphorylated PERK |
| p-IRE1α | Phosphorylated IRE1α |
| XBP1s | Spliced XBP1 |
| ERα | Estrogen receptor alpha |
| Bcl2 | B-cell lymphoma-2 |
| Bax | Bcl2-associated X |
| FBS | Fetal bovine serum |
| MTS | 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium |
| PI | Propidium Iodide |
| DR5 | Death receptor 5 |
| LC3B | Microtubule-associated protein 1 light chain 3 beta |
| HRP | Horse radish peroxidase |
| PBS | Phosphate-buffered saline |
| BSA | Bovine serum albumin |
| DAPI | 4′6-Diamidino-2-Phenylindole |
| SD | Standard deviation |
| Gen | Genistein |
References
- Sorenmo, K.U.; Kristiansen, V.M.; Cofone, M.A.; Shofer, F.S.; Breen, A.M.; Langeland, M.; Mongil, C.M.; Grondahl, A.M.; Teige, J.; Goldschmidt, M.H. Canine mammary gland tumours; a histological continuum from benign to malignant; clinical and histopathological evidence. Vet. Comp. Oncol. 2009, 7, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Brønden, L.B.; Nielsen, S.S.; Toft, N.; Kristensen, A.T. Data from the Danish veterinary cancer registry on the occurrence and distribution of neoplasms in dogs in Denmark. Vet. Rec. 2010, 166, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, G.; Alonso-Diez, Á.; Pérez-Alenza, D.; Peña, L. From conventional to precision therapy in canine mammary cancer: A comprehensive review. Front. Vet. Sci. 2021, 8, 623800. [Google Scholar] [CrossRef] [PubMed]
- Hörnfeldt, M.B.; Mortensen, J.K. Surgical dose and the clinical outcome in the treatment of mammary gland tumours in female dogs: A literature review. Acta Vet. Scand. 2023, 65, 12. [Google Scholar] [CrossRef]
- Machado, M.C.; Yamamoto, P.A.; Pippa, L.F.; de Moraes, N.V.; Neves, F.M.F.; Portela, R.D.; Barrouin-Melo, S.M.; Hielm-Björkman, A.; Godoy, A.; Estrela-Lima, A. Pharmacokinetics of Carboplatin in Combination with Low-Dose Cyclophosphamide in Female Dogs with Mammary Carcinoma. Animals 2022, 12, 3109. [Google Scholar] [CrossRef]
- Xu, E.; Hu, M.; Ge, R.; Tong, D.; Fan, Y.; Ren, X.; Liu, Y. LncRNA-42060 Regulates Tamoxifen Sensitivity and Tumor Development via Regulating the miR-204-5p/SOX4 Axis in Canine Mammary Gland Tumor Cells. Front. Vet. Sci. 2021, 8, 654694. [Google Scholar] [CrossRef]
- Zhou, C.; Lin, Z.; Li, X.; Zhang, D.; Song, P. Establishment and characterization of a multi-drug resistant cell line for canine mammary tumors. Front. Vet. Sci. 2023, 10, 1129756. [Google Scholar] [CrossRef]
- Petrucci, G.N.; Magalhães, T.R.; Dias, M.; Queiroga, F.L. Metronomic chemotherapy: Bridging theory to clinical application in canine and feline oncology. Front. Vet. Sci. 2024, 11, 1397376. [Google Scholar] [CrossRef]
- Sorenmo, K. Canine mammary gland tumors. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 573–596. [Google Scholar] [CrossRef]
- Hsiao, Y.L.; Hsieh, T.Z.; Liou, C.J.; Cheng, Y.H.; Lin, C.T.; Chang, C.Y.; Lai, Y.S. Characterization of protein marker expression, tumorigenicity, and doxorubicin chemoresistance in two new canine mammary tumor cell lines. BMC Vet. Res. 2014, 10, 229. [Google Scholar] [CrossRef]
- Yang, Y.; Mei, C.; Xian, H.; Zhang, X.; Li, J.; Liang, Z.X.; Zhi, Y.; Ma, Y.; Wang, H.J. Toosendanin-induced apoptosis of CMT-U27 is mediated through the mitochondrial apoptotic pathway. Vet. Comp. Oncol. 2023, 21, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Gavrilas, L.I.; Cruceriu, D.; Mocan, A.; Loghin, F.; Miere, D.; Balacescu, O. Plant-Derived Bioactive Compounds in Colorectal Cancer: Insights from Combined Regimens with Conventional Chemotherapy to Overcome Drug-Resistance. Biomedicines 2022, 10, 1948. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, A.M.; Meléndez, C.M.; Uribe, D.; Pedroza-Díaz, J. Synergistic effects of natural compounds and conventional chemotherapeutic agents: Recent insights for the development of cancer treatment strategies. Heliyon 2022, 8, e09519. [Google Scholar] [CrossRef]
- Mehraj, U.; Dar, A.H.; Wani, N.A.; Mir, M.A. Tumor microenvironment promotes breast cancer chemoresistance. Cancer Chemother. Pharmacol. 2021, 87, 147–158. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, Y.; Oyang, L.; Cui, S.; Li, S.; Li, J.; Liu, L.; Li, Y.; Peng, M.; Tan, S.; et al. Endoplasmic reticulum stress—A key guardian in cancer. Cell Death Discov. 2024, 10, 343. [Google Scholar] [CrossRef]
- McGrath, E.P.; Logue, S.E.; Mnich, K.; Deegan, S.; Jager, R.; Gorman, A.M.; Samali, A. The Unfolded Protein Response in Breast Cancer. Cancers 2018, 10, 344. [Google Scholar] [CrossRef]
- Dan, V.M.; Raveendran, R.S.; Baby, S. Resistance to intervention: Paclitaxel in breast cancer. Mini Rev. Med. Chem. 2021, 21, 1237–1268. [Google Scholar] [CrossRef]
- Mu, W.; Zhi, Y.; Zhou, J.; Wang, C.; Chai, K.; Fan, Z.; Lv, G. Endoplasmic reticulum stress and quality control in relation to cisplatin resistance in tumor cells. Front. Pharmacol. 2024, 15, 1419468. [Google Scholar] [CrossRef]
- Chen, S.; Chen, J.; Hua, X.; Sun, Y.; Cui, R.; Sha, J.; Zhu, X. The emerging role of XBP1 in cancer. Biomed. Pharmacother. 2020, 127, 110069. [Google Scholar] [CrossRef]
- Fan, C.F.; Mao, X.Y.; Wang, E.H. Elevated p-CREB-2 (ser 245) expression is potentially associated with carcinogenesis and development of breast carcinoma. Mol. Med. Rep. 2012, 5, 357–362. [Google Scholar] [CrossRef]
- Zheng, Y.Z.; Cao, Z.G.; Hu, X.; Shao, Z.M. The endoplasmic reticulum stress markers GRP78 and CHOP predict disease-free survival and responsiveness to chemotherapy in breast cancer. Breast Cancer Res. Treat. 2014, 145, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Shuda, M.; Kondoh, N.; Imazeki, N.; Tanaka, K.; Okada, T.; Mori, K.; Hada, A.; Arai, M.; Wakatsuki, T.; Matsubara, O.; et al. Activation of the ATF6, XBP1 and grp78 genes in human hepatocellular carcinoma: A possible involvement of the ER stress pathway in hepatocarcinogenesis. J. Hepatol. 2003, 38, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Rasheed, S.; Rehman, K.; Shahid, M.; Suhail, S.; Akash, M.S.H. Therapeutic potentials of genistein: New insights and perspectives. J. Food Biochem. 2022, 46, e14228. [Google Scholar] [CrossRef]
- Bhat, S.S.; Prasad, S.K.; Shivamallu, C.; Prasad, K.S.; Syed, A.; Reddy, P.; Cull, C.A.; Amachawadi, R.G. Genistein: A Potent Anti-Breast Cancer Agent. Curr. Issues Mol. Biol. 2021, 43, 1502–1517. [Google Scholar] [CrossRef]
- Pavese, J.M.; Krishna, S.N.; Bergan, R.C. Genistein inhibits human prostate cancer cell detachment, invasion, and metastasis. Am. J. Clin. Nutr. 2014, 100, 431S–436S. [Google Scholar] [CrossRef]
- Bi, Y.-L.; Min, M.; Shen, W.; Liu, Y. Genistein induced anticancer effects on pancreatic cancer cell lines involves mitochondrial apoptosis, G0/G1cell cycle arrest and regulation of STAT3 signalling pathway. Phytomedicine 2018, 39, 10–16. [Google Scholar] [CrossRef]
- Yeh, T.C.; Chiang, P.C.; Li, T.K.; Hsu, J.L.; Lin, C.J.; Wang, S.W.; Peng, C.Y.; Guh, J.H. Genistein induces apoptosis in human hepatocellular carcinomas via interaction of endoplasmic reticulum stress and mitochondrial insult. Biochem. Pharmacol. 2007, 73, 782–792. [Google Scholar] [CrossRef]
- Naponelli, V.; Piscazzi, A.; Mangieri, D. Cellular and Molecular Mechanisms Modulated by Genistein in Cancer. Int. J. Mol. Sci. 2025, 26, 1114. [Google Scholar] [CrossRef]
- Siwecka, N.; Rozpędek-Kamińska, W.; Wawrzynkiewicz, A.; Pytel, D.; Diehl, J.A.; Majsterek, I. The Structure, Activation and Signaling of IRE1 and Its Role in Determining Cell Fate. Biomedicines 2021, 9, 156. [Google Scholar] [CrossRef]
- Hetz, C.; Papa, F.R. The Unfolded Protein Response and Cell Fate Control. Mol. Cell 2018, 69, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef] [PubMed]
- Lindner, P.; Christensen, S.B.; Nissen, P.; Møller, J.V.; Engedal, N. Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components. Cell Commun. Signal. 2020, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Mohan, N.; Chakrabarti, M.; Banik, N.L.; Ray, S.K. Combination of LC3 shRNA plasmid transfection and genistein treatment inhibited autophagy and increased apoptosis in malignant neuroblastoma in cell culture and animal models. PLoS ONE 2013, 8, e78958. [Google Scholar] [CrossRef]
- Barua, D.; Gupta, A.; Gupta, S. Targeting the IRE1-XBP1 axis to overcome endocrine resistance in breast cancer: Opportunities and challenges. Cancer Lett. 2020, 486, 29–37. [Google Scholar] [CrossRef]
- Chapple, R.H.; Hu, T.; Tseng, Y.-J.; Liu, L.; Kitano, A.; Luu, V.; Hoegenauer, K.A.; Iwawaki, T.; Li, Q.; Nakada, D. ERα promotes murine hematopoietic regeneration through the Ire1α-mediated unfolded protein response. eLife 2018, 7, e31159. [Google Scholar] [CrossRef]
- Boudreau, M.W.; Duraki, D.; Wang, L.; Mao, C.; Kim, J.E.; Henn, M.A.; Tang, B.; Fanning, S.W.; Kiefer, J.; Tarasow, T.M.; et al. A small-molecule activator of the unfolded protein response eradicates human breast tumors in mice. Sci. Transl. Med. 2021, 13, eabf1383. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, Y.-J.; Yoo, M.-J.; Jang, H.; Song, J.; Park, S.-Y.; Choi, J.; Seol, J.-W. The Anticancer Effect of Genistein Through Enhancing PERK Signaling and Suppressing the IRE1α-XBP1 Axis in Canine Mammary Gland Tumor Cells. Animals 2025, 15, 1717. https://doi.org/10.3390/ani15121717
Jang Y-J, Yoo M-J, Jang H, Song J, Park S-Y, Choi J, Seol J-W. The Anticancer Effect of Genistein Through Enhancing PERK Signaling and Suppressing the IRE1α-XBP1 Axis in Canine Mammary Gland Tumor Cells. Animals. 2025; 15(12):1717. https://doi.org/10.3390/ani15121717
Chicago/Turabian StyleJang, Ye-Ji, Min-Jae Yoo, Hyuk Jang, Jun Song, Sang-Youel Park, Jawun Choi, and Jae-Won Seol. 2025. "The Anticancer Effect of Genistein Through Enhancing PERK Signaling and Suppressing the IRE1α-XBP1 Axis in Canine Mammary Gland Tumor Cells" Animals 15, no. 12: 1717. https://doi.org/10.3390/ani15121717
APA StyleJang, Y.-J., Yoo, M.-J., Jang, H., Song, J., Park, S.-Y., Choi, J., & Seol, J.-W. (2025). The Anticancer Effect of Genistein Through Enhancing PERK Signaling and Suppressing the IRE1α-XBP1 Axis in Canine Mammary Gland Tumor Cells. Animals, 15(12), 1717. https://doi.org/10.3390/ani15121717