
Lactobacillus Supplementation Modulates Rumen Microbiota and Metabolism in Yaks Under Fattening Feeding Conditions: A Comprehensive Multi-Omics Analysis

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Models and Experimental Design
2.2. Sample Collection and Measurement
2.3. Statistical Analysis
3. Results
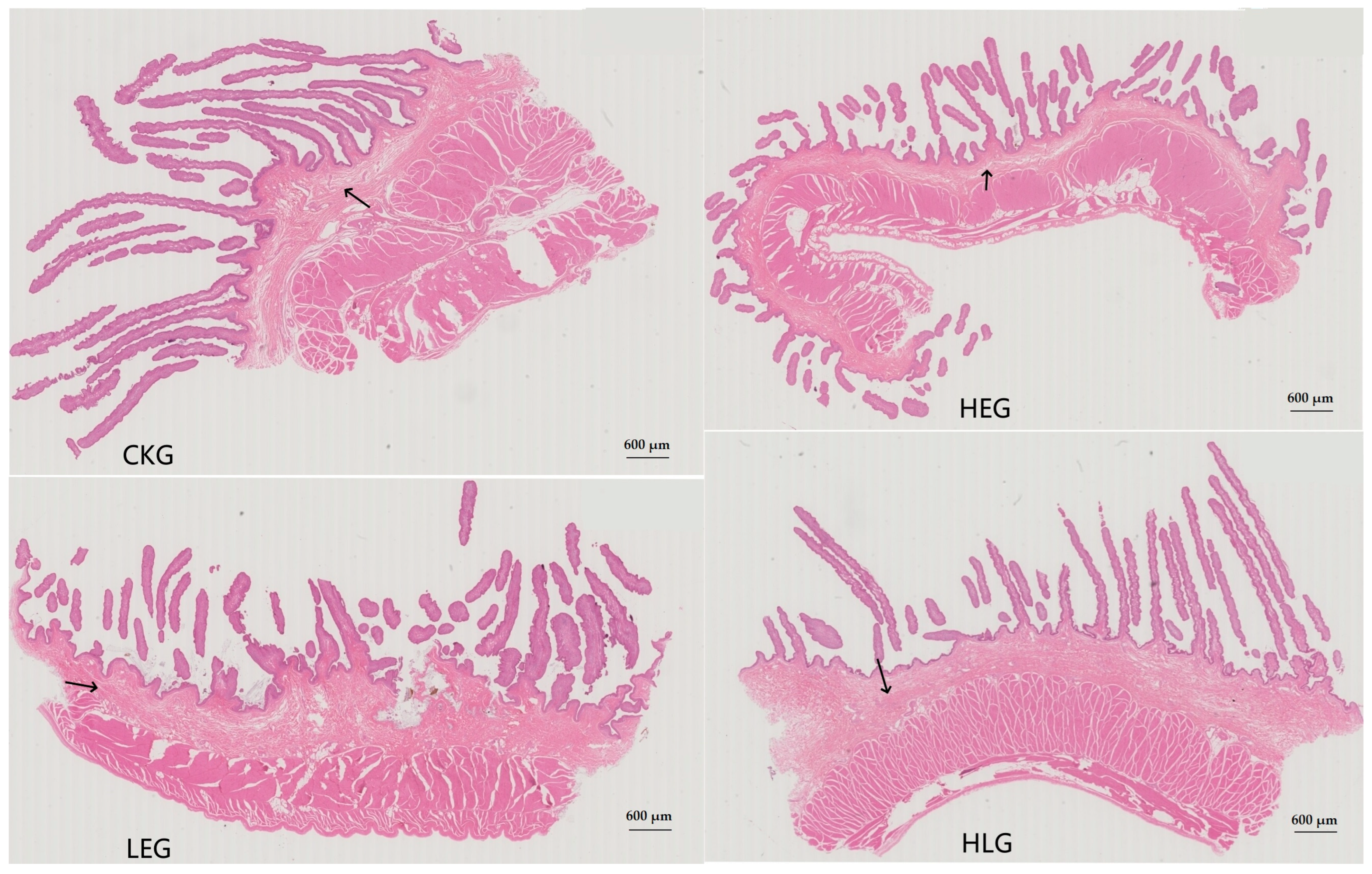
3.1. Changes in Yak Weight Is Linked to the Modulation of HED-Induced Changes by Lactobacillus in the Rumen Structure and Functional Homeostasis
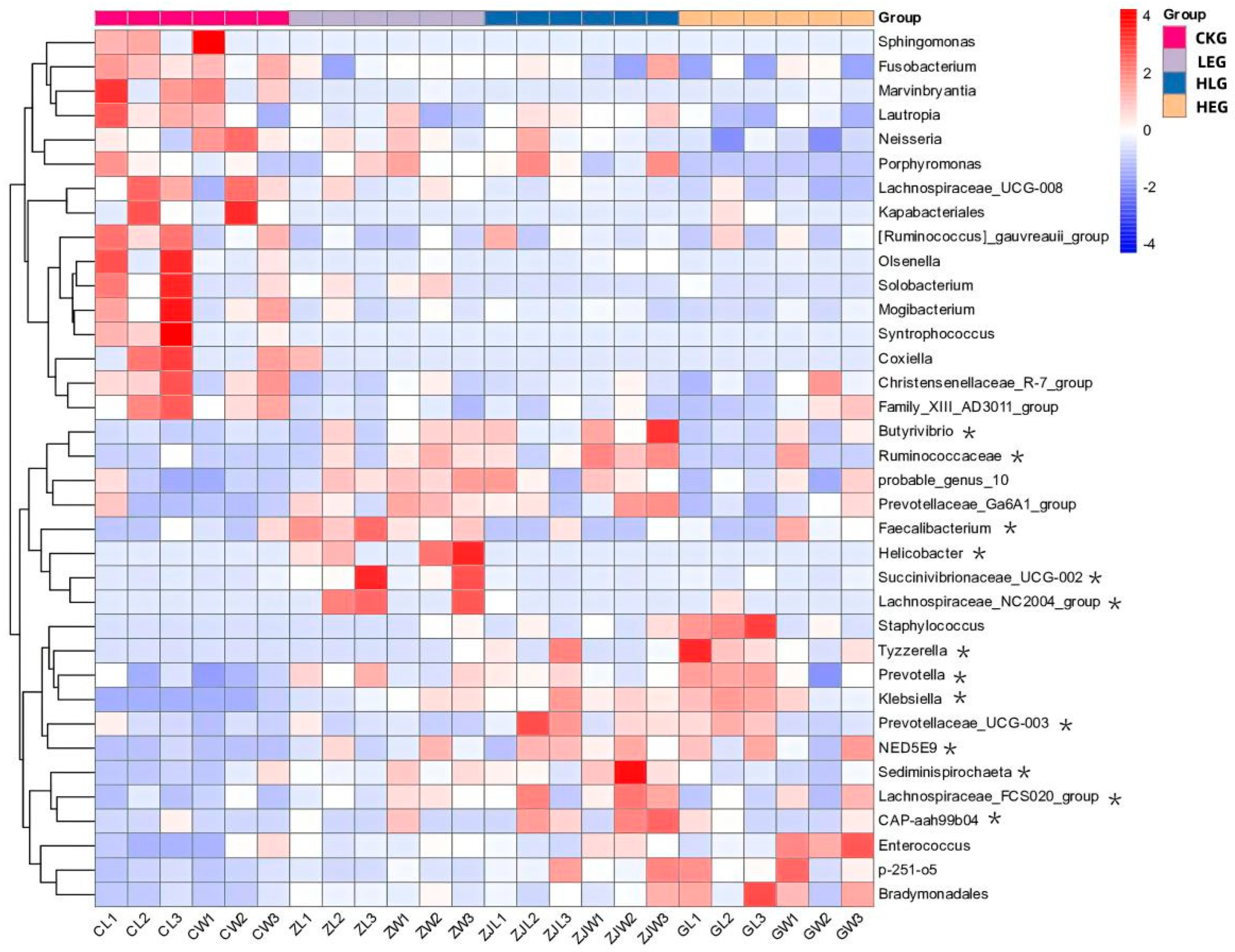
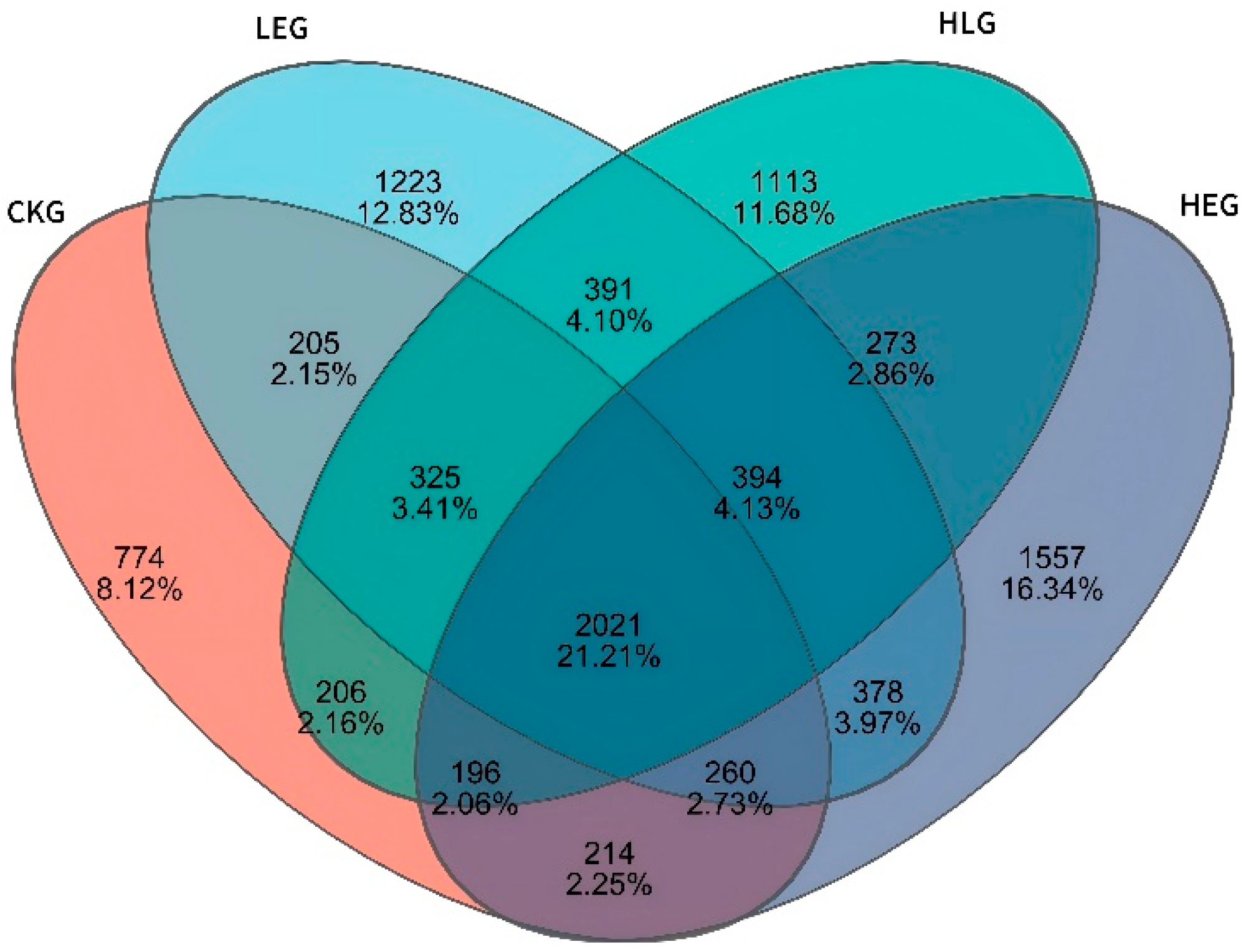
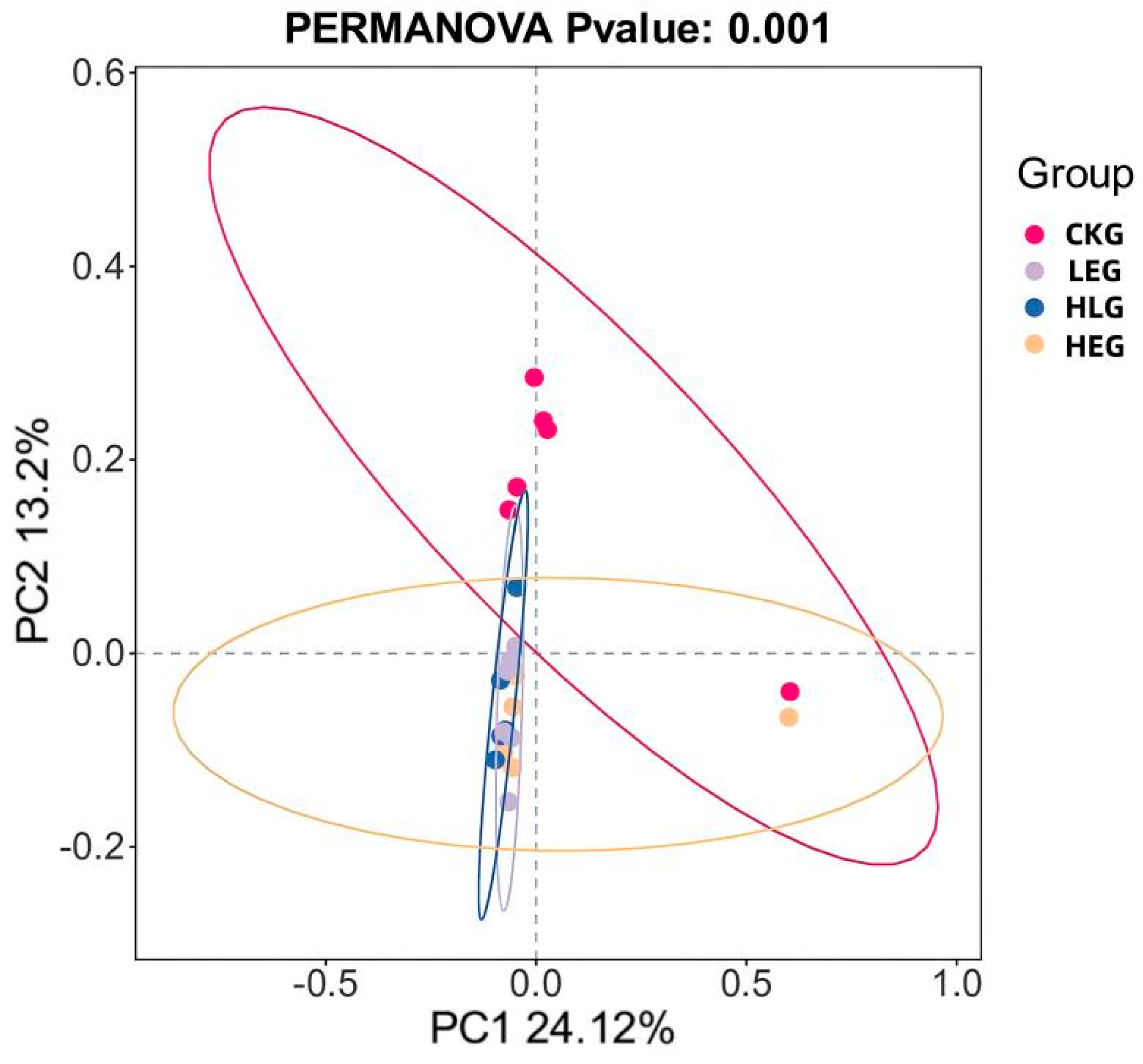
3.2. HED and Lactobacillus Intervention Are Linked to Compositional Rumen Microbiota Changes
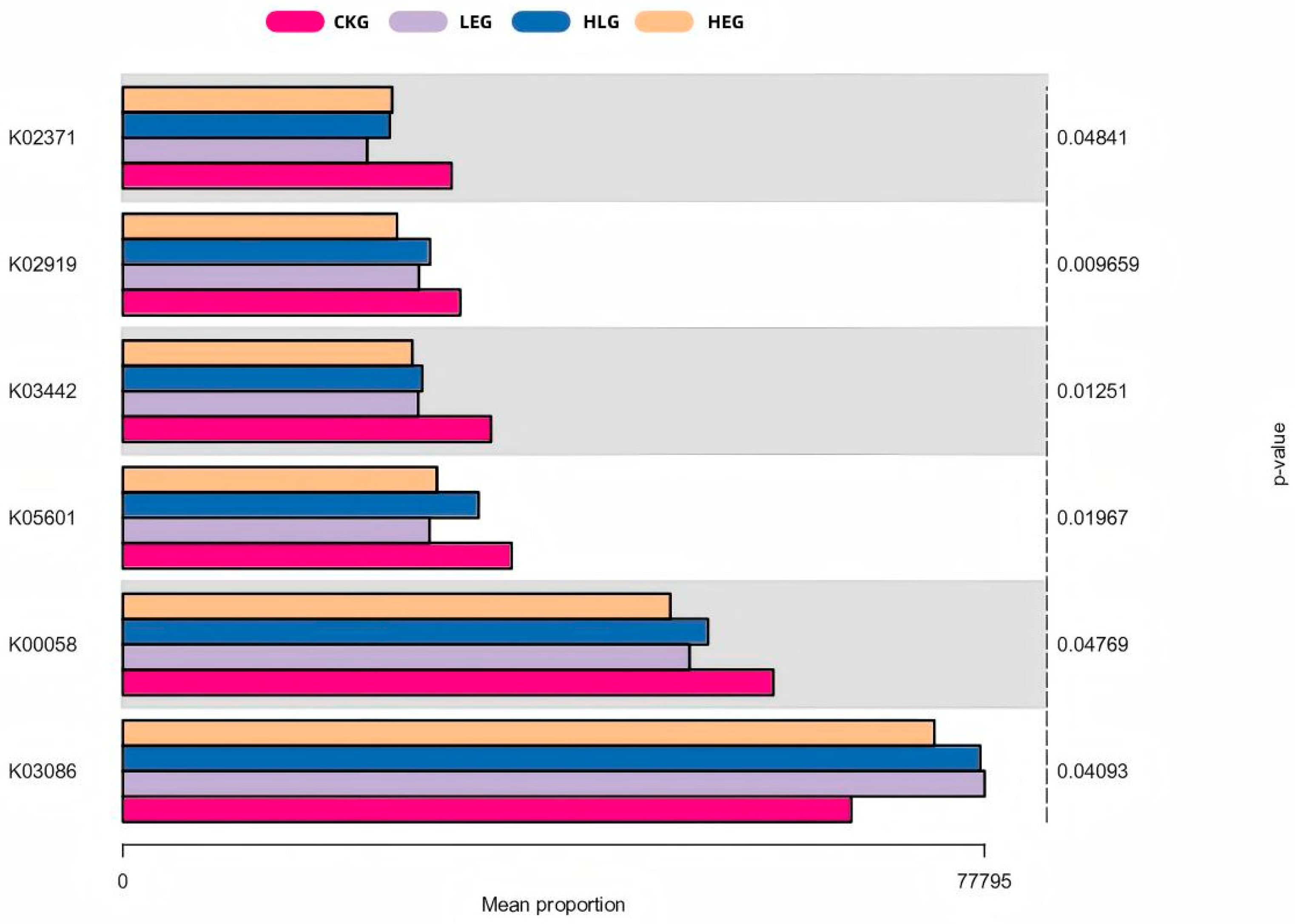
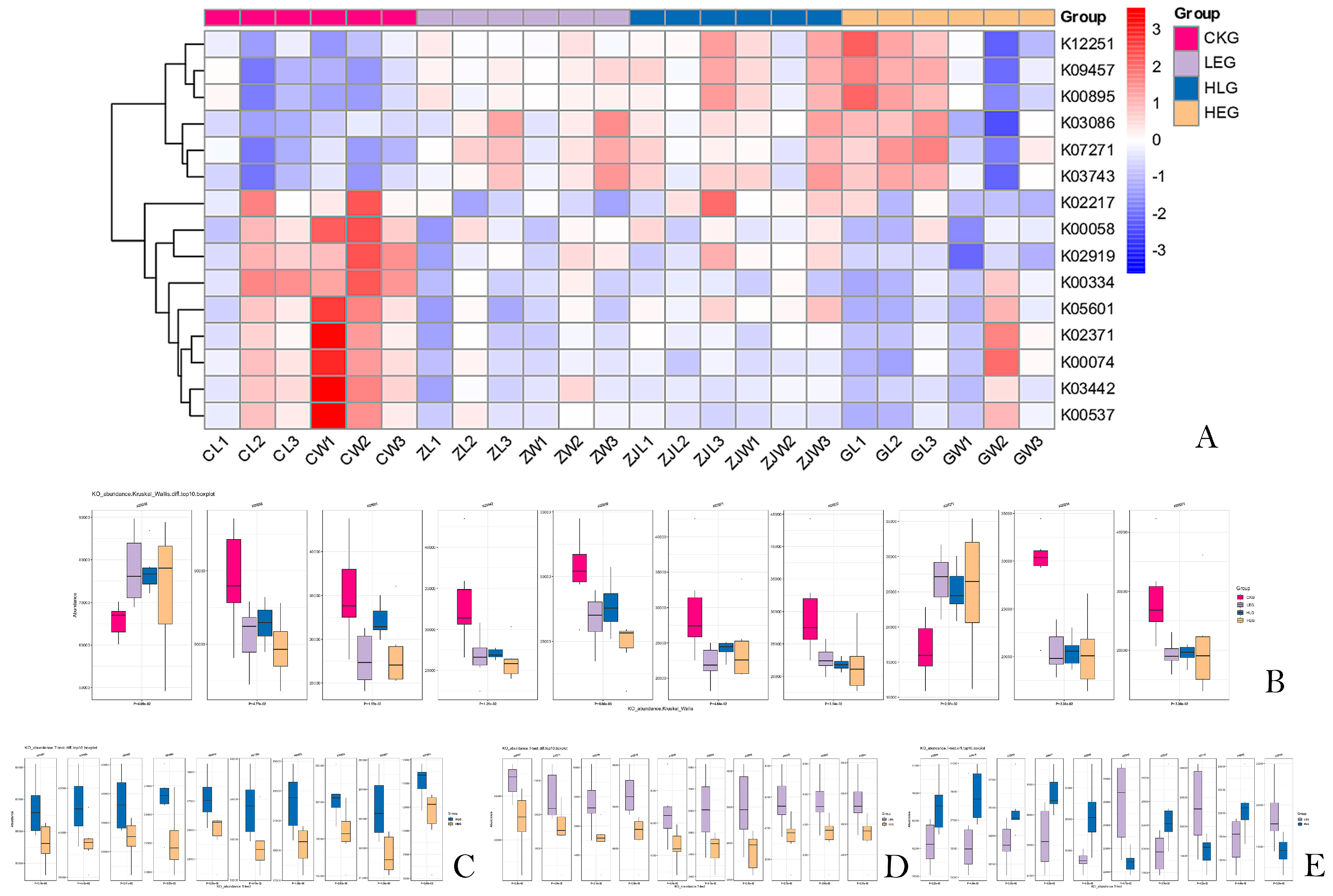
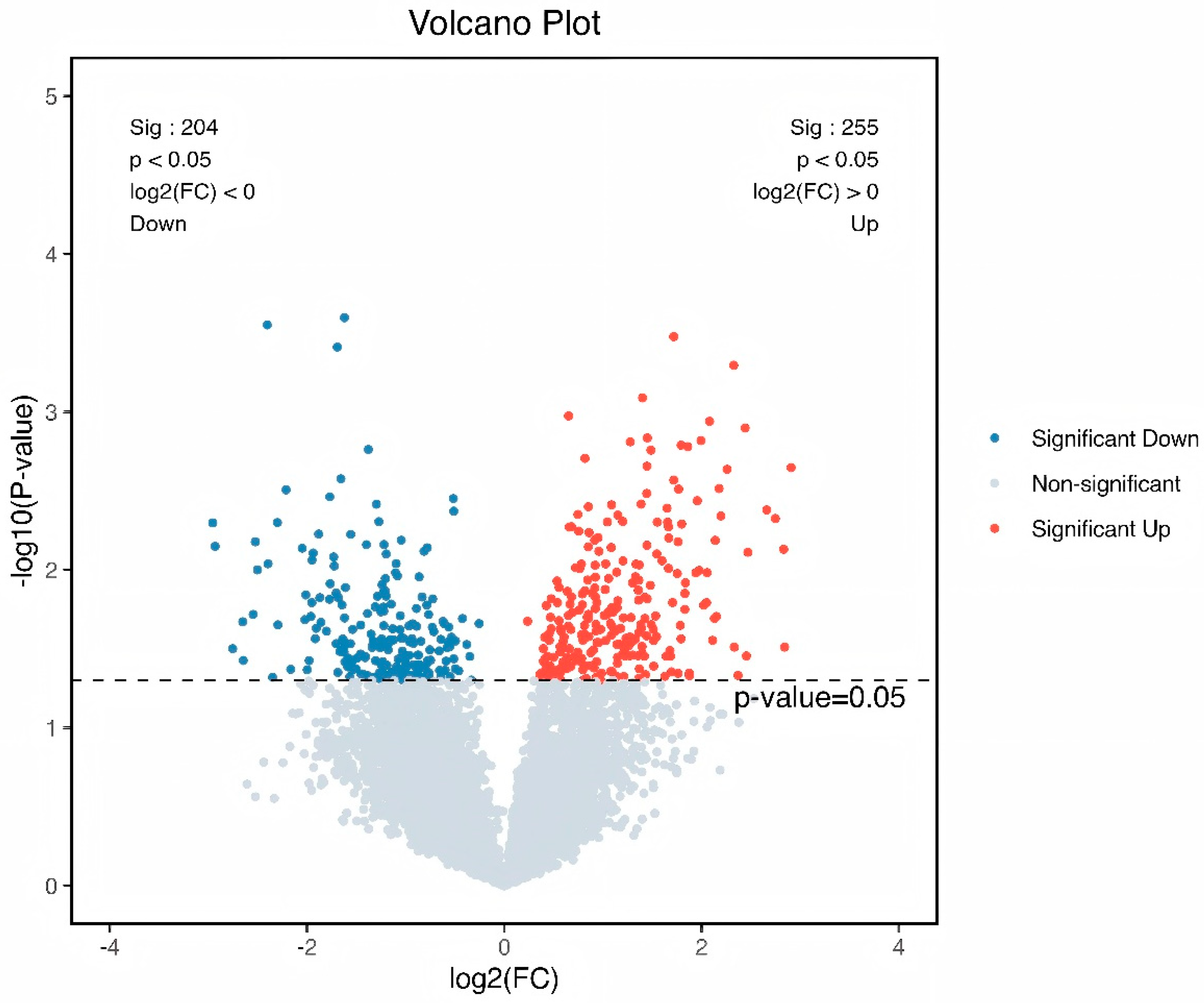
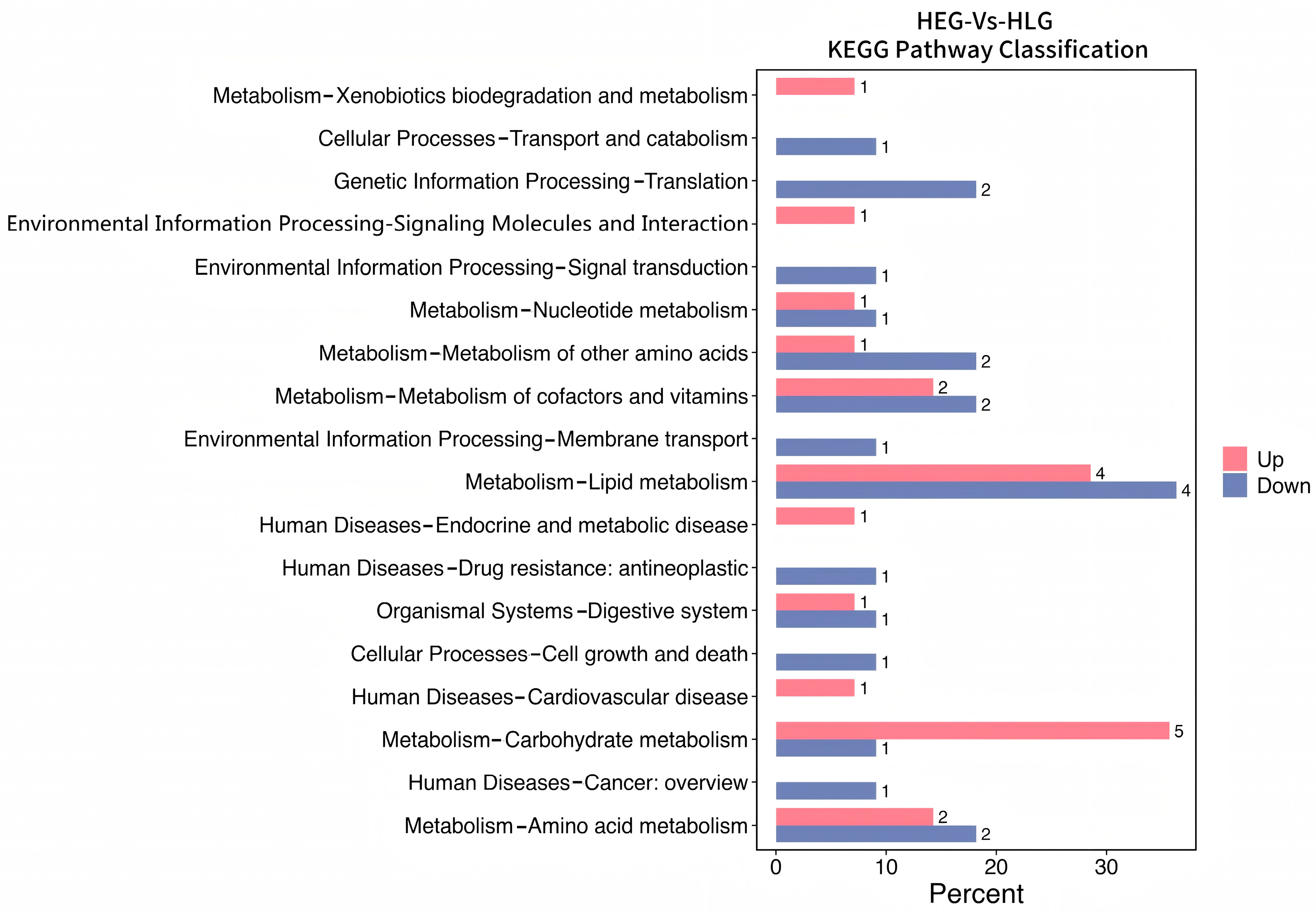
3.3. Identification of HED and Lactobacillus Intervention Treatment Markers by Demonstrating Variations in Kyoto Encyclopedia of Genes and Genomes (KEGG) Orthology (KO)
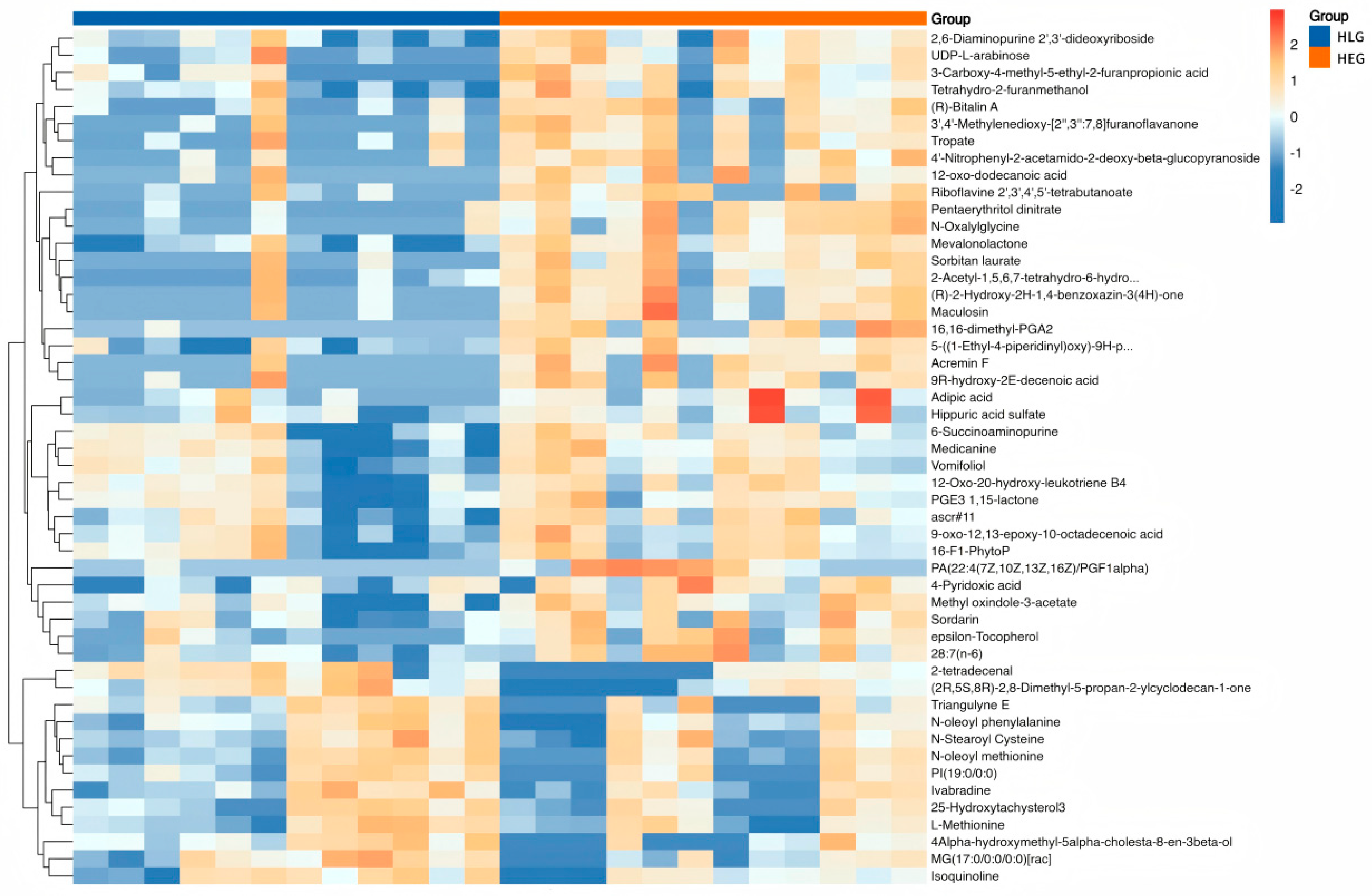
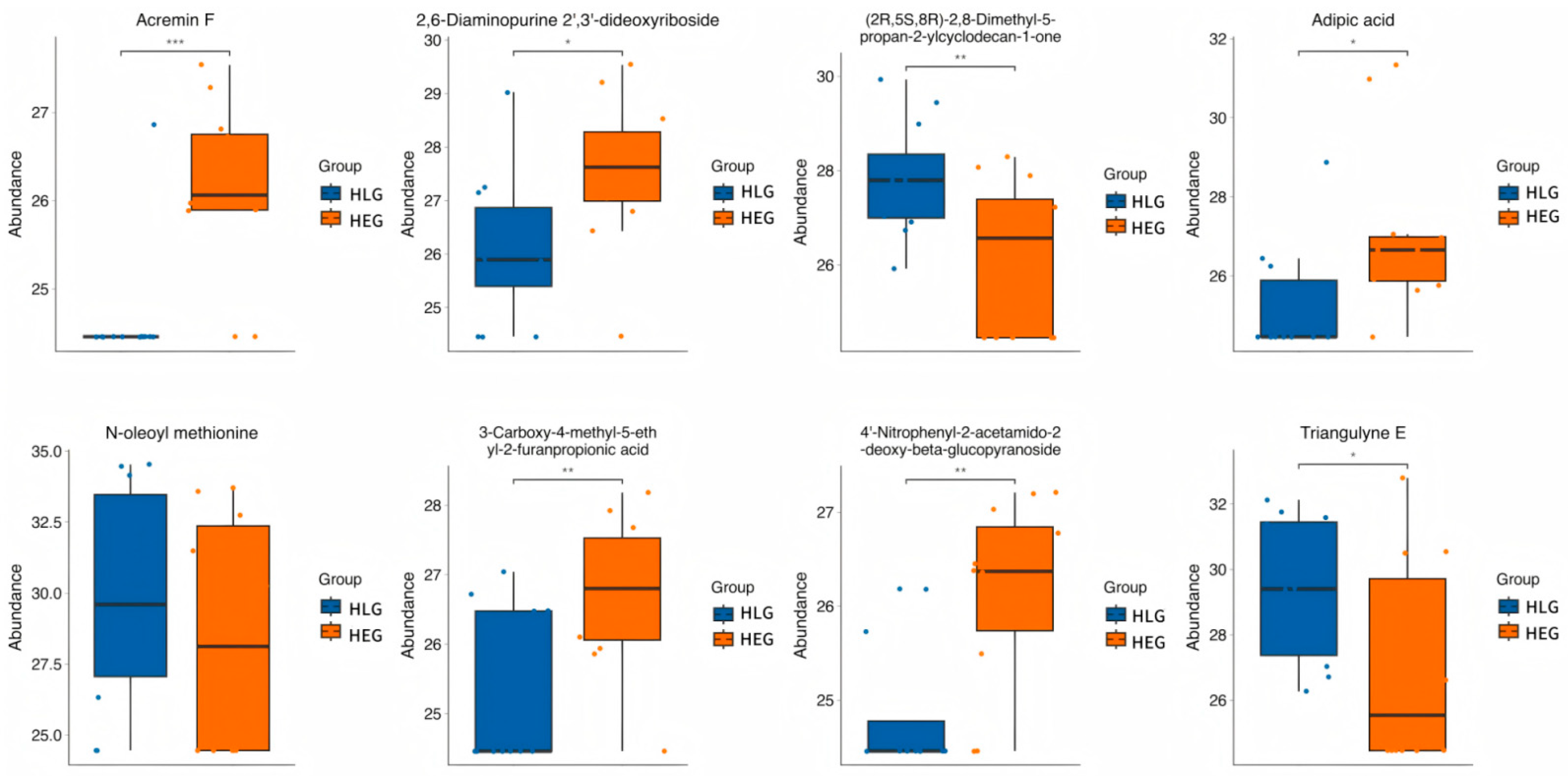
3.4. Demonstration of Metabolomic Alterations Across Different Lactobacillus Supplementation Strategies and Utilization of Signature Metabolites for HED-Mediated Rumen Barrier Dysfunction
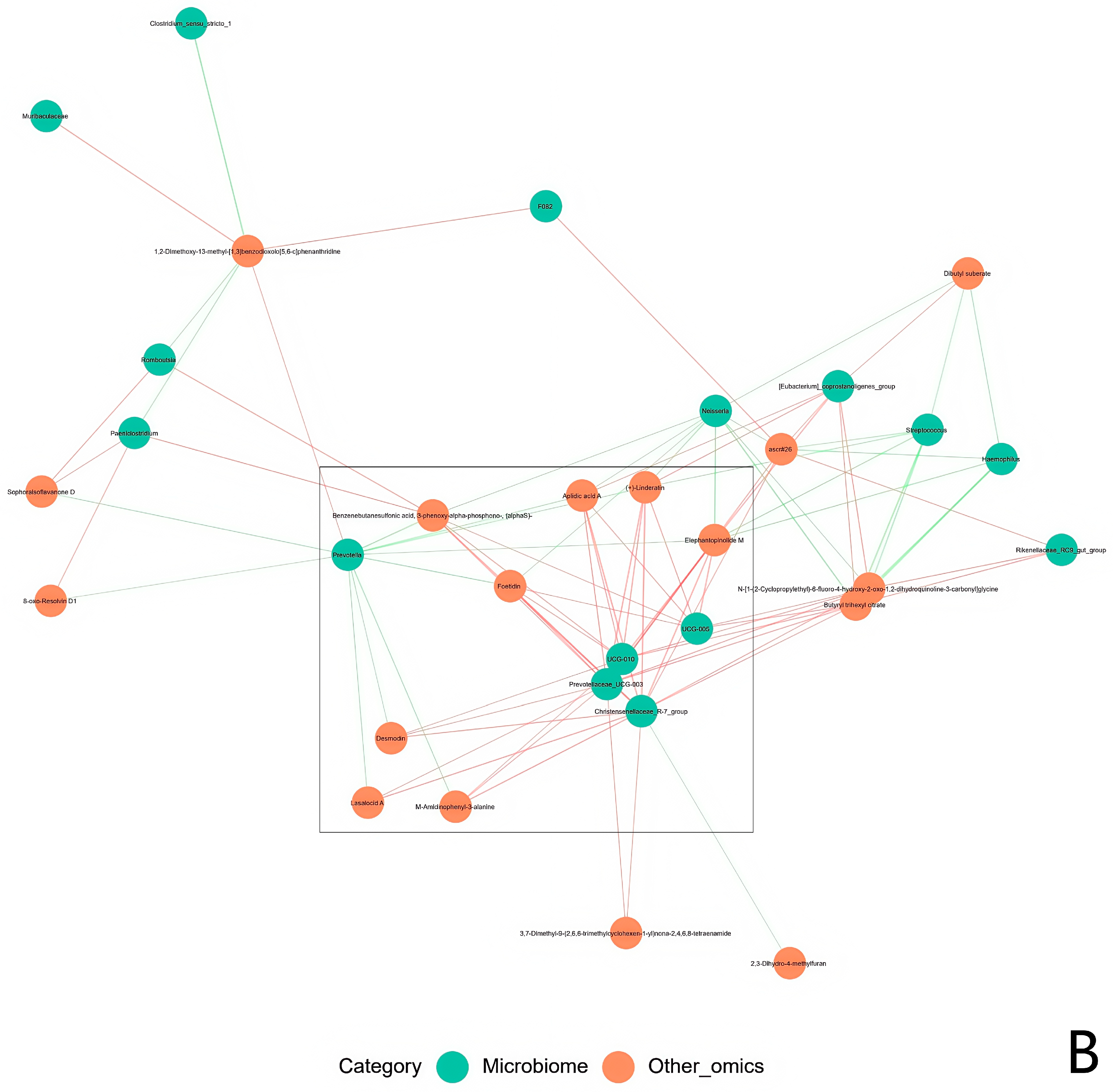
3.5. Multi-Omics Signature Integration for HED and Lactobacillus Intervention Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jia, J.L.; Liang, C.N.; Wu, X.Y.; Xiong, L.; Bao, P.J.; Chen, Q.; Yan, P. Effect of high proportion concentrate dietary on Ashdan Yak jejunal barrier and microbial function in cold season. Res. Vet. Sci. 2021, 140, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Youdol, T.; Mishra, A.K.; Raghuwanshi, M.S.; Ram, H.; Bhattacharjee, S.; Apurva, A.; Saxena, A. Availability and seasonal dynamics of feed sources for Ladakhi cattle breed in the cold arid region of Ladakh and future scope under increasing growing degree days. Biol. Rhythm Res. 2024, 55, 310–323. [Google Scholar] [CrossRef]
- Du, H.D.; Li, K.N.; Guo, W.L.; Na, M.L.; Zhang, J.; Na, R.H. Roughage Sources During Late Gestation and Lactation Alter Metabolism, Immune Function and Rumen Microbiota in Ewes and Their Offsprings. Microorganisms 2025, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.M.; Bano, I.; Qazi, I.H.; Matra, M.; Wanapat, M. “The Yak”—A remarkable animal living in a harsh environment: An overview of its feeding, growth, production performance, and contribution to food security. Front. Vet. Sci. 2023, 10, 1086985. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, S.; Acharya, K.P.; Laven, R.; Acharya, N. Possible consequences of climate change on survival, productivity and reproductive performance, and welfare of Himalayan Yak (Bos grunniens). Vet. Sci. 2022, 9, 449. [Google Scholar] [CrossRef]
- Jing, X.P.; Ding, L.M.; Zhou, J.W.; Huang, X.D.; Degen, A.; Long, R.J. The adaptive strategies of yaks to live in the Asian highlands. Anim. Nutr. 2022, 9, 249–258. [Google Scholar] [CrossRef]
- Zhang, L.; Xia, Z.H.; Fu, J.C.; Yang, Y. Role of the Rumen Epithelium and Associated Changes Under Zealand. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2022. [Google Scholar]
- Amat, S.; Dahlen, C.R.; Swanson, K.C.; Ward, A.K.; Reynolds, L.P.; Caton, J.S. Bovine animal model for studying the maternal microbiome, in utero microbial colonization and their role in offspring development and fetal programming. Front. Microbiol. 2022, 13, 854453. [Google Scholar] [CrossRef]
- Raza, B.; Zheng, Z.; Yang, W. A review on biofloc system technology, history, types, and future economical perceptions in aquaculture. Animals 2024, 14, 1489. [Google Scholar] [CrossRef]
- Deng, X.F.; Li, H.; Wu, A.M.; He, J.; Mao, X.B.; Dai, Z.L.; Tian, G.; Cai, J.Y.; Tang, J.Y.; Luo, Y.H. Composition, Influencing Factors, and Effects on Host Nutrient Metabolism of Fungi in Gastrointestinal Tract of Monogastric Animals. Animals 2025, 15, 710. [Google Scholar] [CrossRef]
- Fu, L.; Liu, L.; Zhang, L.; Hu, Y.H.; Zeng, Y.; Ran, Q.F.; Zhou, Y.; Zhou, P.; Chen, J.C.; Loor, J.; et al. Inoculation of Newborn Lambs with Ruminal Solids Derived from Adult Goats Reprograms the Development of Gut Microbiota and Serum Metabolome and Favors Growth Performance. J. Agric. Food Chem. 2024, 72, 983–998. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Jiang, X.Z.; Yu, Y.; Cui, Y.M.; Wang, W.; Luo, H.L.; Stergiadis, S.; Wang, B. Rumen microbiome-driven insight into bile acid metabolism host metabolic regulation. ISME J. 2024, 18, wrae098. [Google Scholar] [CrossRef] [PubMed]
- Rekhaa, K.; Venkidasamy, B.; Samynathan, R.; Nagella, P.; Rebezov, M.; Khayrullin, M.; Ponomarev, E.; Bouyahya, A.; Sarkar, T.; Shariati, M.A.; et al. Short-chain fatty acid: An updated review on signaling, metabolism, and therapeutic effects. Crit. Rev. Food Sci. Nutr. 2024, 64, 2461–2489. [Google Scholar] [CrossRef] [PubMed]
- Gavriel, M.T. The Effect of Obesity and Weight Loss on the Feline Fecal Microbiota, Inflammation and Intestinal Health Markers. Ph.D. Thesis, The University of Guelph, Guelph, ON, Canada, 2018. [Google Scholar]
- Salsinha, A.S.; Cima, A.; Araújo-Rodrigues, H.; Viana, S.; Reis, F.; Coscueta, E.R.; Rodríguez-Alcalá, L.M.; Relvas, J.B.; Pintado, M. The use of an in vitro fecal fermentation model to uncover the beneficial role of omega-3 and punicic acid in gut microbiota alterations induced by a Western diet. Food Funct. 2024, 15, 6095–6117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, L.J.; Hou, S.Z.; Raza, S.H.A.; Gui, L.S.; Sun, S.N.; Wang, Z.Y.; Yang, B.C.; Yuan, Z.Z.; Simal-Gandara, J.; et al. Metabolomics approach reveals high energy diet improves the quality and enhances the flavor of black Tibetan sheep meat by altering the composition of rumen microbiota. Front. Nutr. 2022, 9, 915558. [Google Scholar] [CrossRef]
- Wang, R.Z.; Bai, B.Q.; Huang, Y.Y.; Degen, A.; Mi, J.D.; Xue, Y.F.; Hao, L.Z. Yaks are dependent on gut microbiota for survival in the environment of the Qinghai Tibet plateau. Microorganisms 2024, 12, 1122. [Google Scholar] [CrossRef]
- Barathan, M.; Ng, S.L.; Lokanathan, Y.; Ng, M.H.; Law, J.X. The profound influence of gut microbiome and extracellular vesicles on animal health and disease. Int. J. Mol. Sci. 2024, 25, 4024. [Google Scholar] [CrossRef]
- Moretti, A.F.; Brizuela, N.S.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E.; Golowczyc, M.A. Current applications and future trends of dehydrated lactic acid bacteria for incorporation in animal feed products. Fermentation 2023, 9, 742. [Google Scholar] [CrossRef]
- Doyle, N.; Mbandlwa, P.; Kelly, W.J.; Attwood, G.; Li, Y.; Ross, R.P.; Stanton, C.; Leahy, S. Use of Lactic Acid Bacteria to Reduce Methane Production in Ruminants, a Critical Review. Front. Microbiol. 2019, 10, 2207. [Google Scholar] [CrossRef]
- Ogunade, I.; Schweickart, H.; McCoun, M.; Cannon, K.; McManus, C. Integrating 16S rRNA Sequencing and LC–MS-Based Metabolomics to Evaluate the Effects of Live Yeast on Rumen Function in Beef Cattle. Animals 2019, 9, 28. [Google Scholar] [CrossRef]
- Rachwał, K.; Gustaw, K. Lactic Acid Bacteria in Sustainable Food Production. Sustainability 2024, 16, 3362. [Google Scholar] [CrossRef]
- Marcari, A.B.; Paiva, A.D.; Simon, C.R.; dos Santos, M.E.S.M. Leaky Gut Syndrome: An Interplay Between Nutrients and Dysbiosis. Curr. Nutr. Rep. 2025, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.I.; Mahmood, S.; Hassan, M.; Sajid, M.; Ahmed, I.; Shokrollahi, B.; Shahzad, A.H.; Abbas, S.; Raza, S.; Khan, K.; et al. Genomic insights into Yak (Bos grunniens) adaptations for nutrient assimilation in high-altitudes. Sci. Rep. 2024, 14, 5650. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.L.; Liang, C.N.; Wu, X.Y.; Xiong, L.; Bao, P.J.; Chen, Q.; Yan, P. Effect of high proportion concentrate dietary on Yak jejunal structure, physiological function and protein composition during cold season. Sci. Rep. 2021, 11, 5502. [Google Scholar] [CrossRef]
- NY/T 815-2004; Feeding Standard of Beef Cattle. National Standards of the People’s Republic of China: Beijing, China, 2004.
- Yang, S. Feed Analysis and Quality Test Technology; Beijing Agriculture Press: Beijing, China, 1993. [Google Scholar]
- Nutrient Requirements of Beef Cattle: Eighth Revised Edition; The National Academies Press: Washington, DC, USA, 2014. [CrossRef]
- Ayalew, W.; Chu, M.; Liang, C.N.; Wu, X.Y.; Yan, P. Adaptation mechanisms of yak (Bos grunniens) to high-altitude environmental stress. Animals. 2021, 11, 2344. [Google Scholar] [CrossRef]
- Liu, S.M.; Yang, Y.Y.; Luo, H.L.; Pang, W.J.; Martin, G.B. Fat deposition and partitioning for meat production in cattle and sheep. Anim. Nutr. 2024, 17, 376–386. [Google Scholar] [CrossRef]
- Rawal, S.; Kaur, H.; Bhathan, S.; Mittal, D.; Kaur, G.; Ali, S.A. Ruminant Gut Microbiota: Interplay, Implications, and Innovations for Sustainable Livestock Production. Sustain. Agric. Rev. 2024, 64, 205–228. [Google Scholar]
- Basson, A.R.; Chen, C.; Sagl, F.; Trotter, A.; Bederman, I.; Gomez-Nguyen, A.; Sundrud, M.S.; Ilic, S.; Cominelli, F.; Rodriguez-Palacios, A. Regulation of intestinal inflammation by dietary fats. Front. Immunol. 2021, 11, 604989. [Google Scholar] [CrossRef] [PubMed]
- Onche, E.; Cho, H.; Rangandang, A.D.; Kang, N.; Kim, S.; Kim, H.; Seo, S. Enhancing Feed Efficiency and Growth in Early-Fattening Hanwoo Steers Through High-Energy Concentrate Feeding. Animals 2025, 15, 490. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Cidan, Y.; Ali, M.; Lu, S.J.; Javed, U.; Cisang, Z.; Gusang, D.; Danzeng, Q.; Li, K.; Basang, W. Evaluating the Effect of Dietary Protein-Energy Ratios on Yak Intestinal Microbiota Using High-Throughput 16S rRNA Gene Sequencing. Vet. Sci. 2025, 12, 208. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, J.X.; Xue, B.C.; Yue, S.M.; Yang, C.; Xue, B. Effects of premating calcium and phosphorus supplementation on reproduction efficiency of grazing Yak heifers. Animals 2021, 11, 554. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Zhang, C.H.; Liu, F.S.; Jin, Z.Y.; Xia, X.L. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. Nutr. 2023, 63, 5841–5855. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; La, Y.; Yang, G.W.; Dai, R.F.; Zhang, J.X.; Zhang, Y.H.; Jin, J.M.; Ma, X.M.; Guo, X.; Chu, M.; et al. Multi-omics revealed the effects of dietary energy levels on the rumen microbiota and metabolites in yaks under house-feeding conditions. Front. Microbiol. 2024, 14, 1309535. [Google Scholar] [CrossRef] [PubMed]
- Varghese, V.K.; Poddar, B.J.; Shah, M.P.; Purohit, H.J.; Khardenavis, A.A. A comprehensive review on current status and future perspectives of microbial volatile fatty acids production as platform chemicals. Sci. Total Environ. 2022, 815, 152500. [Google Scholar] [CrossRef]
- Leite, E.R.; Conde, A.M.J.r.; Fonseca, C.M.B.; Monteiro, H.M.D.; de Carvalho, M.A.M.; de Melo, W.G.G.; Teodoro, A.L.; Silva, D.D.; Oliveira, L.S.D.; Ferraz, J.C.B. Impact of feeding native Caatinga pasture on the rumen histomorphometry of sheep raised in semi-extensive management. Anat. Histol. Embryol. 2024, 53, e13029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, X.D.; Ding, Z.Q.; Kang, Y.D.; Guo, S.K.; Cao, M.L.; Hu, L.Y.; Xiong, L.; Pei, J.; Guo, X. Effects of High-Concentrate Diets on Growth Performance, Serum Biochemical Indexes, and Rumen Microbiota in House-Fed Yaks. Animals 2024, 14, 3594. [Google Scholar] [CrossRef]
- Liu, K.Z.; Zhang, Y.D.; Yu, Z.T.; Xu, Q.B.; Zheng, N.; Zhao, S.G.; Huang, G.X.; Wang, J.Q. Ruminal microbiota–host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef]
- Zhang, J.B.; Ren, X.L.; Wang, S.; Liu, R.D.; Shi, B.; Dong, H.L.; Wu, Q.X. Microbial interventions in yak colibacillosis: Lactobacillus-mediated regulation of intestinal barrier. Front. Cell. Infect. Microbiol. 2024, 14, 1337439. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, H.Z.; Yang, Q.; Yang, D.Y.; Liu, S.J.; Cui, Z.H. Evaluating starter feeding on ruminal function in yak calves: Combined 16S rRNA sequencing and metabolomics. Front. Microbiol. 2022, 13, 821613. [Google Scholar] [CrossRef]
- Pang, K.Y.; Wang, J.M.; Chai, S.T.; Yang, Y.K.; Wang, X.; Liu, S.J.; Ding, C.; Wang, S.X. Ruminal microbiota and muscle metabolome characteristics of Tibetan plateau yaks fed different dietary protein levels. Front. Microbiol. 2024, 15, 1275865. [Google Scholar] [CrossRef]
- Dimou, A.; Tsimihodimos, V.; Bairaktari, E. The critical role of the branched chain amino acids (BCAAs) catabolism-regulating enzymes, branched-chain aminotransferase (BCAT) and branched-chain α-keto acid dehydrogenase (BCKD), in human pathophysiology. Int. J. Mol. Sci. 2022, 23, 4022. [Google Scholar] [CrossRef]
- Fontoura, A.B. The Effects of Heat Stress and Dietary Organic Acid and Pure Botanical Supplementation on Growth and Lactation in Dairy Cattle. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2022. [Google Scholar]
- Wang, J.; Deng, L.F.; Chen, M.X.; Che, Y.Y.; Li, L.; Zhu, L.L.; Chen, G.S.; Feng, T. Phytogenic feed additives as natural antibiotic alternatives in animal health and production: A review of the literature of the last decade. Anim. Nutr. 2024, 17, 244–264. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition (%) | CKG | LEG | HEG |
|---|---|---|---|
| Maize | 0 | 29.8 | 34.4 |
| Cottonseed meal | 0 | 4.6 | 5 |
| Soybean meal | 0 | 6.0 | 6.5 |
| Soybean oil | 0 | 0 | 0.6 |
| Wheat bran | 0 | 5.8 | 1 |
| Molasses | 0 | 1.3 | 0 |
| Premix ② | 0 | 2.5 | 2.5 |
| Wheat straw | 0 | 50 | 50 |
| Native grass | 100 | 0 | 0 |
| Total | 100 | 100 | 100 |
| Nutritional composition ① ③ | |||
| CP (%) | 2.64 | 9.85 | 10.92 |
| NEG (MJ/Kg) | 0.11 | 2.12 | 2.69 |
| ADF (%) | 16.79 | 5.18 | 4.51 |
| NDF (%) | 55.05 | 43.42 | 41.84 |
| Ca | 0.18 | 1.62 | 1.59 |
| P | 0.05 | 0.55 | 0.54 |
| Groups | Initial Body Weight (kg) | Final Body Weight (kg) | Total Gain (kg) | Average Daily Gain (g/d) |
|---|---|---|---|---|
| CKG | 260.15 ± 35.55 | 279.40 ± 16.80 d | 19.25 ± 10.15 c | 113.24 ± 48.90 c |
| LEG | 257.45 ± 27.25 | 317.25 ± 23.50 c | 59.80 ± 21.70 b | 351.76 ± 98.95 b |
| HEG | 255.35 ± 25.35 | 349.85 ± 43.45 b | 94.50 ± 21.25 a | 555.88 ± 113.65 ab |
| HLG | 264.30 ± 29.20 | 367.85 ± 35.20 a | 103.55 ± 21.55 a | 609.12 ± 102.85 a |
| Groups | Acetic Acid (mmol/L) | Propanoic Acid (mmol/L) | Isobutyric Acid (mmol/L) | Butyric Acid (mmol/L) | Isovaleric Acid (mmol/L) | Valeric Acid (mmol/L) | pH | NH3-N (mg/dL) | TVFA (mmol/L) | MCP (g/L) | A/P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CKG | 39.93 ± 2.25 b | 25.33 ± 1.31 | 2.11 ± 0.09 | 22.19 ± 0.36 | 1.56 ± 0.04 b | 3.23 ± 0.08 | 6.81 ± 0.01 a | 26.19 ± 0.11 a | 95.66 ± 1.07 d | 1.92 ± 0.01 a | 1.63 ± 0.09 a |
| LEG | 43.84 ± 1.37 ab | 25.39 ± 1.54 | 2.17 ± 0.08 | 22.88 ± 0.31 | 1.61 ± 0.05 b | 3.35 ± 0.11 | 6.76 ± 0.02 b | 24.92 ± 0.07 c | 100.54 ± 1.16 c | 1.72 ± 0.03 c | 1.62 ± 0.15 b |
| HEG | 44.88 ± 1.33 ab | 27.77 ± 1.56 | 2.25 ± 0.05 | 23.17 ± 0.54 | 1.69 ± 0.11 b | 3.33 ± 0.13 | 6.51 ± 0.01 c | 25.87 ± 0.08 b | 104.31 ± 1.23 b | 1.85 ± 0.02 b | 1.60 ± 0.14 c |
| HLG | 45.48 ± 1.57 a | 28.05 ± 1.13 | 2.30 ± 0.07 | 22.77 ± 0.38 | 1.95 ± 0.11 a | 3.16 ± 0.11 | 6.42 ± 0.01 d | 24.72 ± 0.07 c | 107.04 ± 1.27 a | 1.61 ± 0.03 d | 1.61 ± 0.13 b |
| Groups | Stratum Corneum | Tunica Propria | Mucosal Epithelial Thickness | Submucosal Thickness | Muscle Layer Thickness | Epithelial Papillae Density (/mm2) | Epithelial Papillae Width (mm) | Epithelial Papillae Length (mm) |
|---|---|---|---|---|---|---|---|---|
| CKG | 27.76 ± 1.17 a | 136.94 ± 6.32 a | 102.80 ± 10.37 a | 621.45 ± 10.44 a | 5364.94 ± 137.14 a | 121.18 ± 7.46 a | 1.56 ± 0.01 a | 5.43 ± 0.18 a |
| LEG | 23.22 ± 1.44 c | 121.33 ± 5.21 b | 91.05 ± 9.20 b | 586.60 ± 10.49 c | 4469.45 ± 94.96 c | 93.83 ± 7.06 b | 1.45 ± 0.01 c | 5.05 ± 0.03 b |
| HEG | 18.72 ± 1.20 d | 114.13 ± 7.57 c | 82.20 ± 10.45 c | 564.94 ± 9.50 d | 3878.02 ± 135.23 d | 75.88 ± 5.49 c | 1.42 ± 0.01 c | 4.28 ± 0.05 c |
| HLG | 25.28 ± 1.21 b | 133.92 ± 6.39 a | 98.29 ± 11.18 a | 600.85 ± 11.23 b | 4733.37 ± 129.83 b | 114.21 ± 5.18 a | 1.52 ± 0.01 b | 4.80 ± 0.05 b |
| Groups | ACE | Chao1 | Shannon | Simpson |
|---|---|---|---|---|
| CKG | 815.66 ± 7.355 b | 819.12 ± 76.33 b | 8.91 ± 0.29 a | 0.9854 ± 0.01 b |
| LEG | 959.14 ± 27.01 ab | 963.32 ± 30.61 ab | 8.59 ± 0.07 b | 0.9916 ± 0.02 a |
| HEG | 1004.64 ± 50.09 a | 1006.22 ± 48.40 a | 8.63 ± 0.15 b | 0.9944 ± 0.01 ab |
| HLG | 1032.23 ± 43.44 a | 1032.82 ± 43.15 a | 8.64 ± 0.09 b | 0.9940 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, J.; Bao, P.; Li, N.; Kong, S.; Chu, M.; Chen, Q.; Yan, P. Lactobacillus Supplementation Modulates Rumen Microbiota and Metabolism in Yaks Under Fattening Feeding Conditions: A Comprehensive Multi-Omics Analysis. Animals 2025, 15, 1681. https://doi.org/10.3390/ani15121681
Jia J, Bao P, Li N, Kong S, Chu M, Chen Q, Yan P. Lactobacillus Supplementation Modulates Rumen Microbiota and Metabolism in Yaks Under Fattening Feeding Conditions: A Comprehensive Multi-Omics Analysis. Animals. 2025; 15(12):1681. https://doi.org/10.3390/ani15121681
Chicago/Turabian StyleJia, Jianlei, Pengjia Bao, Ning Li, Siyuan Kong, Min Chu, Qian Chen, and Ping Yan. 2025. "Lactobacillus Supplementation Modulates Rumen Microbiota and Metabolism in Yaks Under Fattening Feeding Conditions: A Comprehensive Multi-Omics Analysis" Animals 15, no. 12: 1681. https://doi.org/10.3390/ani15121681
APA StyleJia, J., Bao, P., Li, N., Kong, S., Chu, M., Chen, Q., & Yan, P. (2025). Lactobacillus Supplementation Modulates Rumen Microbiota and Metabolism in Yaks Under Fattening Feeding Conditions: A Comprehensive Multi-Omics Analysis. Animals, 15(12), 1681. https://doi.org/10.3390/ani15121681