1. Introduction
Mutton is widely produced and consumed globally due to its unique nutritional value and flavor characteristics. According to the National Bureau of Statistics, China’s mutton production has been increasing yearly, reaching 5.2 million tons in 2024 (
http://www.stats.gov.cn, accessed on 2 December 2024). However, consumer demand for mutton is gradually shifting toward high-quality products. Various factors, including the breed, age, gender, and diet of the animal, profoundly influence the quality of mutton. Among these, dietary conditions are particularly crucial in determining meat quality [
1].
Animal diet directly influences meat quality through its effects on fatty acid composition, antioxidant properties, and volatile content [
2]. In recent years, the use of plant-derived active ingredients in animal feed to improve the fatty acid composition of mutton has garnered considerable attention. Plant polyphenols, which are abundant in plants, constitute a class of compounds known for their potent antioxidant properties. Research has shown that plant polyphenols and their derived phenolic active compounds can improve hepatic lipid metabolism in mice, thereby influencing fat deposition. Furthermore, these phenolic active compounds exhibit potentially distinct regulatory effects on fat deposition [
3].
Recently, the natural phenolic compound eugenol has received significant attention. It is primarily sourced from the essential oils of plants such as cinnamon, clove, and basil. It has been found that eugenol significantly influences the aroma of beef by enhancing its flavor profile [
4]. Lourenço et al. [
5], using dual-flow continuous culture fermenters, demonstrated that adding eugenol can alter the composition of fatty acids by affecting the biohydrogenation process.
Another bioactive compound, cinnamaldehyde, extracted from cinnamon, possesses antibacterial and antiviral properties, effectively contributing to the healthy development of animals [
6]. Previous research has shown that adding cinnamaldehyde to the diet can reduce the level of C18:0, while increasing the amounts of C18:1n9c, C18:2n6c, and C18:3n3 [
7]. Additionally, cinnamaldehyde exhibits lipid-modulating effects, significantly decreasing serum triglyceride (TAG) levels while increasing plasma high-density lipoprotein cholesterol levels [
8]. Furthermore, cinnamaldehyde has been shown to upregulate the gene expression of
PPARγ, which is associated with lipid metabolism [
9].
The main constituents of chili pepper oleoresin are capsaicinoids, which can effectively attenuate obesity and metabolic syndrome by favorably modulating lipid metabolism [
10]. Capsaicinoids regulate lipid metabolism in animals by inhibiting lipoprotein lipase (
LPL) and pancreatic lipase, stimulating lipolysis, and preventing adipocyte differentiation [
11,
12,
13,
14]. The addition of capsicum oleoresin to broiler feed can increase the fatty acid composition of breast and leg muscles [
15]. Studies have shown that combining multiple natural plant extracts positively impacts animal growth performance and meat quality [
16,
17]. However, research on the effects of complex plant extracts (CPEs), which consist of eugenol, cinnamaldehyde, and capsicum oleoresin, on lipid deposition and metabolism in sheep remains limited.
Physiologically, communication occurs between fat and muscle tissues, and lipid accumulation in fat tissues can influence muscles, ultimately altering meat quality [
18]. Given that the aforementioned plant extracts affect the body’s lipid metabolism, this study aims to investigate the effects of CPE on sheep growth characteristics, back fat (BF) deposition, and other relevant indicators. Furthermore, this study utilizes lipidomics and transcriptomics technologies to examine the effects of CPE on BF deposition and elucidate the mechanisms underlying the deposition process.
2. Materials and Methods
2.1. Animal and Experimental Design
The CPE (Pancosma XTRACT
® 7065) used in this study was provided by Pancosma (Shanghai, China) Feed Additives Co., Ltd. (Shanghai, China). The manufacturer guarantees that the active ingredient content of the microcapsule product is 5.5% cinnamaldehyde, 9.5% eugenol, and 3.5% capsicum oleoresin. A previous study showed that adding 80 mg/kg of CPE improved the production performance of ewes; hence, 80 mg/kg was selected as the dosage for this experiment [
19].
Animal tests were conducted at the Hailiutu Experimental Base of Inner Mongolia Agricultural University. The experimental protocol was approved by the Inner Mongolia Agricultural University Research Ethics Committee (permission number [2020]065). The experiment lasted 75 days, including 15 days of pre-feeding and 60 days of the formal period. To eliminate sex differences, only female sheep were selected. Thirty-six sheep, approximately 4 months old with similar weight (29.92 ± 2.52 kg), were randomly divided into two groups. Each group contained six pens (six replicates), with three sheep raised in each pen. The experimental basal diet was formulated according to the meat sheep feeding standard (NY/T816-2004). The CTRL group was fed the basal diet without CPE, while the CPE group received the same diet supplemented with 80 mg/kg of CPE. The ingredients and nutrient levels of the basal diet are shown in
Table 1.
Animals were fed twice daily (08:00 a.m. and 05:00 p.m.) to ensure a 10% refusal rate. Water was freely available throughout the experiment period. On the 61st day of the trial, one sheep was randomly selected from each pen, so a total of six sheep were slaughtered per group. Food was withheld for 12 h before slaughter, while water was restricted for 2 h. The animals were humanely slaughtered following Islamic ritual methods by severing the carotid artery and trachea. Approximately 15 cm of the BF tissue was collected, snap frozen in liquid nitrogen, and stored at −80 °C for fatty acid composition and lipidomic and transcriptomic analyses.
2.2. Carcass Traits
On day 61 of the trial period, six sheep were selected from each group for slaughter. Live weight was recorded prior to slaughter. Following slaughter, the head, hooves, and internal organs were removed, and the carcass was weighed after skin removal. The tissue thickness between the 12th and 13th ribs of the left carcass was measured using a vernier caliper at a distance of 11 cm from the spine’s central axis. This measurement was recorded as the carcass fat content (GR value). The muscle area of the longissimus dorsi between the 12th and 13th ribs of the left carcass was traced on sulfate paper and recorded as the loin eye area [
20].
The main hot carcass weight, slaughter rate, and loin eye area were calculated using the following formulas:
Carcass weight (kg) = live weight before slaughter − weight of head, hooves, skin, hair, reproductive organs and surrounding fat, and internal organs (with kidneys and surrounding fat).
Slaughter rate (%) = (hot carcass weight ÷ live weight before slaughter) × 100.
Loin eye area (cm2) = eye muscle height × eye muscle width × 0.7.
2.3. Determination of Fatty Acid Composition
Sample pre-treatment: After slaughter, the tissue was weighed (55 mg), homogenized with 4 mL of n-hexane, and shaken at 50 °C for 30 min. Then, 3 mL of 0.4 mol/L KOH methanol solution was added and shaken at 50 °C for another 30 min. Next, 1 mL of H2O was added and mixed well. After phase separation, the upper layer was carefully collected, and an internal standard (methyl nonadecanoate, Sigma, N5377, Sigma-Aldrich, St. Louis, MO, USA) was added for gas chromatography–mass spectrometry (GC-MS) analysis.
Chromatographic analysis: The treated samples were analyzed using an Agilent chromatographic system (GC-MS 7890B-5977A, Agilent, Santa Clara, CA, USA) with the GC-MS external standard method. A DB-23 column (30 m × 320 μm × 0.25 μm) was used. The inlet and detector temperatures were set at 250 °C and 230 °C, respectively. Shunt injection was performed with a split ratio of 1:5. Helium was used as the carrier gas, and the sample injection volume was 1 μL. The heating procedure of the column box was as follows: (a) the initial temperature was maintained at 50 °C for 1 min; (b) then, the temperature was increased to 175 °C at a rate of 25 °C/min; (c) finally, the temperature was raised to 230 °C at a rate of 4 °C/min, and the whole heating procedure was held for 24.75 min.
2.4. Lipid Extraction, UHPLC-MS/MS Analysis, and Data Search
Lipid metabolite extraction and analysis followed the method described by Guo et al. [
21]. Lipids were extracted using MTBE/methanol/water (10:3:2.5,
v/
v/
v) with phase separation by centrifugation (1000×
g, 10 min). The organic phase was dried under nitrogen, reconstituted in isopropanol, and analyzed by UHPLC-MS/MS (Q Exactive™ HF, Thermo Fisher, Waltham, MA, USA) with a C30 column (150 × 2.1 mm, 40 °C). Gradient elution (0.35 mL/min): 30% B (acetonitrile/isopropanol, 10:90) to 99% B over 16 min. Mobile phases contained 10 mM ammonium acetate and 0.1% formic acid. MS detection used a positive/negative switching mode (spray voltage 3 kV,
m/
z 114–1700) with a collision energy of 25–30 eV. Lipid raw data were then qualitatively and quantitatively analyzed, with data quality control applied before proceeding to bioinformatic analysis. Partial least squares discriminant analysis (PLS-DA) was performed using Python (version 3.5.0) to investigate group separation.
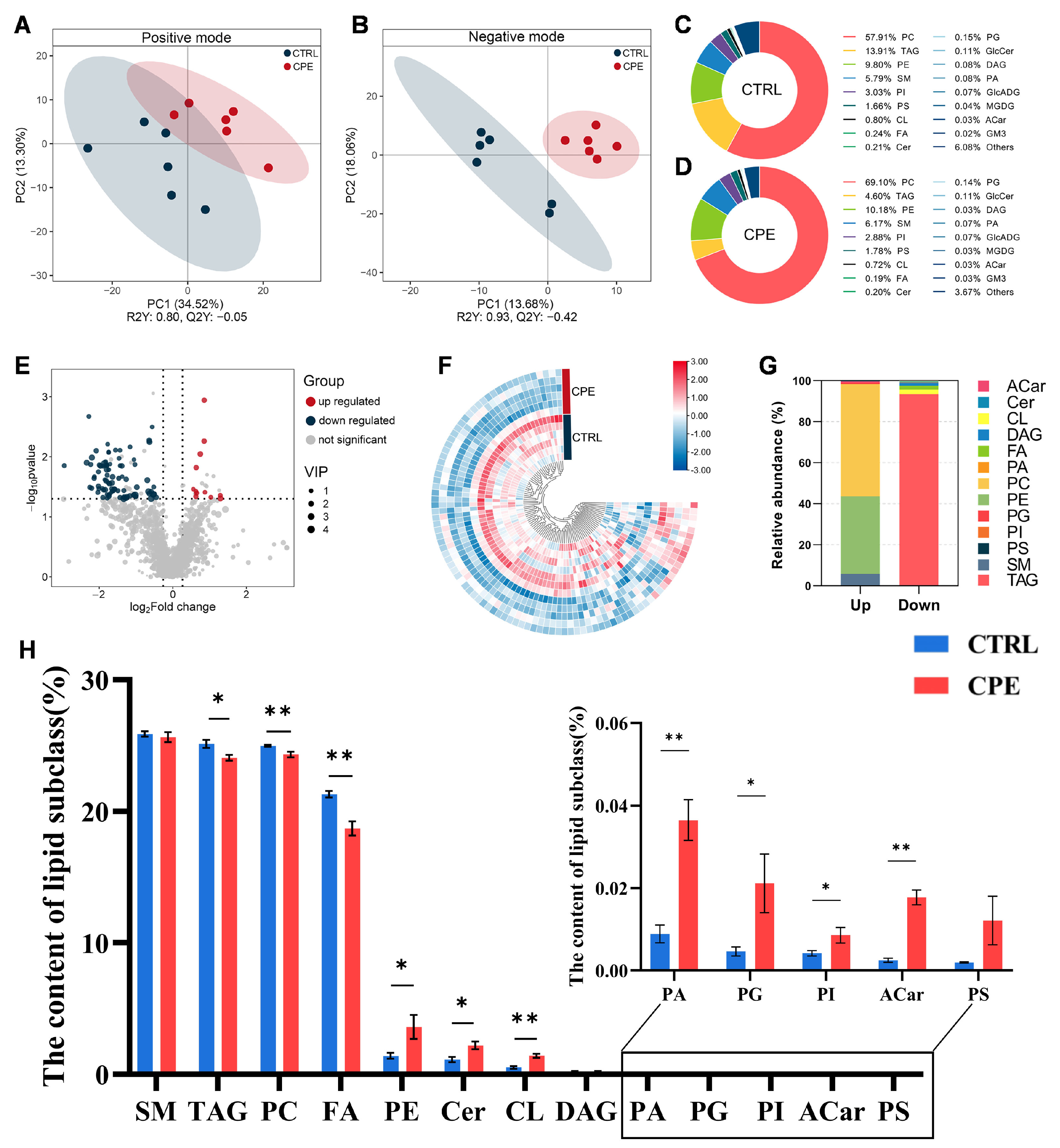
Differentially altered lipid molecules (DALs) between the CTRL and CPE groups were identified based on the following criteria: variable importance in projection (VIP) > 1.0, fold change (FC) > 1.2, or FC < 0.833 and p < 0.05 (calculated using an independent t-test). The VIP score was derived from the first principal component of the PLS-DA model, reflecting the contribution of each lipid molecule to group discrimination. The relative abundance of each lipid species was expressed as a percentage of total lipid molecules.
Pearson correlation analysis was performed to assess the relationship between individual DALs in each group using “SRplot”, an online platform for data analysis and visualization (accessed on 9 November 2023,
https://www.bioinformatics.com.cn).
2.5. Transcriptome Analysis
RNA extraction and analysis followed the method described by Guo et al. [
21]. Raw FASTQ data (raw reads) were initially processed using in-house Perl scripts. Clean data (clean reads) were obtained by removing adapter-containing, ploy-N-containing, and low-quality reads from the raw data. The clean data’s Q20, Q30, and GC contents were also determined. All subsequent analyses were conducted using high-quality and clean data.
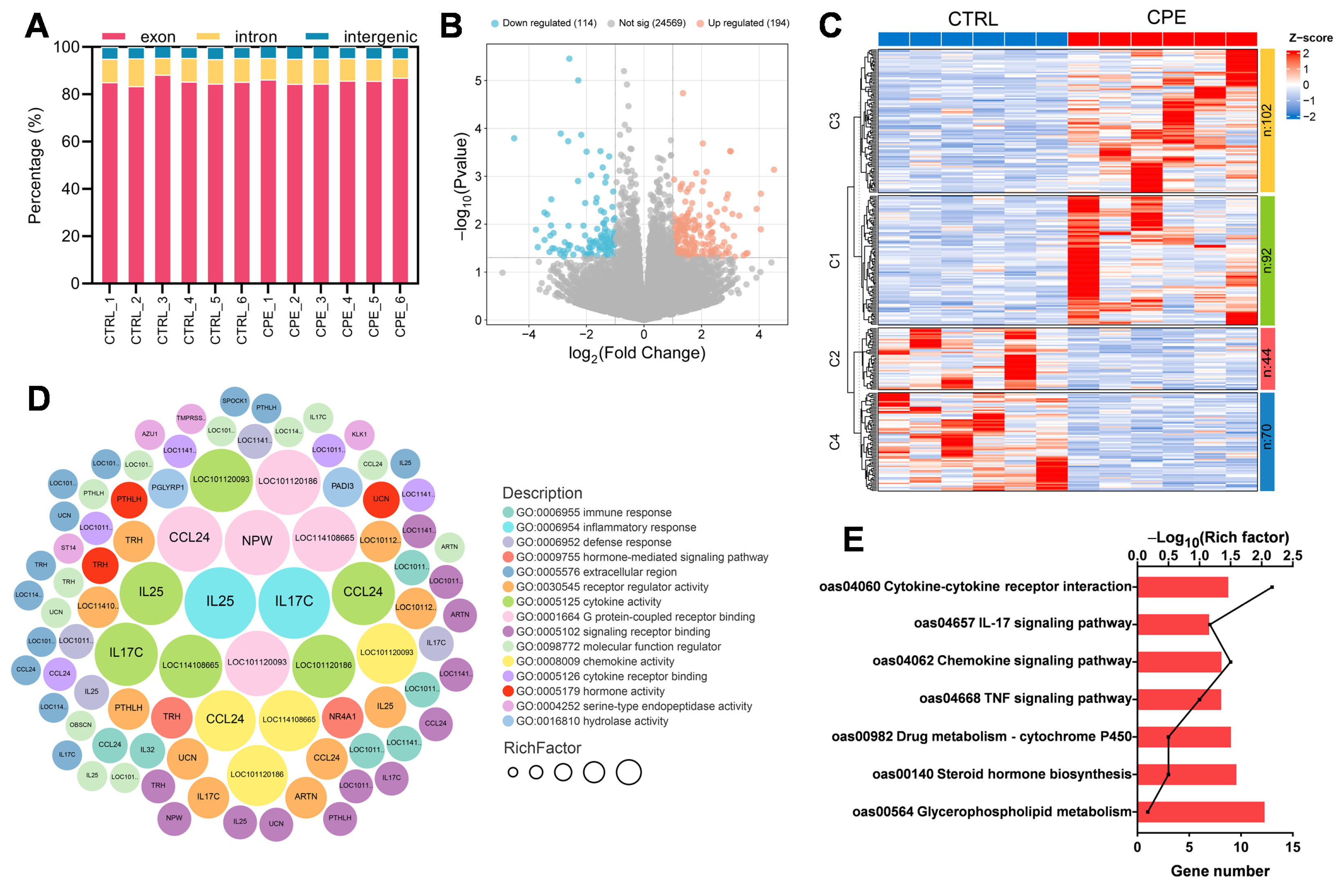
Differential expression analysis between the CTRL and CPE groups was performed using the DESeq2 R package (version 1.20.0). Genes were considered differentially expressed genes (DEGs) if they met the threshold of |log2(FoldChange)| > 1 and p < 0.05. To assess the functional significance of the identified DEGs, an enrichment analysis of the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways and Gene Ontology (GO) terms was performed using the clusterProfiler R package (version 3.8.1). GO enrichment analysis was performed for the Biological Process (BP), Molecular Function (MF), and Cellular Component (CC) categories, while KEGG pathway analysis identified relevant biological pathways associated with differentially expressed genes.
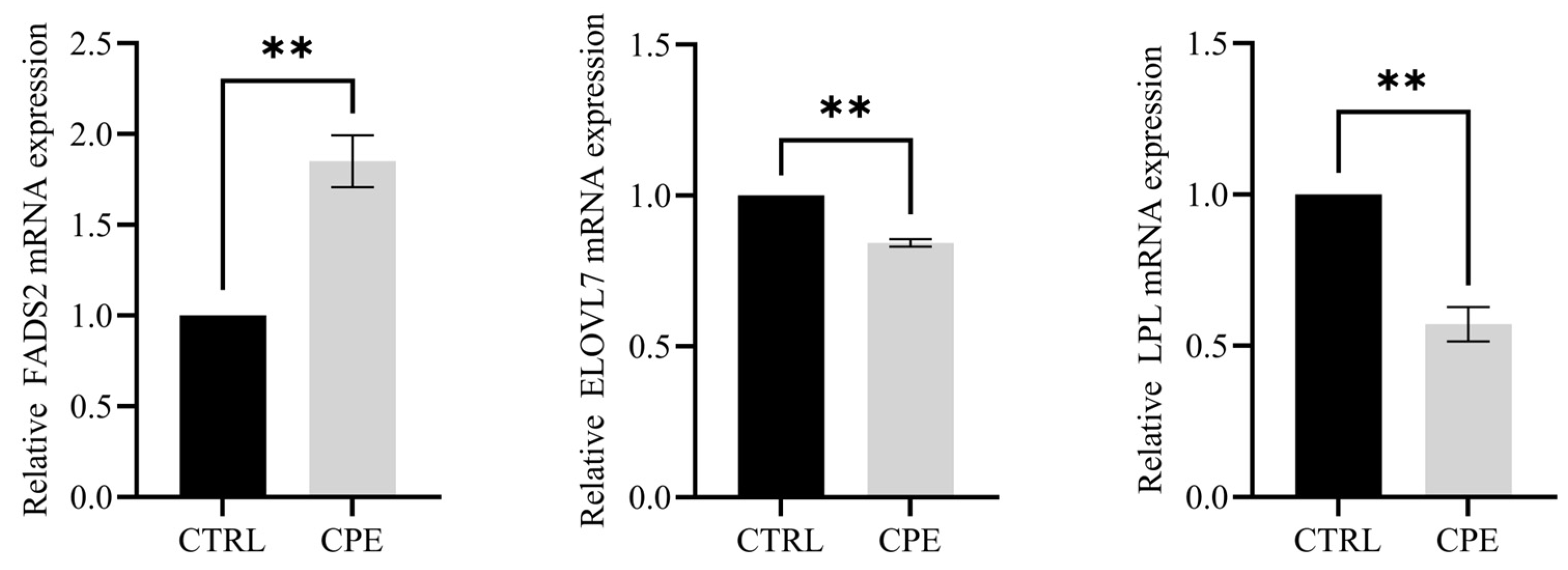
2.6. Quantitative Real-Time PCR (qRT-PCR) Verification
The expression of candidate DEGs was verified using qRT-PCR as described by Guo et al. (2025) [
21]. The mRNA sequences of sheep fatty acid desaturases 2 (
FADS2), the elongase of very-long-chain fatty acids 7 (
ELOVL7), and LPL were retrieved from the official National Center for Biotechnology Information (nih.gov).
Primers were designed using Primer Premier 5.0 software, and the primer sequences (
Table 2) were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). The relative gene expression levels of the
FADS2,
ELOVL7, and
LPL were calculated using the 2
−∆∆CT method [
22].
2.7. Statistical Analysis and Data Visualization
After organizing and preprocessing the raw data using Excel 2021 software, data analyses were performed using the PROC GLM procedure in Statistical Analysis Software (SAS version 9.2; Cary, NC, USA) to assess treatment effects. Model assumptions, including the normality and homogeneity of variances, were checked using the Shapiro–Wilk and Levene’s tests, respectively.
Results were expressed by least square means with estimated standard error (SEM). Statistical significance was set at p < 0.05, with tendencies considered at 0.05 ≤ p < 0.10.
For data visualization, TBtools (v0.6673) [
23] was used for bioinformatics analysis and graphical representation, GraphPad Prism v8 (GraphPad Software, San Diego, CA, USA) was employed for statistical plotting, and OmicShare [
24] was utilized for pathway enrichment and network visualization.
4. Discussion
The GR value serves as a crucial indicator, reflecting the content of carcass adipose tissue, wherein higher values denote greater fat deposition. A previous study suggested that supplementing plant extracts in diets can enhance lipid accumulation in livestock [
25]. In this study, the addition of CPE did not affect the live weight before slaughter, hot carcass weight, slaughter rate, and lion eye area but significantly increased GR values, suggesting that CPE promotes adipose tissue deposition in sheep.
The composition and content of FAs in adipose tissue are closely associated with its nutritional value. SFAs have long been considered detrimental to human health due to their association with an increased risk of cardiovascular diseases, raising concerns regarding their excessive intake. However, our results demonstrate a significant reduction in the SFA content following CPE supplementation. This reduction in SFA levels may be attributed to the unique properties of CPE, which potentially inhibit the synthesis of SFAs.
Conversely, higher levels of UFAs and lower levels of SFAs have been linked to improved lipid profiles, particularly through the reduction in LDL-C, thereby lowering the risk of cardiovascular diseases [
26]. The conversion of SFA to UFA necessitates a series of desaturase-catalyzed reactions, primarily yielding beneficial MUFAs such as C16:1 and C18:1n9c [
27]. In this study, the addition of CPE significantly increased the MUFA content, indicating that CPE supplementation not only promotes adipose tissue deposition but also improves fat deposition. The observed decrease in SFAs and increase in MUFAs suggest that CPE enhances the nutritional value of meat products by shifting the lipid profile toward a more favorable composition.
Furthermore, C18:1n9c, the most abundant and beneficial MUFA in ruminants, produces distinct flavor compounds during cooking and has been shown to exhibit protective effects against cardiovascular diseases [
28]. In this study, the addition of CPE resulted in a significant increase in the C18:1n9c content, thereby enhancing both the sensory quality and health value of sheep meat.
Interestingly, PUFA levels, the PUFA/SFA ratio, n-6 PUFA, and n-3 PUFA were significantly reduced following CPE supplementation. Research suggests that mammals lack the Δ-15 desaturase enzymes, which are necessary for the endogenous synthesis of PUFAs, making dietary intake the primary source [
29]. Therefore, the reduction in PUFA content in the BF of sheep fed with CPE may be attributed to the phenolic compounds’ inhibitory effect on the enzyme activity required for PUFA synthesis.
Lipid peroxidation is a major factor contributing to meat quality deterioration, affecting color stability and nutritional value, and may lead to the formation of potentially toxic compounds. Studies indicate lipid oxidation strongly correlates with lipid molecules, particularly TAG, which belongs to the GL and GP classes [
30,
31]. Since fatty acid composition plays a critical role in lipid oxidation, a lower PUFA content is often associated with enhanced oxidative stability of meat. Specifically, n-3 PUFA has been linked to increased lipid oxidation and TAG content reduction [
32,
33].
Lipidomics analysis revealed a decrease in TAG content in sheep BF following CPE supplementation, along with a significant reduction in n-3 PUFA content. This suggests that CPE supplementation may decrease TAG content by lowering n-3 PUFA levels, thereby mitigating lipid oxidation and enhancing meat quality. Additionally, elevated free fatty acid levels in the blood have been associated with increased TAG accumulation in the liver and muscle. This can inhibit fat synthesis while promoting TAG breakdown, ultimately leading to reduced de novo SFA synthesis [
34]. Consistent with this, our study found a significant decrease in both TAG and SFA content following CPE supplementation, further supporting the role of CPE in modifying lipid metabolism and enhancing the nutritional quality of sheep meat.
Similarly, a study by Liu [
35] demonstrated that adding phenol-rich black artemisia to cashmere goat diets led to a significant reduction in TAG content in the longissimus dorsi muscle, thereby enhancing the nutritional quality of mutton. In this experiment, the lipid subclasses PE, PI, PC, PG, CL, and PA were significantly regulated after adding CPE. These subclasses are associated with GP, which plays a key role in lipid oxidation. Among them, PC and PE are the most abundant. Research indicates that the composition of GP is closely linked to the flavor profile of meat products [
36].
PC, a major structural component of cell membranes, consists of glycerol, two fatty acid chains, and one phosphate ester group, constituting 45% to 55% of the total phospholipid in mammalian cells and organelles. PC metabolism is closely intertwined with fatty acid metabolism, where UFAs within PC are highly susceptible to oxidation, influencing both fatty acid synthesis and breakdown. Consistent with our findings, CPE supplementation led to a significant reduction in PC content and a notable increase in UFA content, suggesting that PC metabolism could modulate the fatty acid composition of BF by regulating UFA levels.
SFAs typically found in PC include C18:0 and C16:0. In contrast, UFAs include C18:1, C18:2, C18:3, and C20:4. The findings from this study showed a significant decrease in PC content and fatty acids within PC in BF following CPE supplementation. Similarly, PE is involved in glycine, serine, threonine, and glycerophospholipids metabolic pathways [
37]. Li et al. [
38] demonstrated that shallot extract can enhance meat quality by modulating PE content. Consistent with our study’s findings, adding CPE upregulates PE levels and subsequently modulates the fatty acid composition in BF.
Fatty acid desaturases (FADS) are crucial in PUFA biosynthesis, catalyzing dehydrogenation reactions that introduce double bonds at specific acyl chain positions. FADS2 and FADS6 function as Δ-6 desaturases, which are responsible for inserting double bonds at Δ-6 (Δ-12) positions. Through transcriptomics analysis, Wang et al. [
39] identified FADS2 as a candidate gene in lipid metabolism, modulating fatty acid unsaturation. Consistent with our findings, dietary CPE supplementation significantly upregulated FADS2 gene expression, resulting in a decrease in SFA content and an increased UFA content, indicating FADS2’s regulatory role in the fatty acid composition. Li et al. [
40] found that supplementing Hu sheep’s diet with grape seed tannin extract, a phenolic substance, upregulated the expression of the FADS2 gene, leading to a significant reduction in C18:0 content.
The elongase of the very-long-chain fatty acid (ELOVLs) family is another key regulator of lipid metabolism and has garnered considerable interest. ELOVLs, primarily in the endoplasmic reticulum, are crucial rate-limiting enzymes in long-chain PUFA synthesis [
41]. Specifically, ELOVL7 catalyzes the elongation of both SFAs and PUFAs [
42,
43]. In line with this study, dietary CPE supplementation notably downregulated ELOVL7 gene expression, resulting in a reduction in SFA and PUFA contents. Previous research indicates that the reduction in C18:1n9c content results from ELOVL7 overexpression, with C18:1n9c content exhibiting a negative correlation with ELOVL7 expression, affirming ELOVL7’s involvement in fatty acid metabolism [
43]. Consistently, CPE supplementation downregulated ELOVL7 gene expression while significantly increasing C18:1n9c content, further supporting its role in fatty acid metabolism.
LPL serves as a crucial rate-limiting enzyme in the hydrolysis of TAG into glycerol and free fatty acids [
44]. TAG, transported in the bloodstream via very-low-density lipoprotein (VLDL), undergoes hydrolysis by LPL, generating free fatty acids that are either stored in adipose tissue or used for energy metabolism. The expression of LPL is subject to regulation by hormones, fasting, supplementary feeding, and dietary factors [
45]. Tian Jia [
46] discovered, in their study, a significant negative correlation between LPL gene expression and UFA content in the triceps brachii muscle of Altai sheep. Similarly, Shimomura et al. [
47] posited that elevated UFA levels could disrupt the transcription of lipase genes, consequently impeding body metabolism and downregulating LPL gene expression. In agreement with these findings, the UFA content significantly increased while LPL gene expression decreased after dietary supplementation with CPE, suggesting that CPE might augment UFA content by suppressing LPL gene expression.
Notably, capsaicin exhibited an inhibitory effect on LPL activity, indicating a potential regulatory role of chili oleoresin present in CPE on sheep BF lipid metabolism via LPL activity modulation. Moreover, the reduction in TAG content observed in this study suggests that the LPL-mediated hydrolysis of TAG was suppressed, leading to lower free fatty acid availability and decreased TAG accumulation in adipose tissue [
48]. Consequently, the reduced expression of the LPL gene leads to decreased free fatty acids produced by lipid metabolism, subsequently lowering TAG accumulation in adipose tissue. In conclusion,
FADS2,
ELOVL7, and
LPL are potential key genes regulating fatty acid composition in BF.
Earlier research suggested that fatty acids deposited in muscle tissue primarily originate from the bloodstream. However, subsequent studies have demonstrated that amino acids regulate bodily fatty acid metabolism. Hayashi [
49] observed a significant enhancement in lymph lipid transport within the rat intestine when amino acids were supplemented in the diet, indicating an interplay between amino acid and fatty acid metabolism. A study confirmed that the phenols in the CPE utilized in this study possess lipid peroxidation-reducing properties [
27]. A metabolomics study by Jintao [
35] demonstrated that supplementing the diet of Artemisia nigrosalis with phenolic substances could mitigate blood lipid peroxidation, facilitate lipid decomposition, lower blood TAG content, and decrease blood C16:0 and C18:0 levels via the arginine and proline metabolic pathways. Moreover, a reduction in the deposition of SFAs, such as C16:0 and C18:0, in muscle was observed, improving meat quality.
Correspondingly, the dietary supplementation of CPE led to a significant reduction in C16:0, C18:0, and SFA levels in BF, along with a downward trend in TAG content. KEGG pathway enrichment analysis identified several DEGs significantly associated with the arginine and proline metabolic pathways. These findings suggest that CPE might mitigate lipid peroxidation by these metabolic pathways, consequently enhancing sheep BF’s fatty acid composition. The synthesis of fatty acids requires considerable energy, including n-3 PUFA. Research indicates that C16:1 can enhance energy availability for white fat metabolism by stimulating lipolysis, glucose uptake, and utilization [
50,
51,
52]. With CPE supplementation, the C16:1 content in sheep BF increased, whereas n-3 PUFA levels decreased notably. This phenomenon might be attributed to a reduction in TAG content in BF, which typically releases ample ATP upon decomposition.
Furthermore, most DEGs significantly associated with the adenosine triphosphate pathway showed a decrease, suggesting a reduced energy demand for n-3 PUFA synthesis. Moreover, KEGG enrichment analysis revealed significant enrichment of pathways such as the IL-17 and TNF signaling pathways, which are associated with inflammation.
Lipid oxidation regulates various physiological processes, including inflammation, immune defense, and metabolic stress [
53]. Putman et al. [
54] reported a strong association between lipid oxidation and an increased incidence of postpartum inflammatory diseases in cows. In the present study, dietary supplementation with CPE markedly decreased n-3 PUFA and TAG levels in BF, thereby attenuating the lipid oxidation process. Hence, this suggests that the regulatory impact of CPE on lipid metabolism in adipose tissue may be linked to modulating the inflammatory response. However, further research is needed to elucidate the underlying mechanism.


 ,
, 

{kind=link}
{kind=link}
{kind=link}