
Dietary Supplementation of Lactobacillus reuteri Modulates Amino Acid Metabolism and Extracellular Matrix in the Gut–Liver Axis of Weaned Piglets

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
2.2. Quantitative Real-Time PCR
2.3. H&E Staining & Masson Staining
2.4. Free Amino Acids Concentration Analysis
2.5. Untargeted Metabolomics Analysis
2.6. Statistical Analysis
3. Results
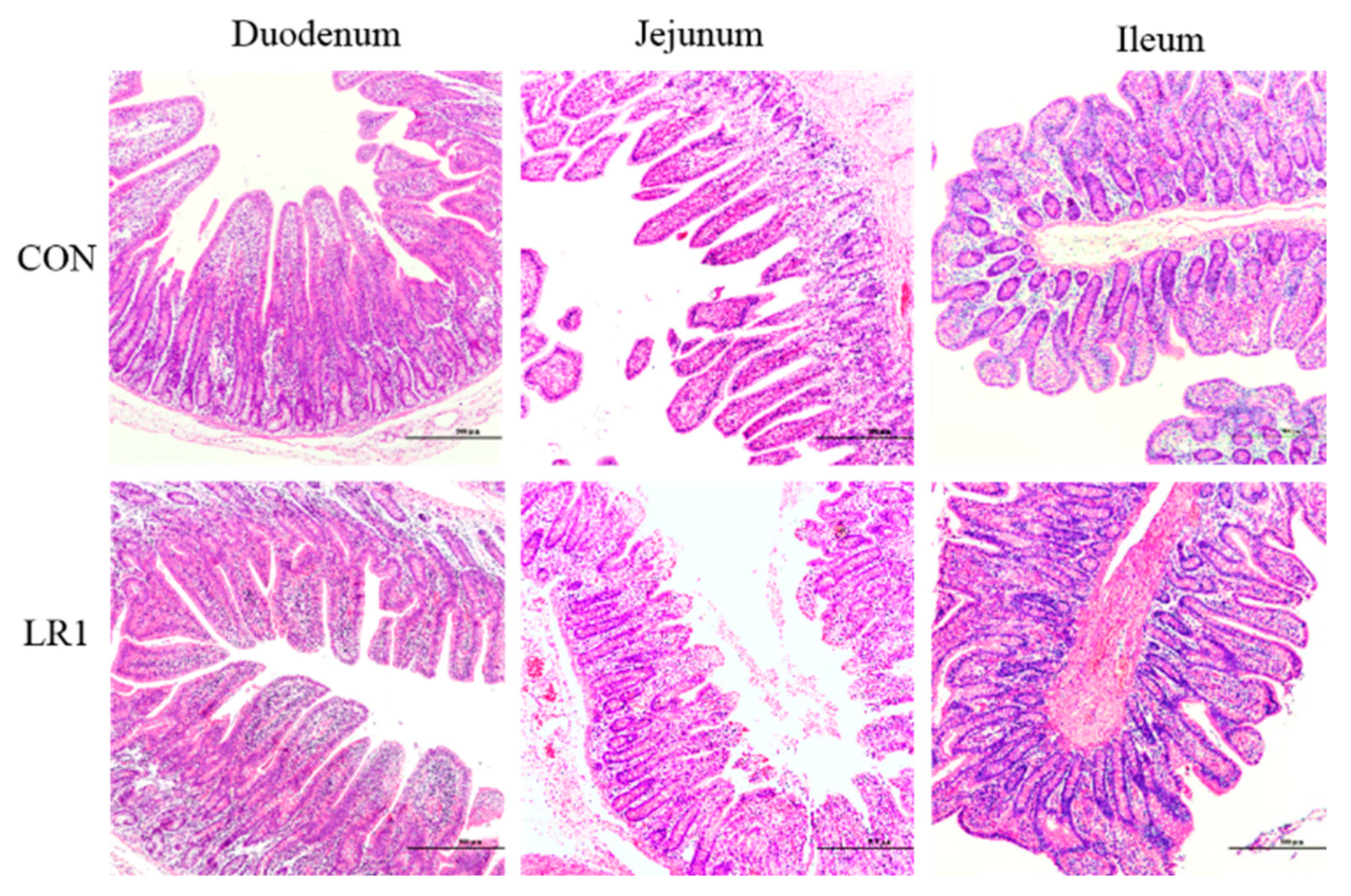
3.1. Growth Performance and Intestinal Morphology of Weaned Piglets
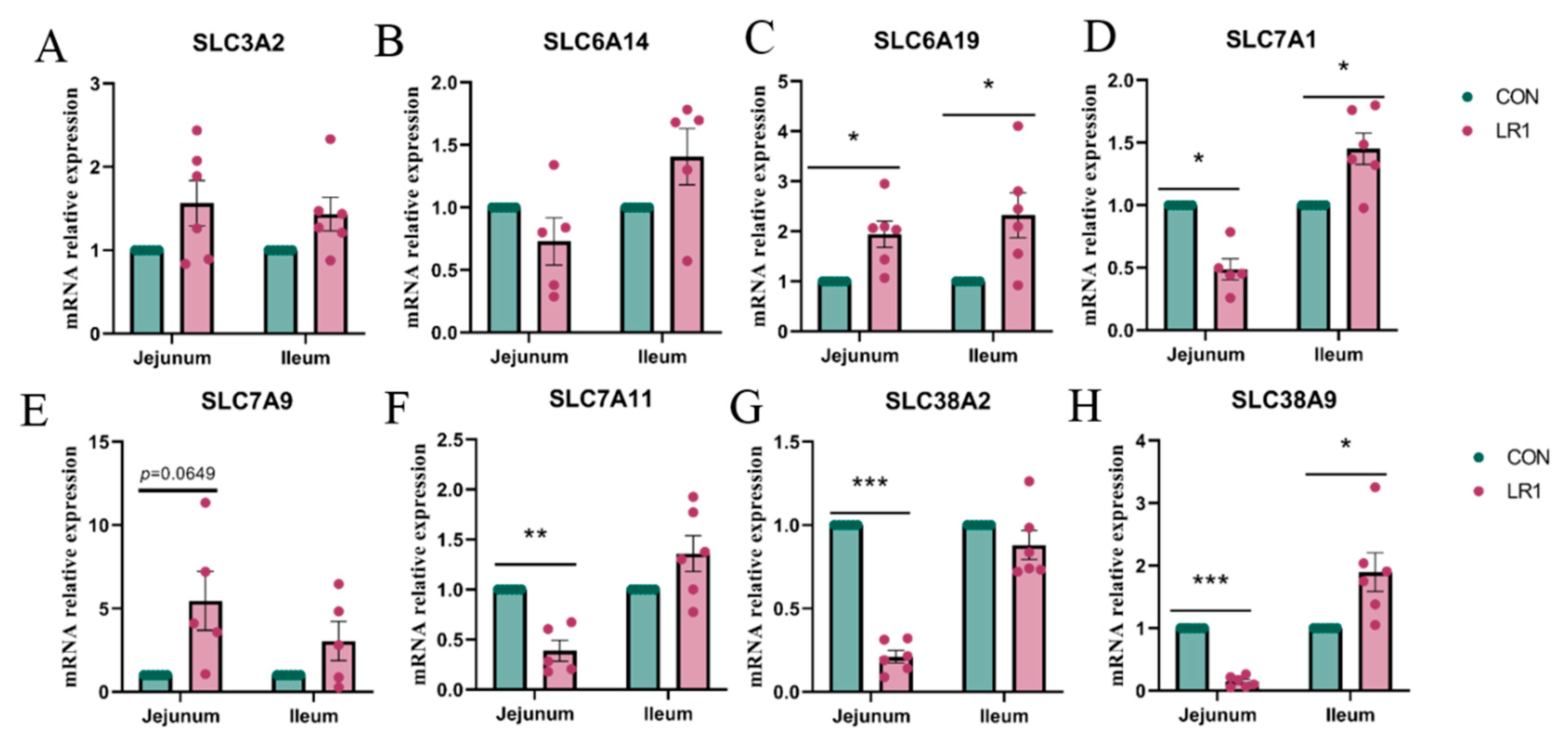
3.2. Amino Acid Transport in the Gut–Liver Axis of Piglets
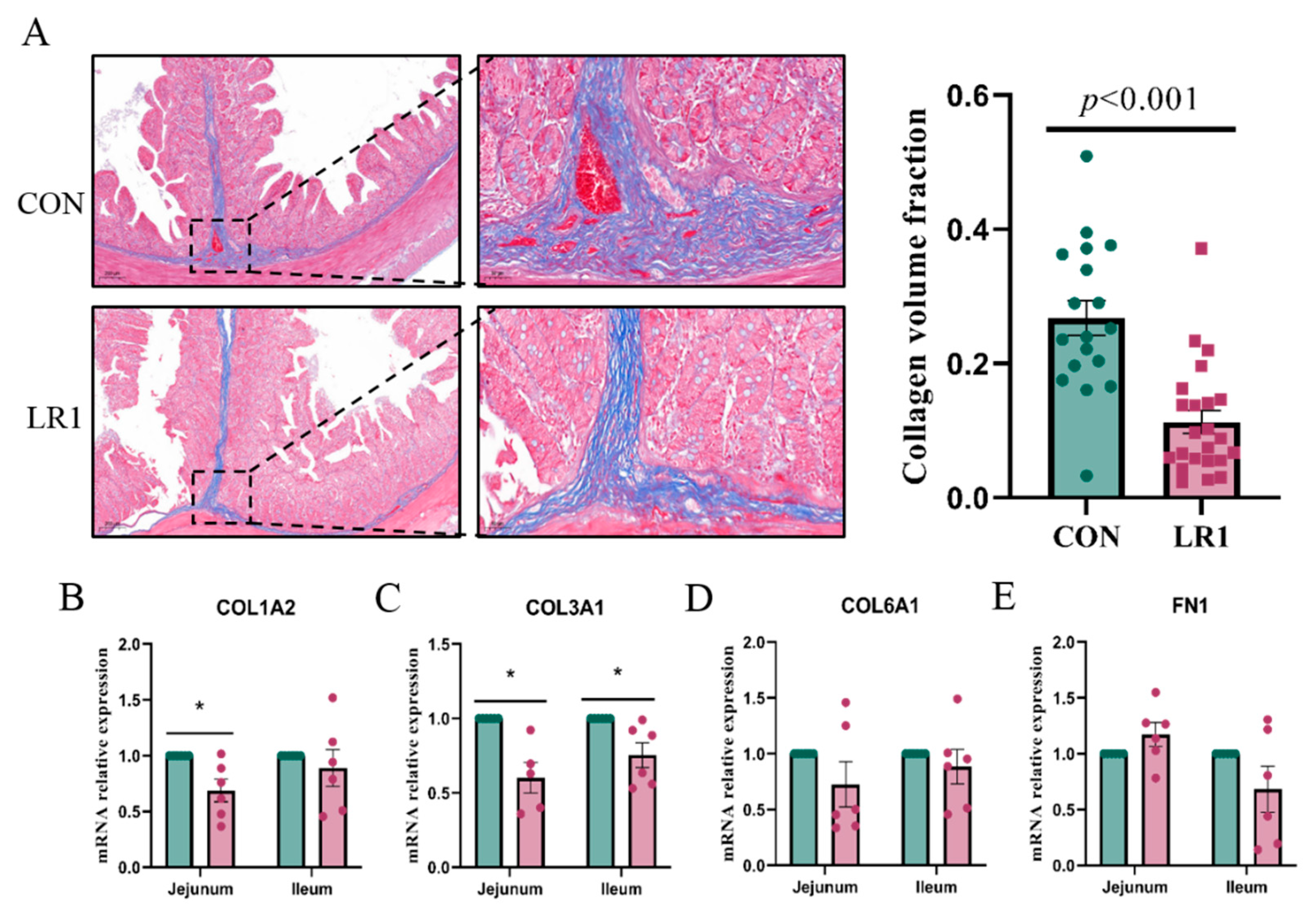
3.3. Structural Changes in the Extracellular Matrix of the Piglet Intestine
3.4. Metabolomics Analysis of Hepatic Portal Vein Plasma in Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Long, D.W.; Long, B.D.; Nawaratna, G.I.; Wu, G. Oral Administration of L-Arginine Improves the Growth and Survival of Sow-Reared Intrauterine Growth-Restricted Piglets. Animals 2025, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.N.; Spencer, J.D.; Almeida, F.R.C.L.; Patterson, J.L.; Chiarini-Garcia, H.; Dyck, M.K.; Foxcroft, G.R. Consequences of a low litter birth weight phenotype for postnatal lean growth performance and neonatal testicular morphology in the pig. Animal 2013, 7, 1681–1689. [Google Scholar] [CrossRef]
- Kirkden, R.D.; Broom, D.M.; Andersen, I.L. Invited review: Piglet mortality: Management solutions. J. Anim. Sci. 2013, 91, 3361–3389. [Google Scholar] [CrossRef]
- Geiping, L.; Hartmann, M.; Kreienbrock, L.; Grosse, B.E. Killing underweighted low viable newborn piglets: Which health parameters are appropriate to make a decision? Porc. Health Manag. 2022, 8, 25. [Google Scholar] [CrossRef]
- Dumitru, A.; Matei, E.; Cozaru, G.C.; Chisoi, A.; Alexandrescu, L.; Popescu, R.C.; Butcaru, M.P.; Dumitru, E.; Rugina, S.; Tocia, C. Endotoxin Inflammatory Action on Cells by Dysregulated-Immunological-Barrier-Linked ROS-Apoptosis Mechanisms in Gut-Liver Axis. Int. J. Mol. Sci. 2024, 25, 2472. [Google Scholar] [CrossRef]
- Chandel, N.S. Amino Acid Metabolism. Cold Spring Harb. Perspect. Biol. 2021, 13, a040584. [Google Scholar] [CrossRef]
- Azad, H.; Akbar, M.Y.; Sarfraz, J.; Haider, W.; Ghazanfar, S. Simulation studies to identify high-affinity probiotic peptides for inhibiting PAK1 gastric cancer protein: A comparative approach. Comput. Biol. Chem. 2025, 115, 108345. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; van de Graaf, S. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, Y.; Zhang, Y.; Dong, J.; Jiang, S.; Tang, Y. DHA-enriched phosphatidylserine ameliorates cyclophosphamide-induced liver injury via regulating the gut-liver axis. Int. Immunopharmacol. 2024, 140, 112895. [Google Scholar] [CrossRef]
- Perumal, S.K.; Arumugam, M.K.; Osna, N.A.; Rasineni, K.; Kharbanda, K.K. Betaine regulates the gut-liver axis: A therapeutic approach for chronic liver diseases. Front. Nutr. 2025, 12, 1478542. [Google Scholar] [CrossRef]
- Mcpeek, A.C.; Patton, B.; Columbus, D.A.; Olver, T.D.; Rodrigues, L.A.; Sands, J.M.; Weber, L.P.; Ferguson, D.P. Low birth weight and reduced postnatal nutrition lead to cardiac dysfunction in piglets. J. Anim. Sci. 2023, 101, skad364. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Wang, H.; Zhang, Q.; Chen, J.; Zhang, X.; Chen, W. Effects of intestinal nutrition and health additives on growth performance, serum biochemical index and digestive enzyme activity in weakly stiff pigs. Chin. J. Vet. Sci. 2019, 39, 2082–2087. [Google Scholar] [CrossRef]
- You, M.; Zhou, L.; Wu, F.; Zhang, L.; Zhu, S.X.; Zhang, H.X. Probiotics for the treatment of hyperlipidemia: Focus on gut-liver axis and lipid metabolism. Pharmacol. Res. 2025, 214, 107694. [Google Scholar] [CrossRef]
- Niu, X.; Chang, G.; Xu, N.; Li, R.; Niu, B.; Mao, R.; Wang, S.; Li, G.; Jiang, J.; Wang, L. Vitamin A-Integrated Cinnamaldehyde Nanoemulsion: A Nanotherapeutic Approach to Counteract Liver Fibrosis via Gut-Liver Axis Modulation. ACS Nano 2025, 19, 10433–10451. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Yi, H.; Yang, G.; Xiong, Y.; Wu, Q.; Xiao, H.; Wen, X.; Yang, X.; Wang, L.; Jiang, Z. Integrated metabolomic and proteomics profiling reveals the promotion of Lactobacillus reuteri LR1 on amino acid metabolism in the gut-liver axis of weaned pigs. Food Funct. 2019, 10, 7387–7396. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, B.; Jin, T.; Ocansey, D.; Jiang, J.; Mao, F. Intestinal Fibrosis in Inflammatory Bowel Disease and the Prospects of Mesenchymal Stem Cell Therapy. Front. Immunol. 2022, 13, 835005. [Google Scholar] [CrossRef]
- Elowsson, R.L.; Lofdahl, A.; Ahrman, E.; Muller, C.; Notermans, T.; Michalikova, B.; Rosmark, O.; Zhou, X.H.; Dellgren, G.; Silverborn, M.; et al. Matrisome Properties of Scaffolds Direct Fibroblasts in Idiopathic Pulmonary Fibrosis. Int. J. Mol. Sci. 2019, 20, 4013. [Google Scholar] [CrossRef]
- Bertero, T.; Oldham, W.M.; Grasset, E.M.; Bourget, I.; Boulter, E.; Pisano, S.; Hofman, P.; Bellvert, F.; Meneguzzi, G.; Bulavin, D.V.; et al. Tumor-Stroma Mechanics Coordinate Amino Acid Availability to Sustain Tumor Growth and Malignancy. Cell Metab. 2019, 29, 124–140. [Google Scholar] [CrossRef]
- Seo, B.R.; Chen, X.; Ling, L.; Song, Y.H.; Shimpi, A.A.; Choi, S.; Gonzalez, J.; Sapudom, J.; Wang, K.; Andresen, E.R.; et al. Collagen microarchitecture mechanically controls myofibroblast differentiation. Proc. Natl. Acad. Sci. USA 2020, 117, 11387–11398. [Google Scholar] [CrossRef]
- Yang, K.M.; Jiang, Z.Y.; Zheng, C.T.; Wang, L.; Yang, X.F. Effect of Lactobacillus plantarum on diarrhea and intestinal barrier function of young piglets challenged with enterotoxigenic Escherichia coli K88. J. Anim. Sci. 2014, 92, 1496–1503. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Wang, L.; Xiong, Y.; Wen, X.; Wang, Z.; Yang, X.; Gao, K.; Jiang, Z. Effects of Lactobacillus reuteri LR1 on the growth performance, intestinal morphology, and intestinal barrier function in weaned pigs. J. Anim. Sci. 2018, 96, 2342–2351. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q. Dietary H. illucens Larvae Meal Improves Growth Performance and Intestinal Barrier Function of Weaned Pigs Under the Environment of Enterotoxigenic Escherichia coli K88. Master’s Thesis, Guizhou University, Guiyang, China, 2022. [Google Scholar]
- Liu, C. Effects of Lactobacillus reuteri from Pigs on Intestinal Barrier Function and Extracellular Matrix of Weaned Piglet. Master’s Thesis, Guangxi University, Nanning, China, 2023. [Google Scholar]
- Yang, G. Effects of Lactobacillus reuteri LR1 on the Growth Performance, Intestinal Barrier Function and Amino Acid Transportation in Weaned Piglets. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2018. [Google Scholar]
- de la Fuente-Nunez, C.; Cesaro, A.; Hancock, R. Antibiotic failure: Beyond antimicrobial resistance. Drug Resist. Updates 2023, 71, 101012. [Google Scholar] [CrossRef]
- Mazhar, M.U.; Naz, S.; Zulfiqar, T.; Khan, J.Z.; Hilal, F.; Ghazanfar, S.; Tipu, M.K. Bacillus subtilis (NMCC-path-14) ameliorates acute phase of arthritis via modulating NF-κB and Nrf-2 signaling in mice model. Inflammopharmacology 2025, 33, 1863–1877. [Google Scholar] [CrossRef]
- Ma, L.; Tao, S.; Song, T.; Lyu, W.; Li, Y.; Wang, W.; Shen, Q.; Ni, Y.; Zhu, J.; Zhao, J.; et al. Clostridium butyricum and carbohydrate active enzymes contribute to the reduced fat deposition in pigs. Imeta 2024, 3, e160. [Google Scholar] [CrossRef]
- Yukino-Iwashita, M.; Nagatomo, Y.; Kawai, A.; Taruoka, A.; Yumita, Y.; Kagami, K.; Yasuda, R.; Toya, T.; Ikegami, Y.; Masaki, N.; et al. Short-Chain Fatty Acids in Gut-Heart Axis: Their Role in the Pathology of Heart Failure. J. Pers. Med. 2022, 12, 1805. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut-heart axis in atherosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef]
- He, X.; Hu, M.; Xu, Y.; Xia, F.; Tan, Y.; Wang, Y.; Xiang, H.; Wu, H.; Ji, T.; Xu, Q.; et al. The gut-brain axis underlying hepatic encephalopathy in liver cirrhosis. Nat. Med. 2025, 31, 627–638. [Google Scholar] [CrossRef]
- Abdulrahim, A.O.; Doddapaneni, N.; Salman, N.; Giridharan, A.; Thomas, J.; Sharma, K.; Abboud, E.; Rochill, K.; Shreelakshmi, B.; Gupta, V.; et al. The gut-heart axis: A review of gut microbiota, dysbiosis, and cardiovascular disease development. Ann. Med. Surg. 2025, 87, 177–191. [Google Scholar] [CrossRef]
- He, Z.; Xiong, H.; Cai, Y.; Chen, W.; Shi, M.; Liu, L.; Wu, K.; Deng, X.; Deng, X.; Chen, T. Clostridium butyricum ameliorates post-gastrectomy insulin resistance by regulating the mTORC1 signaling pathway through the gut-liver axis. Microbiol. Res. 2025, 297, 128154. [Google Scholar] [CrossRef]
- Xia, L.; Wu, Z.; Jin, X.; Xu, H.; Zhang, H.; Zhang, T.; Zhu, J.; Zhang, Z. Application of Lactobacillus reuteri in healthy pig breeding. Swine Ind. Sci. 2025, 42, 76–77. [Google Scholar]
- Zhou, Q.; Wu, F.; Chen, S.; Cen, P.; Yang, Q.; Guan, J.; Cen, L.; Zhang, T.; Zhu, H.; Chen, Z. Lactobacillus reuteri improves function of the intestinal barrier in rats with acute liver failure through Nrf-2/HO-1 pathway. Nutrition 2022, 99–100, 111673. [Google Scholar] [CrossRef]
- Tang, Q.; Yi, H.; Hong, W.; Wu, Q.; Yang, X.; Hu, S.; Xiong, Y.; Wang, L.; Jiang, Z. Comparative Effects of L. plantarum CGMCC 1258 and L. reuteri LR1 on Growth Performance, Antioxidant Function, and Intestinal Immunity in Weaned Pigs. Front. Vet. Sci. 2021, 8, 728849. [Google Scholar] [CrossRef]
- Shu, Z.; Zhang, J.; Zhou, Q.; Peng, Y.; Huang, Y.; Zhou, Y.; Zheng, J.; Zhao, M.; Hu, C.; Lan, S. Effects of inactivated Lactobacillus rhamnosus on growth performance, serum indicators, and colonic microbiota and metabolism of weaned piglets. BMC Vet. Res. 2024, 20, 422. [Google Scholar] [CrossRef]
- Gao, L.M.; Liu, G.Y.; Wang, H.L.; Wassie, T.; Wu, X. Maternal pyrimidine nucleoside supplementation regulates fatty acid, amino acid and glucose metabolism of neonatal piglets. Anim. Nutr. 2022, 11, 309–321. [Google Scholar] [CrossRef]
- Yang, G.; Wang, L.; Yi, H.; Wang, Z.; Yang, X.; Gao, K.; Wen, X.; Jiang, Z. Effects of Lactobacillus reuteri LR1 on Serum Biochemical Indexes and mRNA Expressions of Intestinal Nutrient Transporters of Weaned Piglets. Chin. J. Anim. Nutr. 2018, 30, 4589–4600. [Google Scholar]
- Mi, M.; Shen, Z.; Hu, N.; Zhang, Q.; Wang, B.; Pan, L.; Qin, G.; Bao, N.; Zhao, Y. Effects of diets with different amino acid release characteristics on the gut microbiota and barrier function of weaned pigs. BMC Microbiol. 2023, 23, 18. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, H.; Wang, S.; Zhang, W.; Wang, J.; Tian, H.; Wang, Y.; Ji, H. Fecal Microbiota and Its Correlation with Fatty Acids and Free Amino Acids Metabolism in Piglets After a Lactobacillus Strain Oral Administration. Front. Microbiol. 2019, 10, 785. [Google Scholar] [CrossRef]
- Tie, H.; Wu, P.; Jiang, W.; Liu, Y.; Kuang, S.; Zeng, Y.; Jiang, J.; Tang, L.; Zhou, X.; Feng, L. Dietary nucleotides supplementation affect the physicochemical properties, amino acid and fatty acid constituents, apoptosis and antioxidant mechanisms in grass carp (Ctenopharyngodon idellus) muscle. Aquaculture 2019, 502, 312–325. [Google Scholar] [CrossRef]
- Yoon, K.N.; Lee, H.G.; Yeom, S.J.; Kim, S.S.; Park, J.H.; Song, B.S.; Yi, S.W.; Do, Y.J.; Oh, B.; Oh, S.I.; et al. Lactiplantibacillus argentoratensis AGMB00912 alleviates salmonellosis and modulates gut microbiota in weaned piglets: A pilot study. Sci. Rep. 2024, 14, 15466. [Google Scholar] [CrossRef]
- Gao, T.; Li, R.; Hu, L.; Hu, Q.; Wen, H.; Zhou, R.; Yuan, P.; Zhang, X.; Huang, L.; Zhuo, Y.; et al. Probiotic Lactobacillus rhamnosus GG improves insulin sensitivity and offspring survival via modulation of gut microbiota and serum metabolite in a sow model. J. Anim. Sci. Biotechnol. 2024, 15, 89. [Google Scholar] [CrossRef]
- Wu, G. Metabolism of amino acids in the small intestine: A new perspective on animal nutrition. Feed Ind. 2011, 32, 51–54. [Google Scholar] [CrossRef]
- Hu, S.H.; Feng, Y.Y.; Yang, Y.X.; Ma, H.D.; Zhou, S.X.; Qiao, Y.N.; Zhang, K.H.; Zhang, L.; Huang, L.; Yuan, Y.Y.; et al. Amino acids downregulate SIRT4 to detoxify ammonia through the urea cycle. Nat. Metab. 2023, 5, 626–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, S.; Ma, C.; Song, F.; Li, X.; Wang, H.; Wang, X. Research progress on the intestinal function and transport mechanism of glutamate. Mod. J. Anim. Husb. Vet. Med. 2025, 1, 91–96. [Google Scholar] [CrossRef]
- Ghosh, N.; Mahalanobish, S.; Sil, P.C. Reprogramming of urea cycle in cancer: Mechanism, regulation and prospective therapeutic scopes. Biochem. Pharmacol. 2024, 228, 116326. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Nazemi, M.; Yanes, B.; Martinez, M.L.; Walker, H.J.; Pham, K.; Collins, M.O.; Bard, F.; Rainero, E. The extracellular matrix supports breast cancer cell growth under amino acid starvation by promoting tyrosine catabolism. PLoS Biol. 2024, 22, e3002406. [Google Scholar] [CrossRef]
- Colombero, C.; Remy, D.; Antoine-Bally, S.; Mace, A.S.; Monteiro, P.; Elkhatib, N.; Fournier, M.; Dahmani, A.; Montaudon, E.; Montagnac, G.; et al. mTOR Repression in Response to Amino Acid Starvation Promotes ECM Degradation Through MT1-MMP Endocytosis Arrest. Adv. Sci. 2021, 8, e2101614. [Google Scholar] [CrossRef]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef]
- Liao, X.; Li, X.; Liu, R. Extracellular-matrix mechanics regulate cellular metabolism: A ninja warrior behind mechano-chemo signaling crosstalk. Rev. Endocr. Metab. Disord. 2023, 24, 207–220. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Proportion (%) | 2 Nutrient Component | Content |
|---|---|---|---|
| Corn | 58.06 | NE(Kcal/kg) | 2589 |
| Soybean meal | 16.07 | CP | 22.49 |
| Low protein whey powder | 8.00 | SID Protein | 18.94 |
| Whey powder concentrate | 10.00 | CF | 1.92 |
| Fish meal | 3.00 | EE | 3.56 |
| Soybean oil | 1.00 | SID Lysine | 1.46 |
| Sucrose | 1.00 | SID Threonine | 0.86 |
| 50% choline chloride | 0.20 | SID Methionine | 0.47 |
| NaCl | 0.40 | SID (Methionine + Cysteine) | 0.81 |
| L-lysine | 0.22 | SID Tyrosine | 0.29 |
| DL-methionine | 0.08 | SID Isoleucine | 0.95 |
| L-threonine | 0.01 | SID Valine | 1.00 |
| ZnO | 0.20 | SID Leucine | 1.91 |
| Limestone | 0.44 | SID Phenylalanine | 0.83 |
| Calcium hydrogen phosphate | 0.30 | Total Ca | 0.49 |
| Phytase | 0.02 | STTD Ca | 0.33 |
| 1 Premix | 1.00 | Total P | 0.41 |
| Total | 100 | STTD P | 0.26 |
| Genes | Primer Sequence (5′-3′) | Product Size/bp | Accession Numbers |
|---|---|---|---|
| β-actin | F: CTGCGGCATCCACGAAACT | 380 | XM_003124280 |
| R: AGGGCCGTGATCTCCTTCTG | |||
| COL1A2 | F: GTCTTGGCGGGAACTTTG | 338 | NM_001243655.1 |
| R: ACCGTTGTGACCCCTAATG | |||
| COL3A1 | F: TTTCAAAATCAACACCGACG | 173 | NM_001243297.1 |
| R: CTTGGTTAGGATCAACCCAATA | |||
| COL6A1 | F: ACTCAGAACAACCGGATCG | 177 | XM_021071094.1 |
| R: GCCTTGGCAGGAAATGAC | |||
| FN1 | F: AGCAAGAAGGACAATCGGGG | 325 | XM_003133642.5C |
| R: AGTTGCCACCGTAAGTCTGG | |||
| SLC3A2 | F: AGCAGCGTGACTGTGAAGG | 174 | NM_001012662.2 |
| R: TGGCGGATGTAGGAGAAGAG | |||
| SLC6A14 | F: CCTTGGTCTCGTCTGTGTGA | 136 | NM_001348402.1 |
| R: TCTGTTCCCTCCATAAATCCA | |||
| SLC6A19 | F: TCATCTTCCTCTTCTTCTTCGTG | 155 | XM_003359855.4 |
| R: CTTGACCTTCTGGGATTTGG | |||
| SLC7A1 | F: TCTGGTCCTGGGCTTCATAA | 192 | NM_001012613 |
| R: ACCTTCGTGGCATTGTTCAG | |||
| SLC7A9 | F: GCCTATCAAGGTGCCCATC | 144 | NM_001110171.1 |
| R: AGCGGACGAACAGGAAGTAA | |||
| SLC7A11 | F: TGAATGGTGGTGTGTTTGCT | 101 | NM_014331.3 |
| R: AGTGTGTTTGCGGATGTGAA | |||
| SLC38A2 | F: TCCATTTGATGCCAGTGTTG | 212 | XM_005664159 |
| R: CTCAGAAGAACCAGCGAGGA | |||
| SLC38A9 | F: TCCTGCTGTTCCAGATGATG | 180 | NM_001349383.1 |
| R: TGATTCCTCCTATGTTTGGGTAG |
| Item | CON | LR1 | p-Value |
|---|---|---|---|
| Initial BW (kg) | 5.11 ± 0.09 | 5.09 ± 0.08 | 0.887 |
| Final BW (kg) | 10.91 ± 0.22 b | 11.86 ± 0.32 a | 0.036 |
| ADFI (g/d) | 276.19 ± 11.79 b | 322.22 ± 13.81 a | 0.030 |
| ADG (g/d) | 397.42 ± 3.48 b | 440.89 ± 17.45 a | 0.035 |
| G/F | 0.69 ± 0.03 | 0.73 ± 0.03 | 0.352 |
| Diarrhea ratio | 2.381 ± 1.37 | 1.984 ± 0.79 | 0.815 |
| Item | CON | LR1 | p-Value |
|---|---|---|---|
| Duodenum | |||
| Villus height (μm) | 589.33 ± 38.88 | 474.12 ± 39.26 | 0.064 |
| Crypt depth (μm) | 355.58 ± 46.37 | 477.69 ± 57.71 | 0.130 |
| Villus height–crypt depth | 1.83 ± 0.30 a | 1.05 ± 0.13 b | 0.036 |
| Jejunum | |||
| Villus height (μm) | 457.23 ± 52.61 | 427.13 ± 42.41 | 0.665 |
| Crypt depth (μm) | 320.66 ± 22.87 | 394.25 ± 35.81 | 0.114 |
| Villus height–crypt depth | 1.47 ± 0.19 | 1.13 ± 0.14 | 0.178 |
| Ileum | |||
| Villus height (μm) | 351.10 ± 21.23 b | 448.00 ± 25.79 a | 0.016 |
| Crypt depth (μm) | 301.43 ± 29.39 | 359.17 ± 19.50 | 0.133 |
| Villus height–crypt depth | 1.20 ± 0.09 | 1.27 ± 0.10 | 0.622 |
| Amino Acids | CON | LR1 | p-Value |
|---|---|---|---|
| Alanine | 0.407 ± 0.05 a | 0.196 ± 0.01 b | 0.009 |
| Arginine | 0.029 ± 0.01 | 0.020 ± 0.00 | 0.223 |
| Aspartic Acid | 0.015 ± 0.00 | 0.012 ± 0.00 | 0.381 |
| Asparagine | 0.340 ± 0.01 | 0.030 ± 0.00 | 0.593 |
| β-Alanine | 0.007 ± 0.00 | 0.008 ± 0.00 | 0.327 |
| Cysteine | 0.007 ± 0.00 a | 0.003 ± 0.00 b | 0.020 |
| Glutamic Acid | 0.214 ± 0.04 | 0.123 ± 0.01 | 0.054 |
| Glutamine | 0.639 ± 0.07 a | 0.411 ± 0.03 b | 0.022 |
| Glycine | 0.244 ± 0.03 b | 0.345 ± 0.02 a | 0.009 |
| Histidine | 0.015 ± 0.00 a | 0.009 ± 0.00 b | 0.023 |
| Hydroxyproline | 0.019 ± 0.00 b | 0.027 ± 0.00 a | 0.031 |
| Isoleucine | 0.032 ± 0.00 a | 0.023 ± 0.00 b | 0.008 |
| Lysine | 0.050 ± 0.01 | 0.040 ± 0.00 | 0.329 |
| Methionine | 0.009 ± 0.00 | 0.009 ± 0.00 | 0.908 |
| Ammonia | 0.110 ± 0.04 | 0.054 ± 0.01 | 0.174 |
| Ornithine | 0.021 ± 0.00 | 0.028 ± 0.00 | 0.139 |
| Phenylalanine | 0.020 ± 0.00 | 0.020 ± 0.00 | 0.922 |
| Proline | 0.066 ± 0.01 | 0.058 ± 0.00 | 0.220 |
| Serine | 0.040 ± 0.00 | 0.043 ± 0.00 | 0.486 |
| Taurine | 0.074 ± 0.01 | 0.075 ± 0.01 | 0.979 |
| Threonine | 0.020 ± 0.00 | 0.031 ± 0.00 | 0.055 |
| Tryptophan | 0.021 ± 0.00 | 0.033 ± 0.01 | 0.364 |
| Tyrosine | 0.011 ± 0.00 | 0.013 ± 0.00 | 0.497 |
| Urea | 0.542 ± 0.19 | 0.853 ± 0.12 | 0.200 |
| Valine | 0.052 ± 0.01 | 0.046 ± 0.01 | 0.528 |
| Amino Acids | CON | LR1 | p-Value |
|---|---|---|---|
| Alanine | 4.605 ± 0.11 a | 2.745 ± 0.16 b | <0.001 |
| Anthranilic Acid | 0.024 ± 0.01 | 0.038 ± 0.01 | 0.315 |
| Arginine | 0.047 ± 0.01 | 0.062 ± 0.01 | 0.404 |
| Aspartic Acid | 1.516 ± 0.12 a | 1.041 ± 0.04 b | 0.01 |
| Asparagine | 4.544 ± 0.33 | 3.690 ± 0.22 | 0.057 |
| β-Alanine | 0.187 ± 0.02 | 0.180 ± 0.01 | 0.699 |
| Carnitine | 0.009 ± 0.00 | 0.009 ± 0.00 | 0.938 |
| Cysteine | 0.273 ± 0.04 a | 0.032 ± 0.01 b | 0.002 |
| Cystathionine | 0.016 ± 0.00 | 0.013 ± 0.00 | 0.301 |
| Ethanolamine | 0.298 ± 0.04 | 0.385 ± 0.06 | 0.263 |
| Glutamic Acid | 3.343 ± 0.22 a | 2.703 ± 0.07 b | 0.019 |
| Glutamine | 8.371 ± 0.34 a | 5.447 ± 0.16 b | <0.001 |
| Glycine | 3.624 ± 0.28 | 3.290 ± 0.16 | 0.329 |
| Histidine | 0.484 ± 0.04 a | 0.301 ± 0.02 b | 0.002 |
| Isoleucine | 0.512 ± 0.05 a | 0.341 ± 0.03 b | 0.022 |
| Leucine | 1.135 ± 0.11 a | 0.776 ± 0.06 b | 0.015 |
| Lysine | 1.418 ± 0.12 a | 1.003 ± 0.09 b | 0.019 |
| Methionine | 0.363 ± 0.03 | 0.283 ± 0.02 | 0.071 |
| Ammonia | 4.081 ± 0.37 a | 3.084 ± 0.21 b | 0.041 |
| Ornithine | 0.892 ± 0.09 a | 0.612 ± 0.07 b | 0.029 |
| Phenylalanine | 0.425 ± 0.04 a | 0.258 ± 0.02 b | 0.003 |
| Proline | 1.035 ± 0.10 a | 0.760 ± 0.07 b | 0.045 |
| Serine | 2.053 ± 0.13 a | 1.697 ± 0.06 b | 0.032 |
| Taurine | 0.984 ± 0.06 b | 1.224 ± 0.04 a | 0.007 |
| Threonine | 0.905 ± 0.09 a | 0.683 ± 0.05 b | 0.044 |
| Tryptophan | 0.209 ± 0.04 | 0.171 ± 0.02 | 0.414 |
| Tyrosine | 0.472 ± 0.04 | 0.416 ± 0.04 | 0.329 |
| Urea | 1.569 ± 0.18 | 1.605 ± 0.09 | 0.862 |
| Valine | 1.021 ± 0.10 a | 0.700 ± 0.06 b | 0.019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Wei, Y.; Ruan, S.; Wu, Q.; Xiong, Y.; Wang, L.; Jiang, Z.; Xu, E.; Yi, H. Dietary Supplementation of Lactobacillus reuteri Modulates Amino Acid Metabolism and Extracellular Matrix in the Gut–Liver Axis of Weaned Piglets. Animals 2025, 15, 1567. https://doi.org/10.3390/ani15111567
He Y, Wei Y, Ruan S, Wu Q, Xiong Y, Wang L, Jiang Z, Xu E, Yi H. Dietary Supplementation of Lactobacillus reuteri Modulates Amino Acid Metabolism and Extracellular Matrix in the Gut–Liver Axis of Weaned Piglets. Animals. 2025; 15(11):1567. https://doi.org/10.3390/ani15111567
Chicago/Turabian StyleHe, Yiyi, Yangyang Wei, Shihui Ruan, Qiwen Wu, Yunxia Xiong, Li Wang, Zongyong Jiang, E Xu, and Hongbo Yi. 2025. "Dietary Supplementation of Lactobacillus reuteri Modulates Amino Acid Metabolism and Extracellular Matrix in the Gut–Liver Axis of Weaned Piglets" Animals 15, no. 11: 1567. https://doi.org/10.3390/ani15111567
APA StyleHe, Y., Wei, Y., Ruan, S., Wu, Q., Xiong, Y., Wang, L., Jiang, Z., Xu, E., & Yi, H. (2025). Dietary Supplementation of Lactobacillus reuteri Modulates Amino Acid Metabolism and Extracellular Matrix in the Gut–Liver Axis of Weaned Piglets. Animals, 15(11), 1567. https://doi.org/10.3390/ani15111567