
Environmental Risks and Toxicity of Fipronil and Imidacloprid Used in Pets Ectoparasiticides

 , , , ,
, , , ,  ,
,  , ,
, ,  , and
, and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Field Activities
Water Sampling
2.2. Laboratory Activities
2.2.1. List of Reagents Used in the Laboratory Experiments
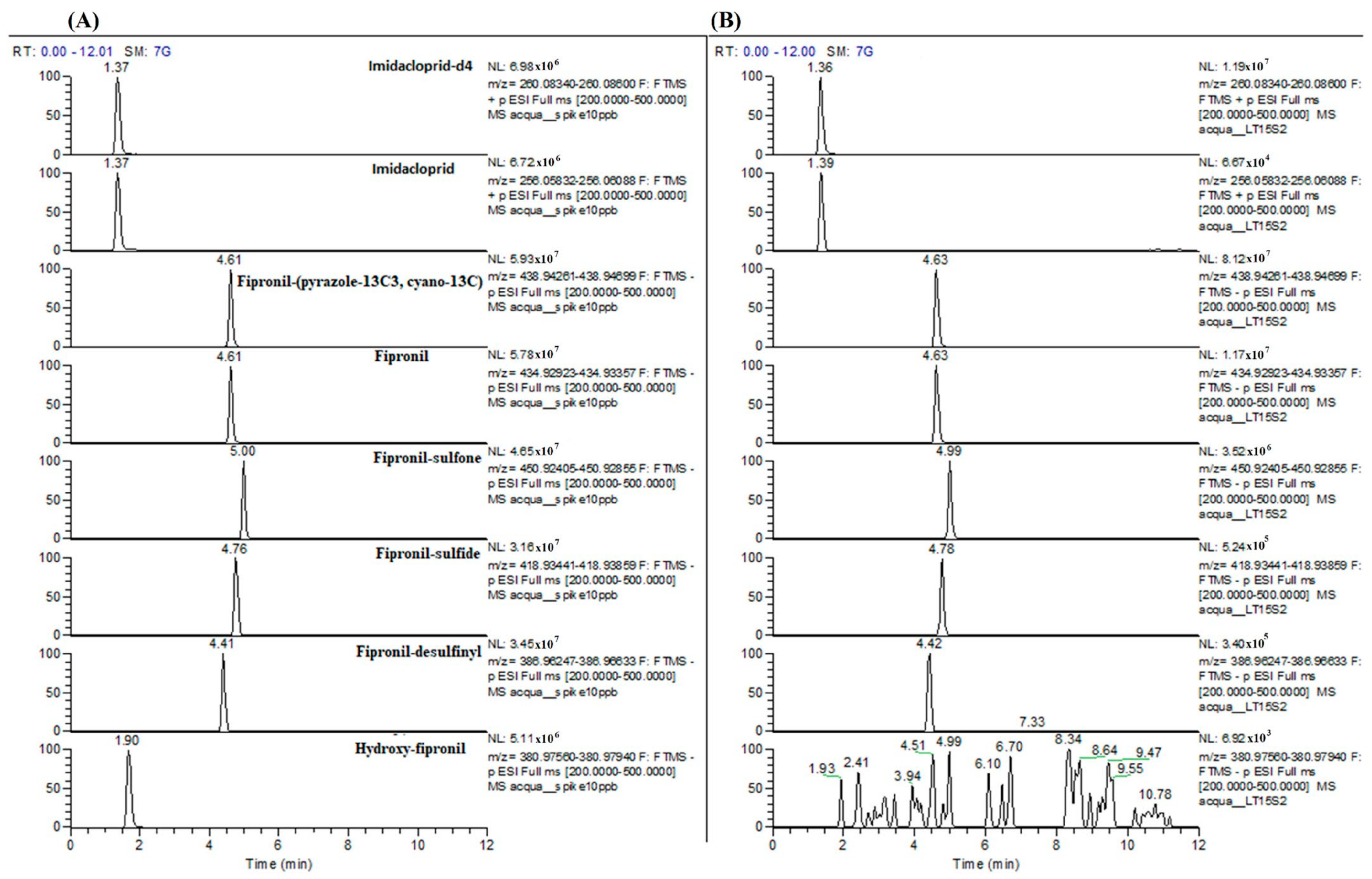
2.2.2. Chemical Quantification of Fipronil and Imidacloprid
Chemical Quantification in Water Samples
Validation of the Analytical Method
2.2.3. Ecotoxicological Test Battery for Marine Species
Phaeodactylum tricornutum Growth Inhibition
Acartia tonsa Toxicity Assays
- Acute toxicity assay: The acute toxicity of the FIP and IMID dilution series was tested in accordance with UNICHIM 2365:2012 [31]. Briefly, six replicates of 2 mL for each assessed concentration were placed in a 24-well plate. Five freshly collected fertilized A. tonsa eggs/nauplii were placed in each well and incubated at 20 ± 1 °C for 48 h in darkness, for a total of 30 eggs/nauplii per sample dilution. The number of immobilized individuals was counted at both 24 and 48 h. Ni2+ was used as the reference toxicant (EC50 0.243 mg Ni2+ L−1, 95% C.I. = 0.201–0.299 mg Ni2+ L−1).
- Chronic toxicity assay: The chronic toxicity of the FIP and IMID concentration series was tested in accordance with UNICHIM 2366:2012 [32]. Briefly, three replicates of 30 mL for each assessed concentration were placed in a 50 mL glass beaker. A total of 10 freshly collected A. tonsa fertilized eggs/nauplii were placed in each well and incubated at 20 ± 1 °C for 7 days, with a photoperiod of 12:10 h (light:darkness). Each sample/dilution was renewed twice during the exposure (every two days). At the end of the exposure period, the number of immobilized individuals was counted. Ni2+ was used as a reference toxicant (EC50 of 0.037 mg Ni2+ L−1, 95% C.I. = 0.028–0.050 mg Ni2+ L−1).
In Vitro Biochemical Responses of Mytilus galloprovincialis
2.2.4. In Vitro Cell-Based Approaches
Cell Viability Assay
Intracellular ROS Detection
Detection of Lipid Peroxidation (LPO) Levels
2.3. Statistical Analyses
3. Results
3.1. Fipronil and Imidacloprid Concentrations in Wastewater
3.2. Validation of the Analytical Method for Fipronil and Imidacloprid
3.3. Ecotoxicological Test Battery Responses
3.3.1. Phaeodactylum tricornutum Growth Inhibition
3.3.2. Acartia tonsa Toxicity Assays
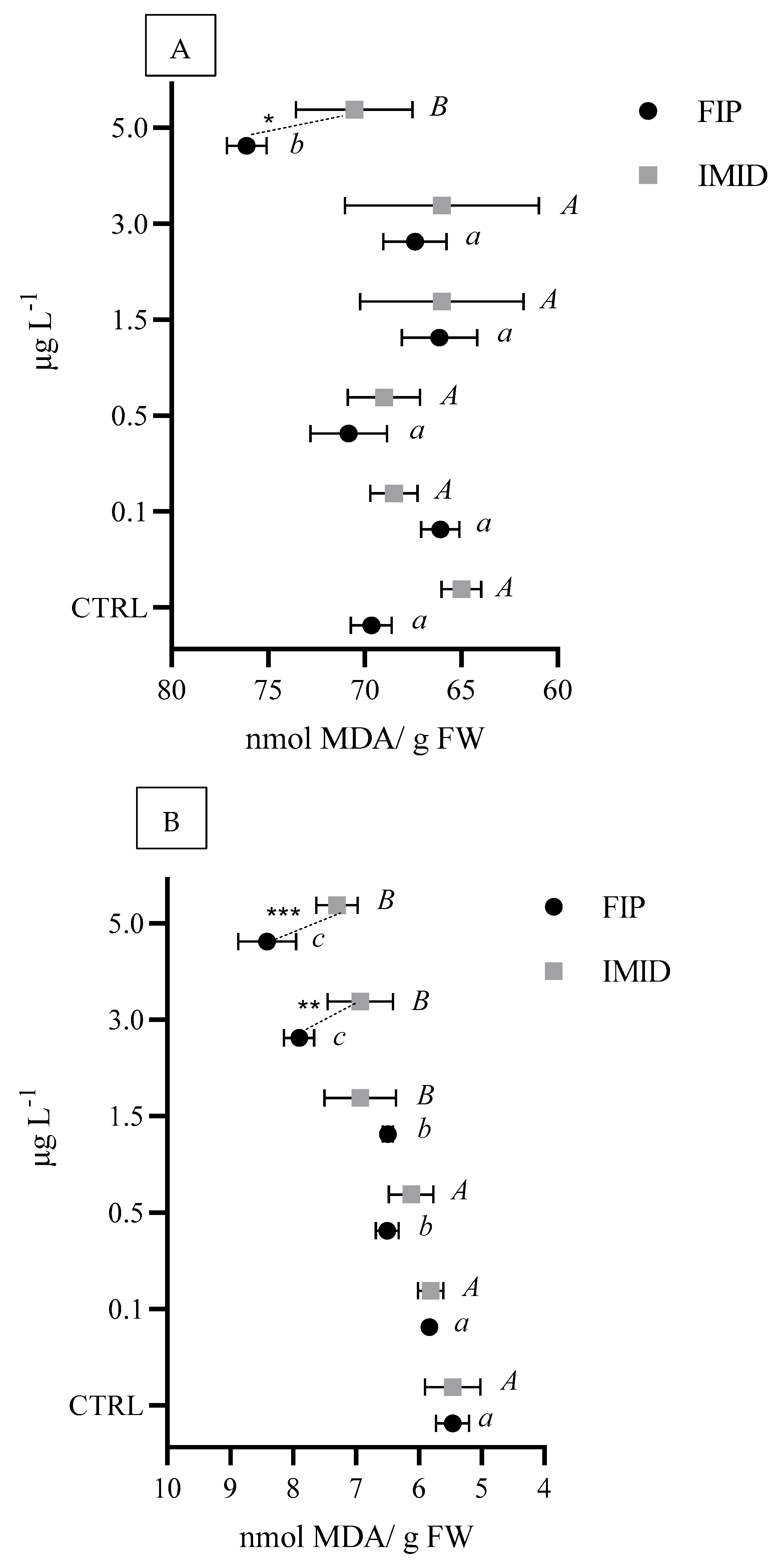
3.3.3. Biochemical Markers of Mytilus galloprovincialis
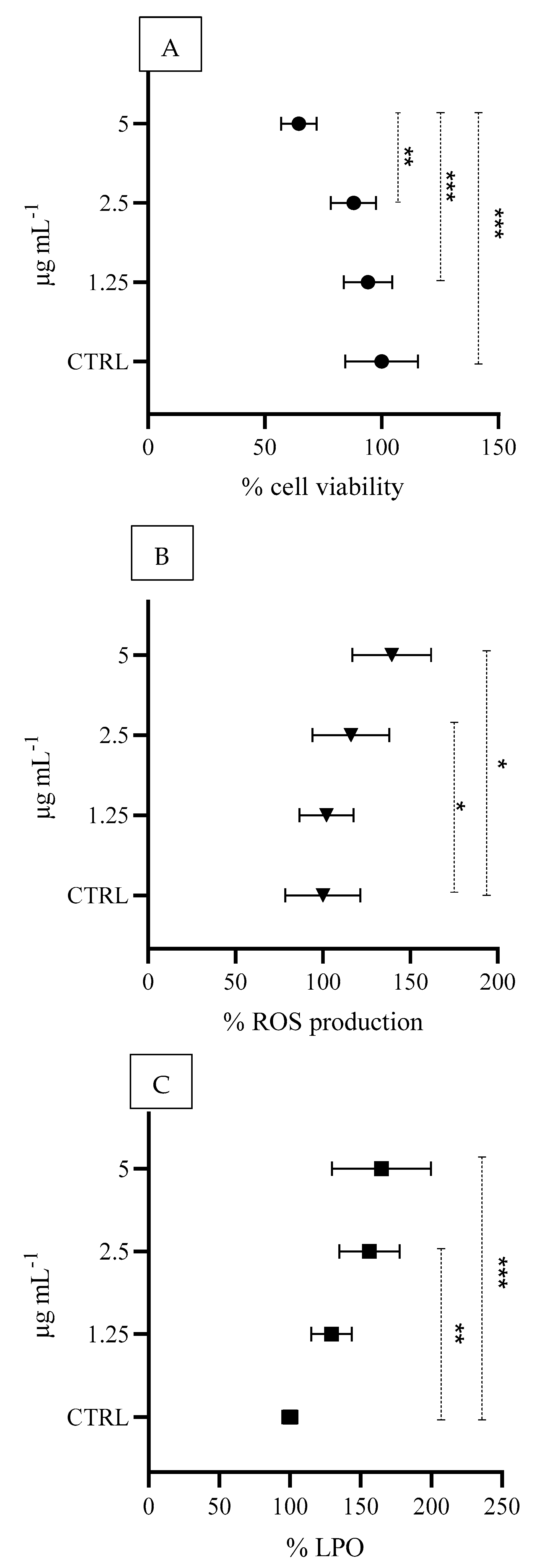
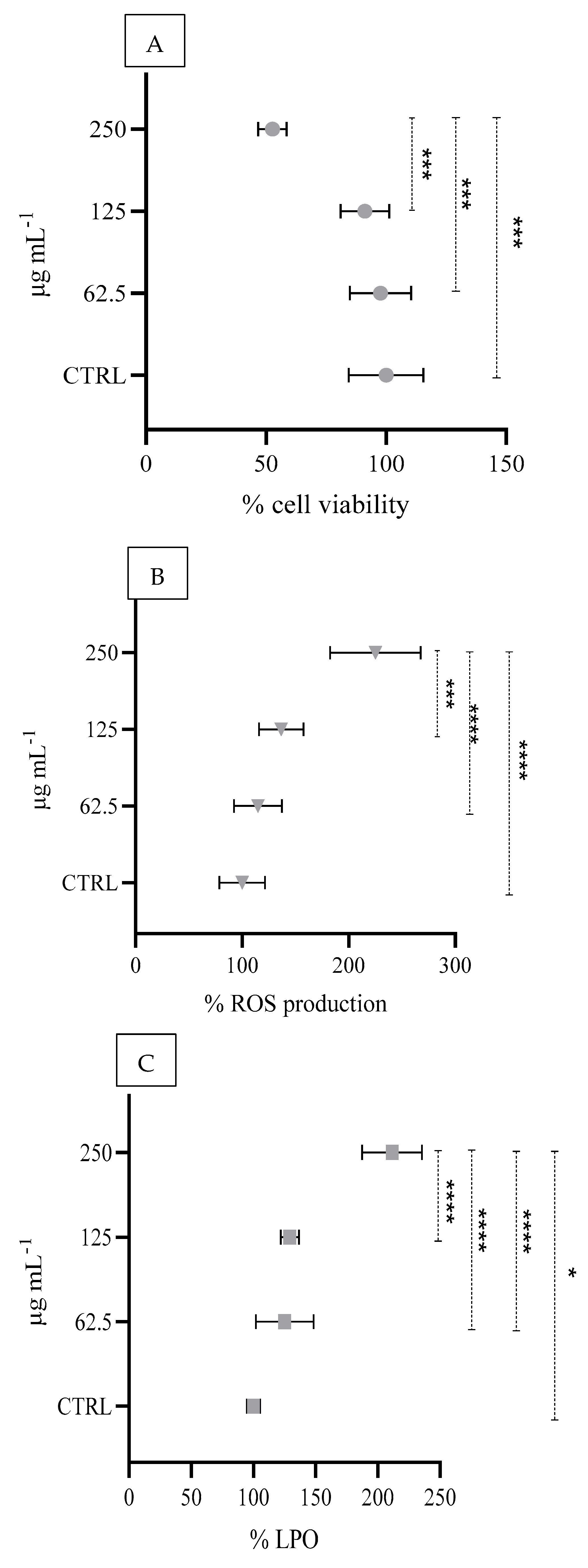
3.4. In Vitro Cell-Based Responses
4. Discussion
4.1. Fipronil and Imidacloprid Concentrations in Wastewater
4.2. Ecotoxicological Assessment
4.3. In Vitro Cell-Based Toxicity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Medicines Agency (EMA)/Committee for Medicinal Products for Veterinary Use (CVMP). Reflection Paper on the Environmental Risk Assessment of Ectoparasiticidal Veterinary Medicinal Products Used in Cats and Dogs. 2023. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/reflection-paper-environmental-risk-assessment-ectoparasiticidal-veterinary-medicinal-products-used-cats-and-dogs_en.pdf (accessed on 2 April 2025).
- Grand View Research. Animal Health Market Size, Share & Trends Analysis Report by Product (Biologics, Pharmaceuticals), by Animal Type (Companion, Production), by Distribution Channel, by End-Use, By Region, and Segment Forecasts, 2025–2030. 2023. Available online: https://www.grandviewresearch.com (accessed on 2 April 2025).
- European Scientific Counsel Companion Animal Parasites (ESCCAP). Control of Ectoparasites in Dogs and Cats, ESCCAP Guideline 03, 7th ed.; ESCCAP: Malvern, UK, 2022; pp. 1–33. Available online: https://www.esccap.org/link-document/63/ (accessed on 2 April 2025).
- Pfister, K.; Armstrong, R. Systemically and cutaneously distributed ectoparasiticides: A review of the efficacy against ticks and fleas on dogs. Parasites Vectors 2016, 9, 436. [Google Scholar] [CrossRef] [PubMed]
- Lavan, R.; Armstrong, R.; Normile, D.; Vaala, W. Adherence to veterinary recommendations for ectoparasiticides purchased by cat owners in the USA. Parasites Vectors 2020, 13, 541. [Google Scholar] [CrossRef] [PubMed]
- Yuskiv, I.D.; Tishyn, O.L.; Yuskiv, L.L. Evaluation of efficacy of a drug based on fipronil, ivermectin, and pyriproxyfen against ecto- and endoparasitic invasions of dogs and cats. Regul. Mech. Biosyst. 2024, 15, 113–118. [Google Scholar] [CrossRef]
- Ziliotto, L.; Luna, S.P.L.; Filho, D.A.A.; Resende, L.O.; Aun, A.G.; Braz, M.G. Genotoxicity assessment of fipronil (Frontline Plus®). Pesq. Vet. Bras. 2017, 37, 257–260. [Google Scholar] [CrossRef]
- Cravedi, J.P.; Delous, G.; Zalko, D.; Viguié, C.; Debrauwer, L. Chemosphere disposition of fipronil in rats. Chemosphere 2013, 93, 2276–2283. [Google Scholar] [CrossRef]
- Singh, N.S.; Sharma, R.; Singh, S.K.; Singh, D.K. A comprehensive review of environmental fate and degradation of fipronil and its toxic metabolites. Environ. Res. 2021, 199, 111316. [Google Scholar] [CrossRef]
- Zhao, X.; Yeh, J.Z.; Salgado, V.L.; Narahashi, T. Sulfone metabolite of fipronil blocks γ-aminobutyric acid- and glutamate-activated chloride channels in mammalian and insect neurons. J. Pharmacol. Exp. Ther. 2005, 314, 363–373. [Google Scholar] [CrossRef]
- Wan, Y.; Han, Q.; Wang, Y.; He, Z. Five degradates of imidacloprid in source water, treated water, and tap water in Wuhan, central China. Sci. Total Environ. 2020, 741, 140227. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). Environmental Impact Assessment (EIA) for Veterinary Medicinal Products (VMPs)—Phase I. 2000. Available online: https://www.ema.europa.eu/en/environmental-impact-assessment-veterinary-medicinal-products-support-vich-guidelines-gl6-gl38-scientific-guideline (accessed on 2 April 2025).
- Koslowski, S.; Latapy, C.; Auvray, P.; Blondel, M.; Meijer, L. Long-term fipronil treatment induces hyperactivity in female mice. Int. J. Environ. Res. Public Health 2020, 17, 1579. [Google Scholar] [CrossRef]
- Ferreira, T.P.; Bauerfeldt, G.F.; Castro, R.N.; Magalhães, V.S.; Alves, M.C.; Scott, F.B.; Cid, Y.P. Determination of fipronil and fipronil-sulfone in surface waters of the Guandu River Basin by high-performance liquid chromatography with mass spectrometry. Bull. Environ. Contam. Toxicol. 2022, 108, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Perkins, R.; Barron, L.; Glauser, G.; Whitehead, M.; Woodward, G.; Goulson, D. Down-the-drain pathways for fipronil and imidacloprid applied as spot-on parasiticides to dogs: Estimating aquatic pollution. Sci. Total Environ. 2024, 917, 170175. [Google Scholar] [CrossRef] [PubMed]
- Perkins, R.; Whitehead, M.; Civil, W.; Goulson, D. Potential role of veterinary flea products in widespread pesticide contamination of English rivers. Sci. Total Environ. 2021, 755, 143560. [Google Scholar] [CrossRef] [PubMed]
- de la Casa-Resino, I.; Villegas Grondona, J.M.; Puig Infante, A.; Haro Castuera, A.; Bartolom’e Rodríguez, A.; Herrera Hern’andez, E.M.; Rubio, C.F.; Caixach, J.; Carapeto García, R. Residues of Imidacloprid in Surface Water Treatment Plants and Watercourses in Spain: Are Veterinary Medicinal Products (VMPs) a Significant Source of Emission? Spanish Agency of Medicines and Medical Devices (AEMPS). 2022. Available online: https://www.aemps.gob.es/publicaciones/publica/docs/Imidacloprid.pdf (accessed on 6 May 2025).
- Finckh, S.; Beckers, L.-M.; Busch, W.; Carmona, E.; Dulio, V.; Kramer, L.; Krauss, M.; Posthuma, L.; Schulze, T.; Slootweg, J.; et al. A risk based assessment approach for chemical mixtures from wastewater treatment plant effluents. Environ. Int. 2022, 164, 107234. [Google Scholar] [CrossRef]
- Budd, R.; Teerlink, J.; Alaimo, C.; Wong, L.; Young, T.M. Sub-sewershed monitoring to elucidate down-the-drain pesticide sources. Environ. Sci. Technol. 2023, 57, 5404–5413. [Google Scholar] [CrossRef]
- UNESCO. Wastewater: The Untapped Resource. The United Nations World Water Development Report 2017. United Nations Educational, Scientific and Cultural Organization (UNESCO); UNESCO: Paris, France, 2017. [Google Scholar]
- Burdon, F.J.; Munz, N.A.; Reyes, M.; Focks, A.; Joss, A.; Räsänen, K.; Altermatt, F. Agricultural and urban land uses drive in-stream concentrations and fluxes of pesticides at the catchment scale. Sci. Total Environ. 2019, 682, 737–746. [Google Scholar]
- Giannelli, A.; Schnyder, M.; Wright, I.; Charlier, J. Control of companion animal parasites and impact on One Health. One Health 2024, 18, 100679. [Google Scholar] [CrossRef]
- Testa, C.; Salis, S.; Rubattu, N.; Roncada, P.; Miniero, R.; Brambilla, G. Occurrence of fipronil in residential house dust in the presence and absence of pets: A hint for a comprehensive toxicological assessment. J. Environ. Sci. Health Part B 2019, 54, 441–448. [Google Scholar] [CrossRef]
- Dos Santos, G.C.M.; Scott, F.B.; Campos, D.R.; Magalhaes, V.D.S.; Borges, D.A.; Miranda, F.R.; Alves, M.C.C.; Pereira, G.A.; Moreira, L.O.; Lima, E.A.S.; et al. Oral pharmacokinetic profile of fipronil and efficacy against flea and tick in dogs. J. Vet. Pharmacol. Ther. 2022, 45, 23–33. [Google Scholar] [CrossRef]
- Anthe, M.; Valles-Ebeling, B.; Achtenhagen, J.; Arenz-Leufen, M.; Atkinson, J.; Starp, M.; Corsing, C. Development of an aquatic exposure assessment model for imidacloprid in sewage treatment plant discharges arising from use of veterinary medicinal products. Environ. Sci. Eur. 2020, 32, 147. [Google Scholar] [CrossRef]
- European Commission. European Commission SANTE/11312/2021: Analytical Quality Control and Method Validation Procedures for Pesticide Residues Analysis in Food and Feed. Available online: https://food.ec.europa.eu/system/files/2023-11/pesticides_mrl_guidelines_wrkdoc_2021-11312.pdf (accessed on 2 April 2025).
- ISO 10253:2016; Water Quality—Marine Algal Growth Inhibition Test with Skeletonema sp. and Phaeodactylum tricornutum. ISO: Geneva, Switzerland, 2016.
- ASTM E1218-04(2012); Standard Guide for Conducting Static Toxicity Tests with Microalgae. ASTM: West Conshohocken, PA, USA, 2012.
- Rodrigues, L.H.R.; Arenzon, A.; Raya-Rodriguez, M.T.; Fontoura, N.F. Algal density assessed by spectrophotometry: A calibration curve for the unicellular algae Pseudokirchneriella subcapitata. J. Environ. Chem. Ecotoxicol. 2011, 3, 225–228. [Google Scholar]
- UNICHIM M.U. 2365:2012; Qualità dell’acqua: Determinazione Dell’inibizione Della Mobilità di Naupli di Acartia tonsa Dana (Crustacea: Copepoda) Dopo 24 h e 48 h di Esposizione. Associazione per l‘Unificazione del Settore dell’Industria Chimica: Milan, Italy, 2012; p. 22.
- UNICHIM M.U. 2366:2012; Qualità Dell’acqua: Determinazione Dell’inibizione Della Mobilità di Naupli di Acartia tonsa Dana (Crustacea: Copepoda) Dopo 7 Giorni di Esposizione. Associazione per l‘Unificazione del Settore dell’Industria Chimica: Milan, Italy, 2012; p. 24.
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Giannessi, J.; De Marchi, L.; Meucci, V.; Intorre, L.; Monni, G.; Baratti, M.; Pretti, C. Subcellular tissue-specific responses of Mytilus galloprovincialis to fluoroquinolone antibiotics. Environ. Toxicol. Pharmacol. 2023, 104, 104306. [Google Scholar] [CrossRef]
- Liu, S.; Wang, T.; Lu, J.; Li, Z. Seawater quality criteria derivation and ecological risk assessment for the neonicotinoid insecticide imidacloprid in China. Mar. Pollut. Bull. 2023, 190, 114871. [Google Scholar] [CrossRef]
- Dulio, V.; Alygizakis, N.; Ng, K.; Schymanski, E.L.; Andres, S.; Vorkamp, K.; Hollender, J.; Finckh, S.; Aalizadeh, R.; Ahrens, L.; et al. Beyond target chemicals: Updating the NORMAN prioritisation scheme to support the EU chemicals strategy with semi-quantitative suspect/non-target screening data. Environ. Sci. Eur. 2024, 36, 113. [Google Scholar] [CrossRef]
- Arslan, P.; Günal, A.Ç. Does fipronil affect on aquatic organisms? Physiological, biochemical, and histopathological alterations of non-target freshwater mussel species. Water 2023, 15, 334. [Google Scholar] [CrossRef]
- Clasen, V.L.; Loro, R.; Cattaneo, B.; Moraes, T.; Lópes, L.A.; de Avila, R.; Zanella, G.B.; Reimche, B.; Baldisserotto, B. Effects of the commercial formulation containing fipronil on the non-target organism Cyprinus carpio: Implications for rice-fish cultivation. Ecotoxicol. Environ. Saf. 2012, 77, 45–51. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.A.; Raimundo, C.C.M.; dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Yoloğlu, E. Alterations in some biochemical responses of freshwater mussels in acute imidacloprid exposure. Adiyaman Univ. J. Sci. 2019, 9, 213–229. [Google Scholar] [CrossRef]
- Pagano, M.; Stara, A.; Aliko, V.; Faggio, C. Impact of neonicotinoids to aquatic invertebrates—In vitro studies on Mytilus galloprovincialis: A review. J. Mar. Sci. Eng. 2020, 8, 801. [Google Scholar] [CrossRef]
- Kuchovská, E.; Morin, B.; López-Cabeza, R.; Barré, M.; Gouffier, C.; Bláhová, L.; Cachot, J.; Bláha, L.; Gonzalez, P. Comparison of imidacloprid, propiconazole, and nanopropiconazole effects on the development, behavior, and gene expression biomarkers of the Pacific oyster (Magallana gigas). Sci. Total Environ. 2021, 764, 142921. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.I.; Vargas, M. Toxicity and hazard assessment of fipronil to Daphnia pulex. Ecotoxicol. Environ. Saf. 2005, 62, 11–16. [Google Scholar] [CrossRef]
- Wu, C.H.; Lu, C.W.; Hsu, T.H.; Wu, W.J.; Wang, S.E. Neurotoxicity of fipronil affects sensory and motor systems in zebrafish. Pestic. Biochem. Physiol. 2021, 177, 104896. [Google Scholar] [CrossRef]
- Konwick, B.J.; Fisk, A.T.; Garrison, A.W.; Avants, J.K.; Black, M.C. Acute enantioselective toxicity of fipronil and its desulfinyl photoproduct to Ceriodaphnia dubia. Environ. Toxicol. Chem. 2005, 24, 2350–2355. [Google Scholar] [CrossRef]
- Qin, G.; Presley, S.M.; Anderson, T.A.; Gao, W.; Maul, J.D. Effects of predator cues on pesticide toxicity: Toward an understanding of the mechanism of the interaction. Environ. Toxicol. Chem. 2011, 30, 1926–1934. [Google Scholar] [CrossRef]
- Hayasaka, D.; Korenaga, T.; Suzuki, K.; Sánchez-Bayo, F.; Goka, K. Differences in susceptibility of five cladoceran species to two systemic insecticides, imidacloprid and fipronil. Ecotoxicology 2012, 21, 421–427. [Google Scholar] [CrossRef]
- U.S. EPA—United States Environmental Protection Agency. US EPA Database. 2018. Available online: https://www.epa.gov (accessed on 2 April 2025).
- Stevens, M.M.; Burdett, A.S.; Mudford, E.M.; Helliwell, S.; Doran, G. The acute toxicity of fipronil to two non-target invertebrates associated with mosquito breeding sites in Australia. Acta Trop. 2011, 117, 125–130. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Conclusion on the peer review of the pesticide risk assessment for bees for the active substance imidacloprid considering all uses other than seed treatments and granules. EFSA J. 2015, 13, 4211. [Google Scholar]
- European Food Safety Authority (EFSA). Peer review of the pesticide risk assessment for the active substance imidacloprid in light of confirmatory data submitted. EFSA J. 2016, 14, e04607. [Google Scholar]
- Merga, L.B.; Van den Brink, P.J. Ecological effects of imidacloprid on a tropical freshwater ecosystem and subsequent recovery dynamics. Sci. Total Environ. 2021, 784, 147167. [Google Scholar] [CrossRef] [PubMed]
- Picone, M.; Distefano, G.G.; Marchetto, D.; Russo, M.; Baccichet, M.; Zangrando, R.; Gambaro, A.; Ghirardini, A.V. Inhibition of larval development of marine copepods Acartia tonsa by neonicotinoids. Toxics 2022, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Smit, C.E.; Posthuma-Doodeman, C.J.A.M.; van Vlaardingen, P.L.A.; de Jong, F.M.W. Ecotoxicity of imidacloprid to aquatic organisms: Derivation of water quality standards for peak and long-term exposure. Hum. Ecol. Risk Assess. 2015, 21, 1608–1630. [Google Scholar] [CrossRef]
- Leri, M.; Vasarri, M.; Barletta, E.; Schiavone, N.; Bergonzi, M.C.; Bucciantini, M.; Degl’Innocenti, D. The protective role of oleuropein aglycone against pesticide-induced toxicity in a human keratinocytes cell model. Int. J. Mol. Sci. 2023, 24, 14553. [Google Scholar] [CrossRef]
- Silva, A.M.; Martins-Gomes, C.; Silva, T.L.; Coutinho, T.E.; Souto, E.B.; Andreani, T. In vitro assessment of pesticides toxicity and data correlation with pesticides physicochemical properties for prediction of toxicity in gastrointestinal and skin contact exposure. Toxics 2022, 10, 378. [Google Scholar] [CrossRef]
- Raabe, H.A.; Costin, G.E.; Allen, D.G.; Lowit, A.; Corvaro, M.; O’Dell, L.; Breeden-Alemi, J.; Page, K.; Perron, M.; Silva, T.F.; et al. Human relevance of in vivo and in vitro skin irritation tests for hazard classification of pesticides. Cutan. Ocul. Toxicol. 2024, 44, 1–21. [Google Scholar] [CrossRef]
- Vidau, C.; González-Polo, R.A.; Niso-Santano, M.; Gómez-Sánchez, R.; Bravo-San Pedro, J.M.; Pizarro-Estrella, E.; Blasco, R.; Brunet, J.-L.; Belzunces, L.P.; Fuentes, J.M. Fipronil is a powerful uncoupler of oxidative phosphorylation that triggers apoptosis in human neuronal cell line SHSY5Y. Neurotoxicology 2011, 32, 935–943. [Google Scholar] [CrossRef]
- Qu, H.; Ma, R.; Liu, D.; Wang, P.; Huang, L.; Qiu, X.; Zhou, Z. Enantioselective toxicity, degradation and transformation of the chiral insecticide fipronil in two algae cultures. Environ. Toxicol. Chem. 2022, 41, 1234–1245. [Google Scholar]
- Sardo, A.M.; Soares, A.M. Comparative toxicity of imidacloprid and its transformation product 6-chloronicotinic acid to non-target aquatic organisms: Microalgae Desmodesmus subspicatus and amphipod Gammarus fossarum. Pest. Biochem. Physiol. 2010, 98, 129–137. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| µg L−1 | ||||||
|---|---|---|---|---|---|---|
| Sample | IMID | FIP | FIP-Sulfone | FIP-Sulfide | FIP-Desulfinil | Hydroxy-FIP |
| T0 | n.d. | <LOQ | 0.05 | n.d. | n.d. | n.d. |
| T7 | 0.36 | 0.42 | 0.10 | <LOQ | <LOQ | n.d. |
| T15 | 0.06 | 1.36 | 0.40 | 0.07 | 0.05 | n.d. |
| T30 | 0.08 | 0.16 | 0.31 | <LOQ | 0.06 | n.d. |
| T60 | 0.20 | 0.05 | 0.13 | <LOQ | <LOQ | n.d. |
| mean | 0.18 | 0.50 | 0.20 | - | 0.06 | - |
| sd | 0.12 | 0.52 | 0.13 | - | 0.01 | - |
| Analytes | Validation Levels (µg L−1) | Recovery * % | CV * Intraday % | CV * Interday % | Linearity R2 | Matrix Effect % |
|---|---|---|---|---|---|---|
| IMID | LOQ 0.05 1.0 | 99 100 | 5 4 | 7 6 | 0.9997 | 98 |
| FIP | LOQ 0.05 1.0 | 99 100 | 5 3 | 7 5 | 0.9994 | 98 |
| FIP-sulfide | LOQ 0.05 1.0 | 98 101 | 5 3 | 7 6 | 0.9991 | 97 |
| FIP-sulfone | LOQ 0.05 1.0 | 97 99 | 5 3 | 8 6 | 0.9993 | 97 |
| FIP-desulfinyl | LOQ 0.05 1.0 | 97 100 | 6 4 | 9 7 | 0.9995 | 96 |
| Hydroxy-FIP | LOQ 0.05 1.0 | 96 98 | 9 6 | 11 8 | 0.9990 | 94 |
| ECx | µg L−1 (95% C.I.) | |
|---|---|---|
| FIP | IMID | |
| EC10 | 1.70 (0.06–6.59) | 2.59 (0.80–6.33) |
| EC20 | 2.80 (0.436–8.51) | 7.60 (3.12–15.8) |
| EC50 | - | 47.50 (25.30–90.10) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marchi, L.; Oliva, M.; Nobile, M.; Carere, M.; Chiesa, L.M.; Degl’Innocenti, D.; Lacchetti, I.; Mancini, L.; Meucci, V.; Pretti, C.; et al. Environmental Risks and Toxicity of Fipronil and Imidacloprid Used in Pets Ectoparasiticides. Animals 2025, 15, 1533. https://doi.org/10.3390/ani15111533
De Marchi L, Oliva M, Nobile M, Carere M, Chiesa LM, Degl’Innocenti D, Lacchetti I, Mancini L, Meucci V, Pretti C, et al. Environmental Risks and Toxicity of Fipronil and Imidacloprid Used in Pets Ectoparasiticides. Animals. 2025; 15(11):1533. https://doi.org/10.3390/ani15111533
Chicago/Turabian StyleDe Marchi, Lucia, Matteo Oliva, Maria Nobile, Mario Carere, Luca Maria Chiesa, Donatella Degl’Innocenti, Ines Lacchetti, Laura Mancini, Valentina Meucci, Carlo Pretti, and et al. 2025. "Environmental Risks and Toxicity of Fipronil and Imidacloprid Used in Pets Ectoparasiticides" Animals 15, no. 11: 1533. https://doi.org/10.3390/ani15111533
APA StyleDe Marchi, L., Oliva, M., Nobile, M., Carere, M., Chiesa, L. M., Degl’Innocenti, D., Lacchetti, I., Mancini, L., Meucci, V., Pretti, C., Vasarri, M., Villa, R. E., & Intorre, L. (2025). Environmental Risks and Toxicity of Fipronil and Imidacloprid Used in Pets Ectoparasiticides. Animals, 15(11), 1533. https://doi.org/10.3390/ani15111533