Comparative Analysis of Proteomic Characteristics in Seminal Plasma Between Horses and Donkeys



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
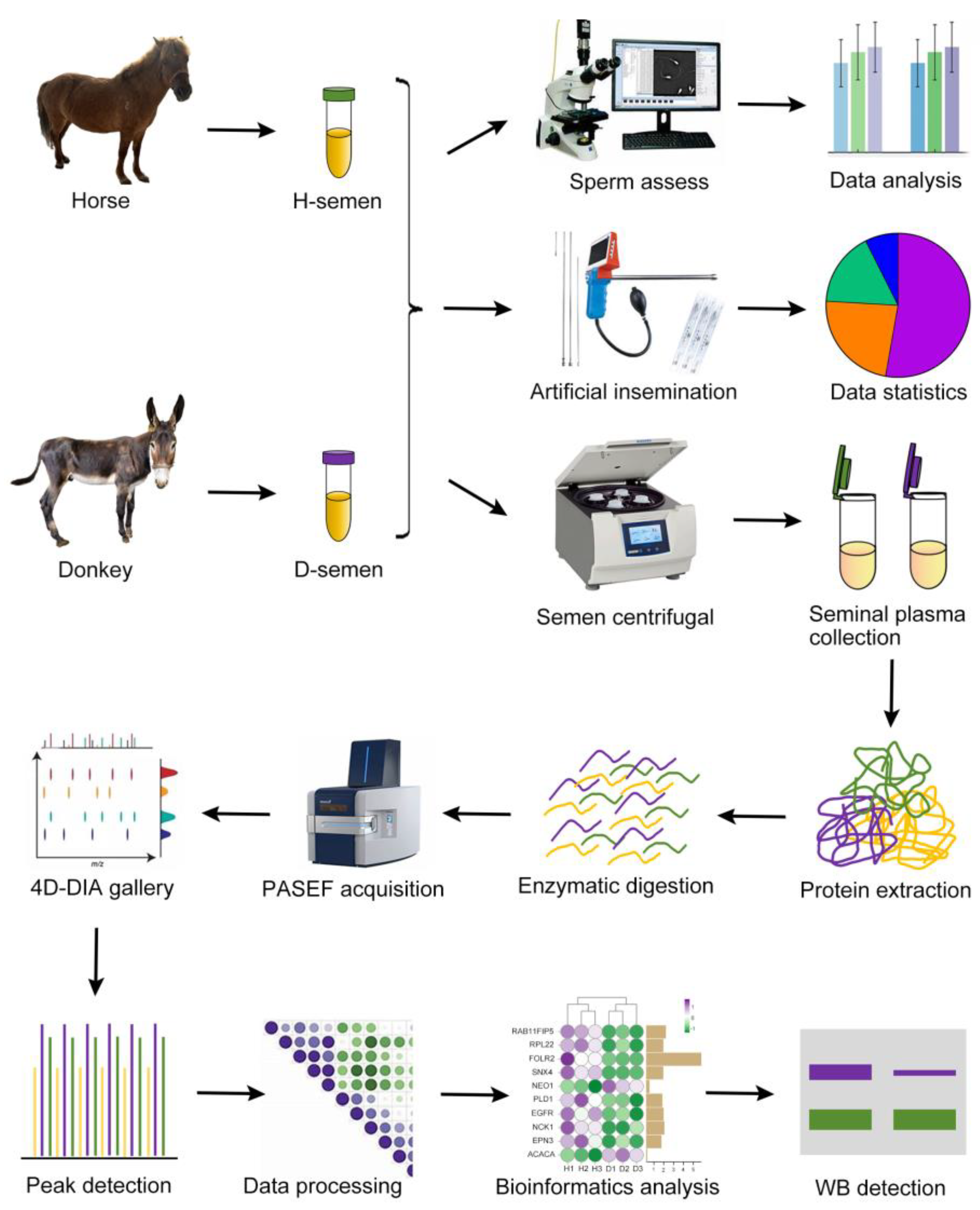
2.1. Experimental Design
2.2. Trial Animals
2.3. Sperm Assessment
2.4. Artificial Insemination
2.5. Seminal Plasma Preparation
2.6. Protein Extraction and Digestion
2.7. LC-MS/MS Detection
2.8. Database Search
2.9. Bioinformatics Analysis
2.10. PPI and Correlation Matrix
2.11. Western Blot Verification
2.12. Statistical Analysis
3. Results
3.1. Evaluation of Horse and Donkey Sperm Quality and Gestation
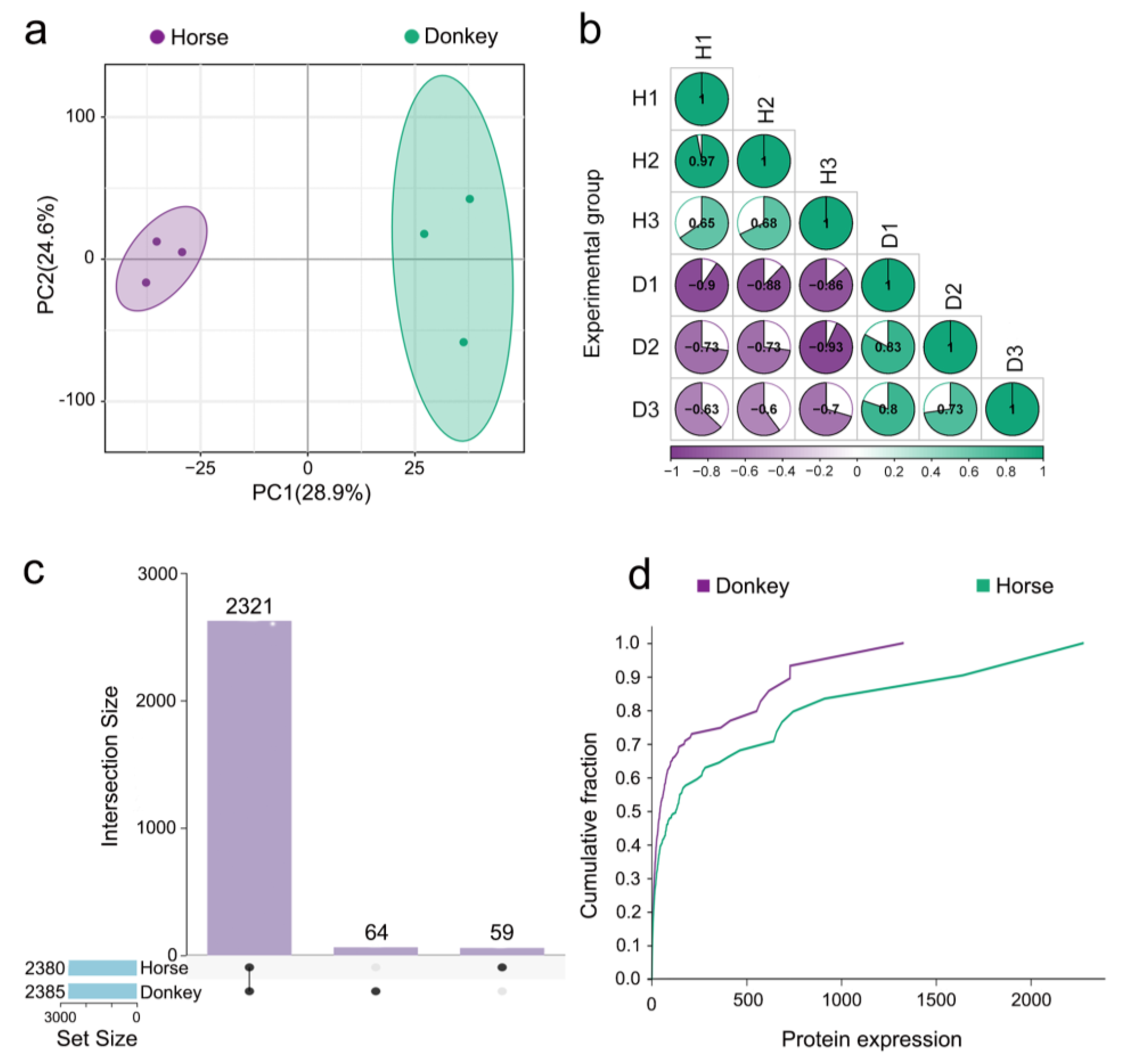
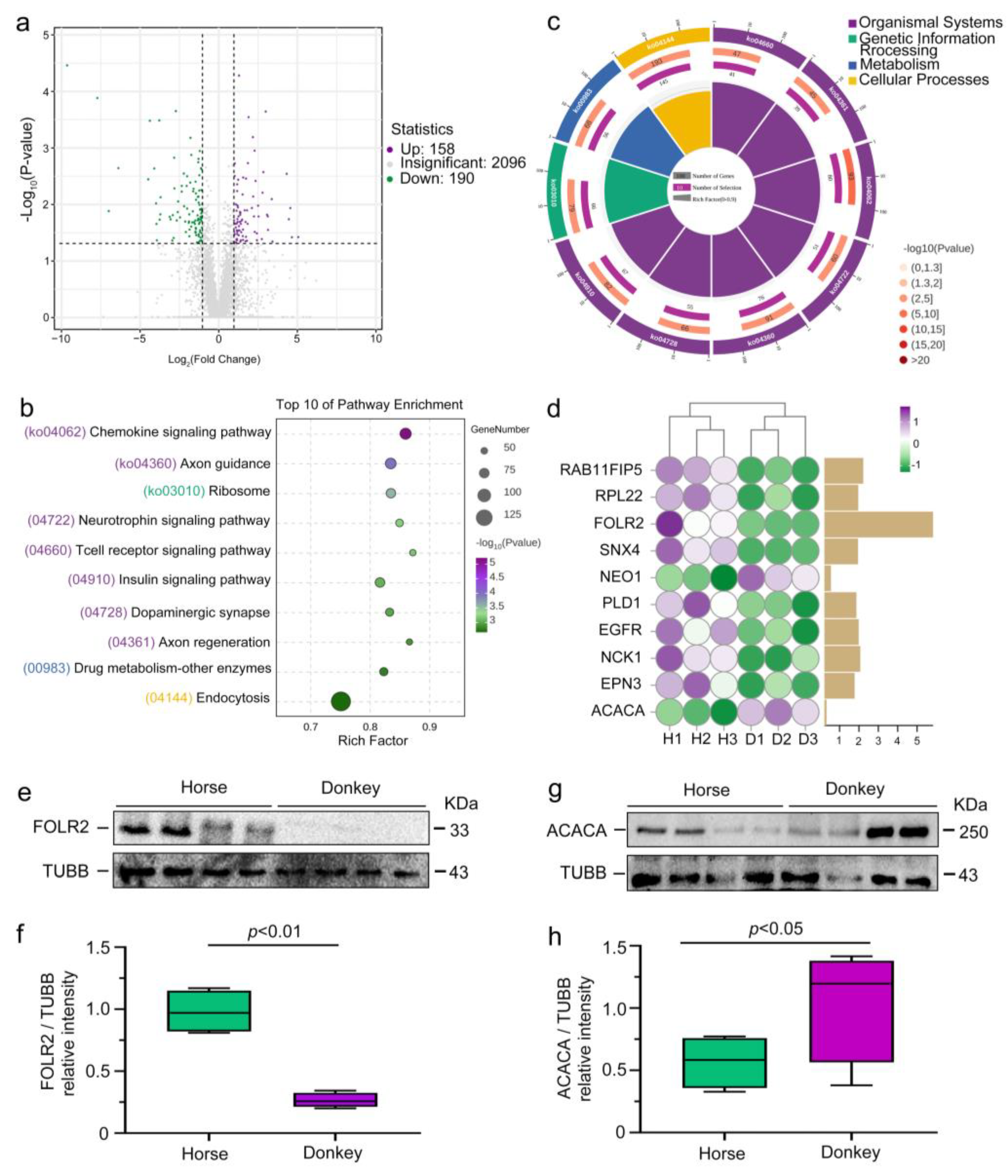
3.2. Display of the Seminal Plasma Proteome in Horses and Donkeys
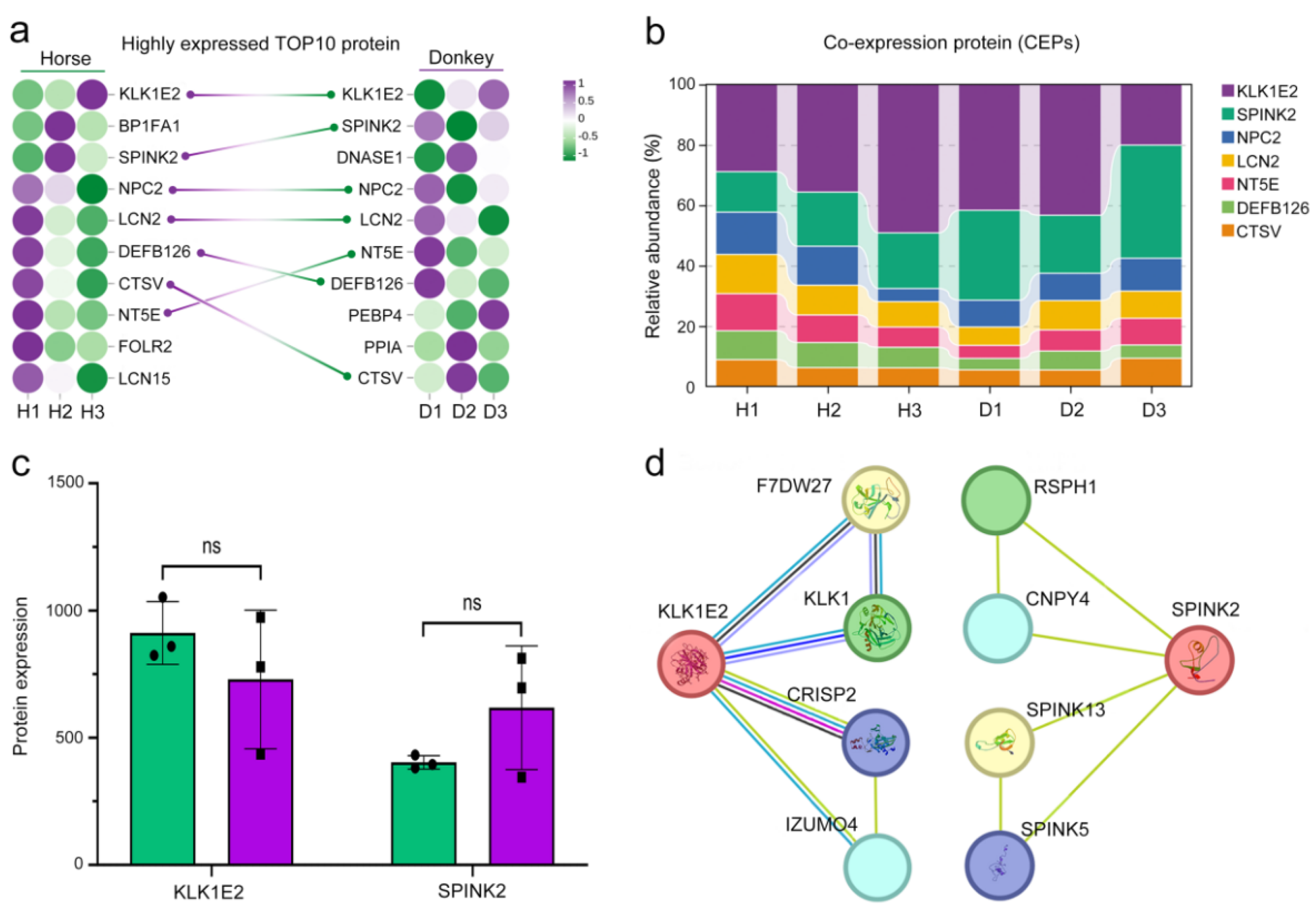
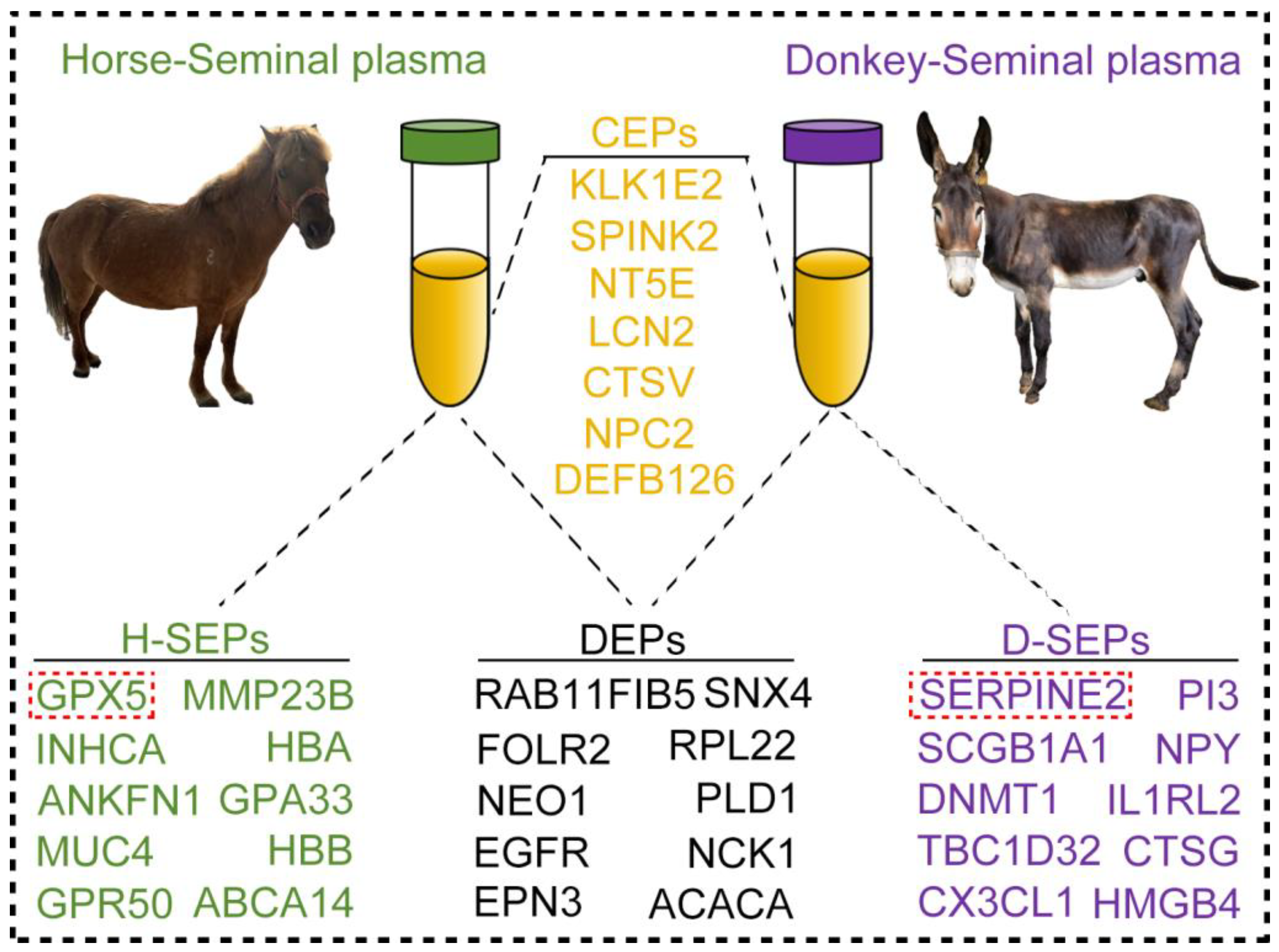
3.3. Presentation of Seminal Plasma CEPs in Horses and Donkeys
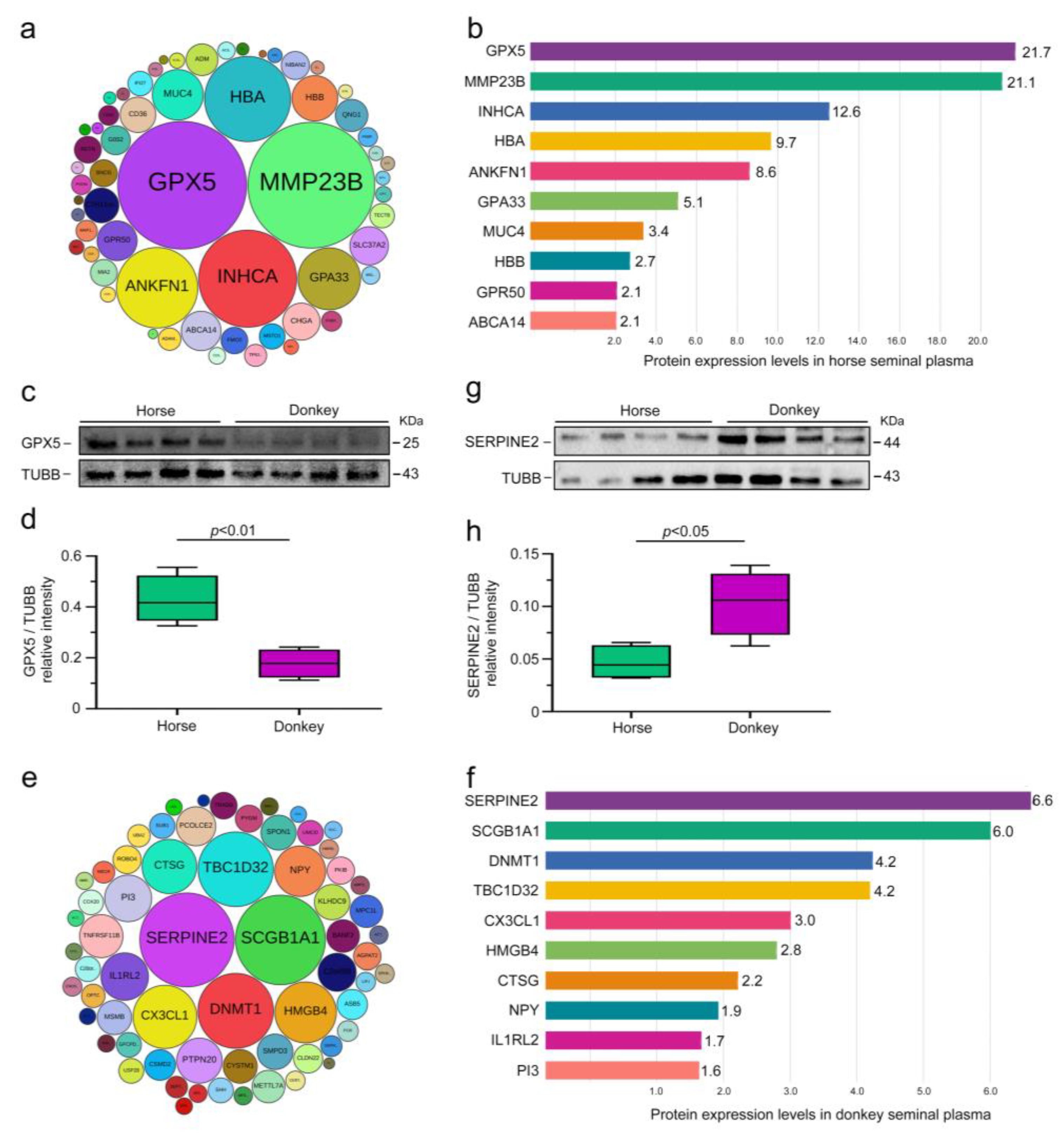
3.4. Classification of Seminal Plasma SEPs in Horses and Donkeys
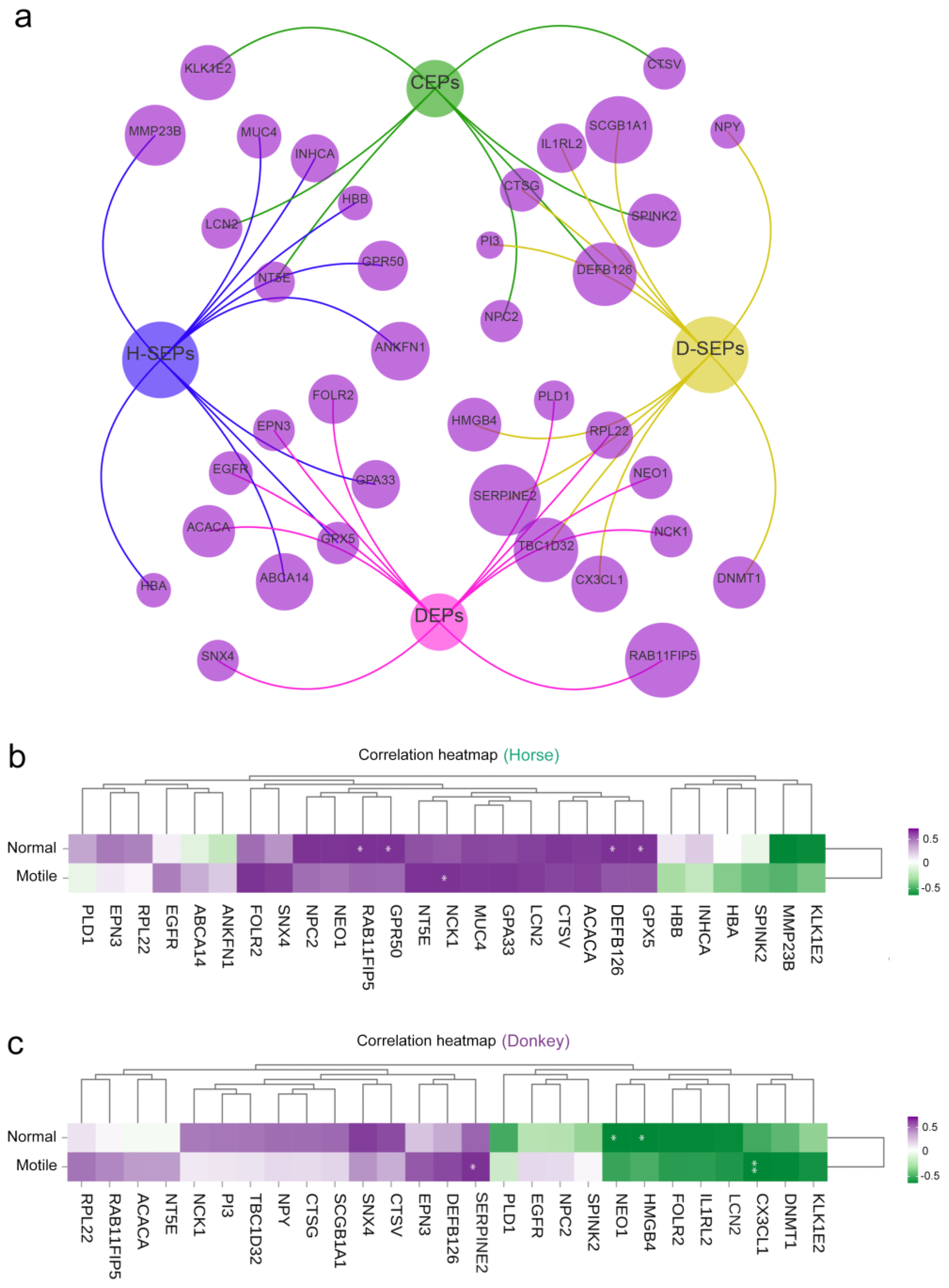
3.5. Comparison of Seminal Plasma DEPs in Horses and Donkeys
3.6. Correlation Between Horse and Donkey Seminal Plasma Protein and Sperm Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, W.; Sun, Q.; Hu, S.; Yu, P.; Kan, S.; Zhang, W. Equus mitochondrial pangenome reveals independent domestication imprints in donkeys and horses. Sci. Rep. 2025, 15, 6803. [Google Scholar] [CrossRef]
- Ren, X.; Liu, Y.; Zhao, Y.; Li, B.; Bai, D.; Bou, G.; Zhang, X.; Du, M.; Wang, X.; Bou, T.; et al. Analysis of the Whole-Genome Sequences from an Equus Parent-Offspring Trio Provides Insight into the Genomic Incompatibilities in the Hybrid Mule. Genes 2022, 13, 2188. [Google Scholar] [CrossRef]
- Bukhari, S.; Parkes, R.S.V. Assessing the impact of draught load pulling on welfare in equids. Front. Vet. Sci. 2023, 10, 1214015. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, D.; Moroni, R.; Bocci, C.; Camillo, F.; Rota, A.; Panzani, D. Interspecific and Intraspecific Artificial Insemination in Domestic Equids. Animals 2023, 13, 582. [Google Scholar] [CrossRef]
- Carluccio, A.; Bucci, R.; Fusi, J.; Robbe, D.; Veronesi, M.C. Effect of age and of reproductive status on reproductive indices in horse mares carrying mule pregnancies. Heliyon 2020, 6, e05175. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Wen, X.; He, Q.; Yi, M.; Dugarjaviin, M.; Bou, G. Comparative Study on the Sperm Proteomes of Horses and Donkeys. Animals 2024, 14, 2237. [Google Scholar] [CrossRef]
- Rodriguez-Martinez, H.; Martinez, E.A.; Calvete, J.J.; Peña Vega, F.J.; Roca, J. Seminal Plasma: Relevant for Fertility? Int. J. Mol. Sci. 2021, 22, 4368. [Google Scholar] [CrossRef]
- Vickram, S.; Rohini, K.; Srinivasan, S.; Nancy Veenakumari, D.; Archana, K.; Anbarasu, K.; Jeyanthi, P.; Thanigaivel, S.; Gulothungan, G.; Rajendiran, N.; et al. Role of Zinc (Zn) in Human Reproduction: A Journey from Initial Spermatogenesis to Childbirth. Int. J. Mol. Sci. 2021, 22, 2188. [Google Scholar] [CrossRef]
- Ali, W.; Deng, K.; Bian, Y.; Liu, Z.; Zou, H. Spectacular role of epididymis and bio-active cargo of nano-scale exosome in sperm maturation: A review. Biomed. Pharmacother. 2023, 164, 114889. [Google Scholar] [CrossRef] [PubMed]
- Ebel, F.; Ulloa, O.; Strobel, P.; Ramírez-Reveco, A. Semen Quality and Freezability Analyses in the Ejaculates of Two Poitou Donkeys in the Southern Hemisphere. Front. Vet. Sci. 2021, 8, 662887. [Google Scholar] [CrossRef]
- Kumar, V.; Hassan, M.I.; Tomar, A.K.; Kashav, T.; Nautiyal, J.; Singh, S.; Singh, T.P.; Yadav, S. Proteomic analysis of heparin-binding proteins from human seminal plasma: A step towards identification of molecular markers of male fertility. J. Biosci. 2009, 34, 899–908. [Google Scholar] [CrossRef]
- Weigel Muñoz, M.; Carvajal, G.; Curci, L.; Gonzalez, S.N.; Cuasnicu, P.S. Relevance of CRISP proteins for epididymal physiology, fertilization, and fertility. Andrology 2019, 7, 610–617. [Google Scholar] [CrossRef]
- Kyaw, H.M.; Sato, H.; Tagami, T.; Yanagawa, Y.; Nagano, M.; Katagiri, S. Effects of milk osteopontin on the endometrial epidermal growth factor profile and restoration of fertility in repeat breeder dairy cows. Theriogenology 2022, 184, 26–33. [Google Scholar] [CrossRef]
- Xu, B.; Bai, X.; Zhang, J.; Li, B.; Zhang, Y.; Su, R.; Wang, R.; Wang, Z.; Lv, Q.; Zhang, J.; et al. Metabolomic analysis of seminal plasma to identify goat semen freezability markers. Front. Vet. Sci. 2023, 10, 1132373. [Google Scholar] [CrossRef]
- Llavanera, M. Evaluation of sperm quality and male fertility: The use of molecular markers in boar sperm and seminal plasma. Anim. Reprod. Sci. 2024, 269, 107545. [Google Scholar] [CrossRef]
- Na, L.; Xu, M.; Chen, J.L.; Chen, G.J.; Sun, J.; Zhang, Q.; Li, J.Q.; Guo, X.L.; Zuo, Z.F.; Liu, X.Z.; et al. 4D-DIA quantitative proteomics revealed the core mechanism of diabetic retinopathy after berberine treatment. Eur. J. Pharmacol. 2023, 958, 175947. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, P.; Wan, Q.; Ruan, X.; Wu, P.; Hao, Y.; Zhang, Z.; Sun, J.; Nie, W.; Chen, S. High-Coverage Four-Dimensional Data-Independent Acquisition Proteomics and Phosphoproteomics Enabled by Deep Learning-Driven Multidimensional Predictions. Anal. Chem. 2023, 95, 7495–7502. [Google Scholar] [CrossRef]
- Deng, W.; Zhong, Z.; Tong, Y.; Liu, J.; Wang, X.; Xu, L.; Li, Y.; Chen, X.; Wei, Q.; Rao, J. 4D DIA-PRM proteomic study identifying modulated pathways and biomarkers associated with pelvic organ prolapse. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2023, 1230, 123916. [Google Scholar] [CrossRef]
- Dong, T.; Liang, Y.; Chen, H.; Li, Y.; Li, Z.; Gao, X. Quantitative proteomics revealed protein biomarkers to distinguish malignant pleural effusion from benign pleural effusion. J. Proteom. 2024, 302, 105201. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, G.; Xian, M.; Zhang, G.; Wen, F.; Hu, Z.; Hu, J. Proteomic Analysis of Frozen-Thawed Spermatozoa with Different Levels of Freezability in Dairy Goats. Int. J. Mol. Sci. 2023, 24, 15550. [Google Scholar] [CrossRef]
- Suliman, Y.; Becker, F.; Tuchscherer, A.; Wimmers, K. Seasonal variations in quantitative and qualitative sperm characteristics in fertile and subfertile stallions. Arch. Anim. Breed. 2020, 63, 145–154. [Google Scholar] [CrossRef]
- Gloria, A.; Contri, A.; De Amicis, I.; Robbe, D.; Carluccio, A. Differences between epididymal and ejaculated sperm characteristics in donkey. Anim. Reprod. Sci. 2011, 128, 117–122. [Google Scholar] [CrossRef]
- Płaczkowska, S.; Rodak, K.; Kmieciak, A.; Gilowska, I.; Kratz, E.M. Exploring correlations: Human seminal plasma and blood serum biochemistry in relation to semen quality. PLoS ONE 2024, 19, e0305861. [Google Scholar] [CrossRef]
- Tosi, U.; Bernabò, N.; Verni, F.; Valbonetti, L.; Muttini, A.; Mattioli, M.; Barboni, B. Postpartum reproductive activities and gestation length in Martina Franca jennies, an endangered Italian donkey breed. Theriogenology 2013, 80, 120–124. [Google Scholar] [CrossRef]
- Morrell, J.M.; Richter, J.; Martinsson, G.; Stuhtmann, G.; Hoogewijs, M.; Roels, K.; Dalin, A.M. Pregnancy rates after artificial insemination with cooled stallion spermatozoa either with or without single layer centrifugation. Theriogenology 2014, 82, 1102–1105. [Google Scholar] [CrossRef]
- Griffin, R.A.; Swegen, A.; Baker, M.A.; Ogle, R.A.; Smith, N.; Aitken, R.J.; Skerrett-Byrne, D.A.; Fair, S.; Gibb, Z. Proteomic analysis of spermatozoa reveals caseins play a pivotal role in preventing short-term periods of subfertility in stallions. Biol. Reprod. 2022, 106, 741–755. [Google Scholar] [CrossRef]
- Yu, J.; Li, M.; Ji, C.; Li, X.; Li, H.; Liu, G.; Wang, Y.; Liu, G.; Wang, T.; Che, X.; et al. Comparative proteomic analysis of seminal plasma proteins in relation to freezability of Dezhou donkey semen. Anim. Reprod. Sci. 2021, 231, 106794. [Google Scholar] [CrossRef]
- Novak, S.; Smith, T.A.; Paradis, F.; Burwash, L.; Dyck, M.K.; Foxcroft, G.R.; Dixon, W.T. Biomarkers of in vivo fertility in sperm and seminal plasma of fertile stallions. Theriogenology 2010, 74, 956–967. [Google Scholar] [CrossRef]
- Thélie, A.; Rehault-Godbert, S.; Poirier, J.C.; Govoroun, M.; Fouchécourt, S.; Blesbois, E. The seminal acrosin-inhibitor ClTI1/SPINK2 is a fertility-associated marker in the chicken. Mol. Reprod. Dev. 2019, 86, 762–775. [Google Scholar] [CrossRef]
- Busso, D.; Oñate-Alvarado, M.J.; Balboa, E.; Castro, J.; Lizama, C.; Morales, G.; Vargas, S.; Härtel, S.; Moreno, R.D.; Zanlungo, S. Spermatozoa from mice deficient in Niemann-Pick disease type C2 (NPC2) protein have defective cholesterol content and reduced in vitro fertilising ability. Reprod. Fertil. Dev. 2014, 26, 609–621. [Google Scholar] [CrossRef]
- Watanabe, H.; Takeo, T.; Tojo, H.; Sakoh, K.; Berger, T.; Nakagata, N.; Mak, T.W.; Kondoh, G. Lipocalin 2 binds to membrane phosphatidylethanolamine to induce lipid raft movement in a PKA-dependent manner and modulates sperm maturation. Development 2014, 141, 2157–2164. [Google Scholar] [CrossRef] [PubMed]
- Cochran, S.D.; Cole, J.B.; Null, D.J.; Hansen, P.J. Single nucleotide polymorphisms in candidate genes associated with fertilizing ability of sperm and subsequent embryonic development in cattle. Biol. Reprod. 2013, 89, 69. [Google Scholar] [CrossRef] [PubMed]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W.; et al. A common mutation in the defensin DEFB126 causes impaired sperm function and subfertility. Sci. Transl. Med. 2011, 3, 92ra65. [Google Scholar] [CrossRef]
- Tollner, T.L.; Yudin, A.I.; Treece, C.A.; Overstreet, J.W.; Cherr, G.N. Macaque sperm coating protein DEFB126 facilitates sperm penetration of cervical mucus. Hum. Reprod. 2008, 23, 2523–2534. [Google Scholar] [CrossRef]
- Aram, R.; Chan, P.T.K.; Cyr, D.G. Beta-defensin126 is correlated with sperm motility in fertile and infertile men. Biol. Reprod. 2020, 102, 92–101. [Google Scholar] [CrossRef]
- Syrkasheva, A.G.; Dolgushina, N.V.; Romanov, A.Y.; Burmenskaya, O.V.; Makarova, N.P.; Ibragimova, E.O.; Kalinina, E.A.; Sukhikh, G.T. Cell and genetic predictors of human blastocyst hatching success in assisted reproduction. Zygote 2017, 25, 631–636. [Google Scholar] [CrossRef]
- Novak, S.; Ruiz-Sánchez, A.; Dixon, W.T.; Foxcroft, G.R.; Dyck, M.K. Seminal plasma proteins as potential markers of relative fertility in boars. J. Androl. 2010, 31, 188–200. [Google Scholar] [CrossRef]
- Vilagran, I.; Castillo-Martín, M.; Prieto-Martínez, N.; Bonet, S.; Yeste, M. Triosephosphate isomerase (TPI) and epididymal secretory glutathione peroxidase (GPX5) are markers for boar sperm quality. Anim. Reprod. Sci. 2016, 165, 22–30. [Google Scholar] [CrossRef]
- Lu, C.H.; Lee, R.K.; Hwu, Y.M.; Chu, S.L.; Chen, Y.J.; Chang, W.C.; Lin, S.P.; Li, S.H. SERPINE2, a serine protease inhibitor extensively expressed in adult male mouse reproductive tissues, may serve as a murine sperm decapacitation factor. Biol. Reprod. 2011, 84, 514–525. [Google Scholar] [CrossRef]
- Li, S.H.; Hwu, Y.M.; Lu, C.H.; Lin, M.H.; Yeh, L.Y.; Lee, R.K. Serine Protease Inhibitor SERPINE2 Reversibly Modulates Murine Sperm Capacitation. Int. J. Mol. Sci. 2018, 19, 1520. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, C.; Han, Y.; Feng, R.; Cui, X.; Zhou, Y.; Li, Z.; Bai, Q. RNA sequencing profiling of mRNAs, long noncoding RNAs, and circular RNAs in Trigeminal Ganglion following Temporomandibular Joint inflammation. Front. Cell Dev. Biol. 2022, 10, 945793. [Google Scholar] [CrossRef] [PubMed]
- Battistini, C.; Tamagnone, L. Transmembrane semaphorins, forward and reverse signaling: Have a look both ways. Cell. Mol. Life Sci. 2016, 73, 1609–1622. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Liu, L.; Dong, M.; Xue, W.; Zhou, S.; Li, X.; Guo, S.; Yan, W. Prenatal exposure to bisphenol AF induced male offspring reproductive dysfunction by triggering testicular innate and adaptive immune responses. Ecotoxicol. Environ. Saf. 2023, 259, 115030. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Gaskins, A.J.; Hood, R.B.; Ford, J.B.; Hauser, R.; Smith, A.K.; Everson, T.M. Former smoking associated with epigenetic modifications in human granulosa cells among women undergoing assisted reproduction. Sci. Rep. 2024, 14, 5009. [Google Scholar] [CrossRef]
- Holm, J.; Hansen, S.I. Characterization of soluble folate receptors (folate binding proteins) in humans. Biological roles and clinical potentials in infection and malignancy. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140466. [Google Scholar] [CrossRef]
- Nawaz, F.Z.; Kipreos, E.T. Emerging roles for folate receptor FOLR1 in signaling and cancer. Trends Endocrinol. Metab. 2022, 33, 159–174. [Google Scholar] [CrossRef]
- Dong, J.; Li, M.; Peng, R.; Zhang, Y.; Qiao, Z.; Sun, N. ACACA reduces lipid accumulation through dual regulation of lipid metabolism and mitochondrial function via AMPK−PPARα−CPT1A axis. J. Transl. Med. 2024, 22, 196. [Google Scholar] [CrossRef]
- Abah, K.O.; Fontbonne, A.; Partyka, A.; Nizanski, W. Effect of male age on semen quality in domestic animals: Potential for advanced functional and translational research? Vet. Res. Commun. 2023, 47, 1125–1137. [Google Scholar] [CrossRef]
- Suarez, S.S. Control of hyperactivation in sperm. Hum. Reprod. Update 2008, 14, 647–657. [Google Scholar] [CrossRef]
- Antonouli, S.; Di Nisio, V.; Messini, C.; Samara, M.; Salumets, A.; Daponte, A.; Anifandis, G. Sperm plasma membrane ion transporters and male fertility potential: A perspective under the prism of cryopreservation. Cryobiology 2024, 114, 104845. [Google Scholar] [CrossRef]
- Tiwari, M.; Gujar, G.; Shashank, C.G.; Ponsuksili, S. Selection signatures for high altitude adaptation in livestock: A review. Gene 2024, 927, 148757. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, X.; Ren, H.; He, Q.; Yi, M.; Ulaangerel, T.; Bou, G. Comparative Analysis of Proteomic Characteristics in Seminal Plasma Between Horses and Donkeys. Animals 2025, 15, 1532. https://doi.org/10.3390/ani15111532
Wen X, Ren H, He Q, Yi M, Ulaangerel T, Bou G. Comparative Analysis of Proteomic Characteristics in Seminal Plasma Between Horses and Donkeys. Animals. 2025; 15(11):1532. https://doi.org/10.3390/ani15111532
Chicago/Turabian StyleWen, Xin, Hong Ren, Qianqian He, Minna Yi, Tseweendolmaa Ulaangerel, and Gerelchimeg Bou. 2025. "Comparative Analysis of Proteomic Characteristics in Seminal Plasma Between Horses and Donkeys" Animals 15, no. 11: 1532. https://doi.org/10.3390/ani15111532
APA StyleWen, X., Ren, H., He, Q., Yi, M., Ulaangerel, T., & Bou, G. (2025). Comparative Analysis of Proteomic Characteristics in Seminal Plasma Between Horses and Donkeys. Animals, 15(11), 1532. https://doi.org/10.3390/ani15111532