
Effects of Chenopodium album L. Substitution Levels and Harvest Time on In Vitro Rumen Fermentation and Methane Production in Early-Fattening Hanwoo Steers

 ,
,  , , , , , , and
, , , , , , and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
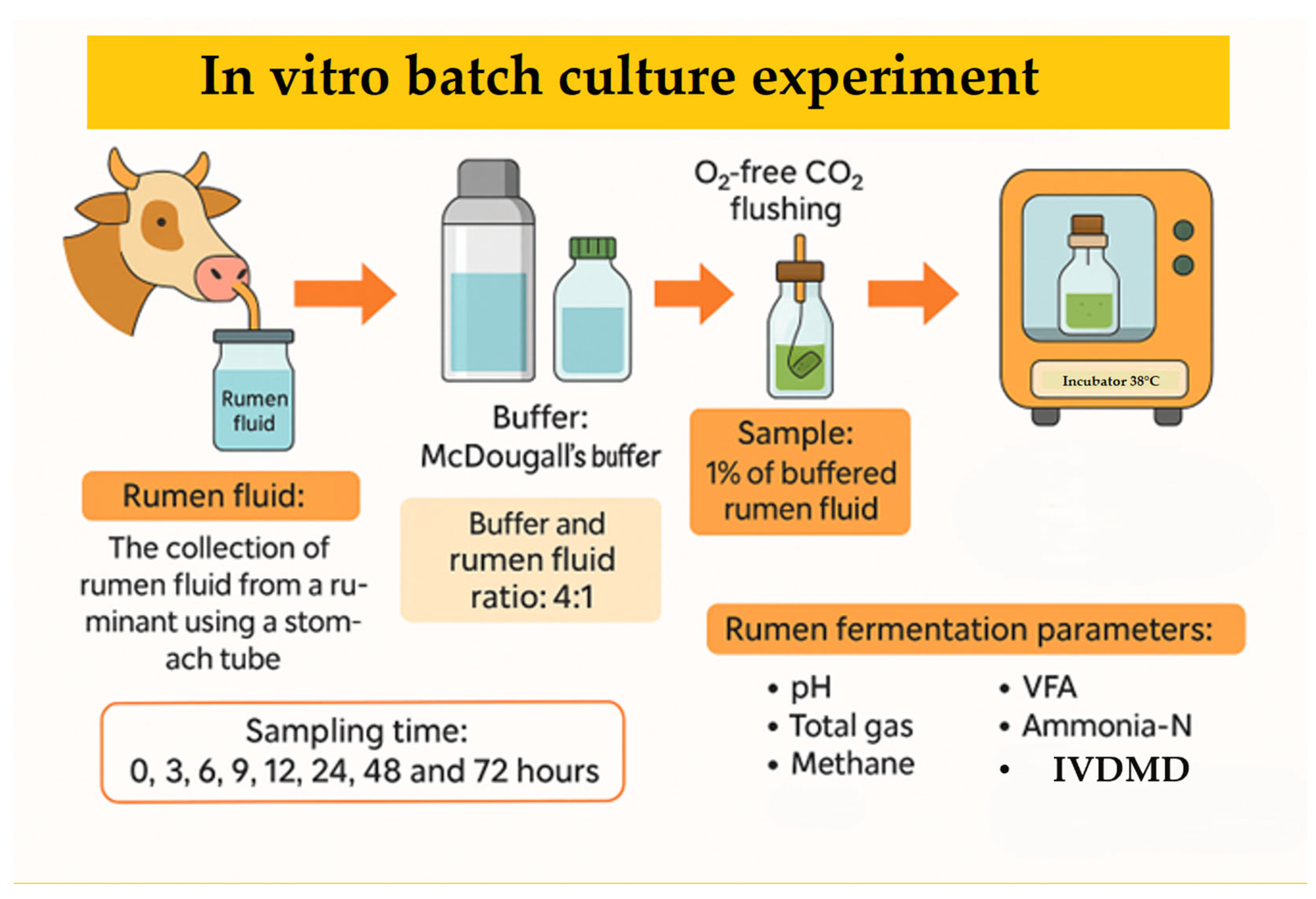
2.1. Experimental Design, Feed Preparation, and In Vitro Fermentation Procedure
2.2. Chemical Analysis and In Vitro Fermentation Measurements
2.3. Statistical Analysis
3. Results
3.1. Chemical Composition and Saponins Content of Chenopodium album L. at Different Harvest Times
3.2. Evaluation of the Effect of Harvest Time of Chenopodium album L. on In Vitro Rumen Fermentation
3.3. Effect of Chenopodium album L. Substitution Levels on In Vitro Rumen Fermentation in Early-Fattening Hanwoo
4. Discussion
4.1. Chemical Compositions and Saponins Content of Chenopodium album L. at Different Harvest Times
4.2. Evaluation of the Effect of Harvest Time of Chenopodium album L. on In Vitro Rumen Fermentation
4.3. Effect of Chenopodium album L. Substitution Levels on In Vitro Rumen Fermentation in Early-Fattening Hanwoo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CAL | Chenopodium Album L. |
| CP | Crude protein |
| NDF | Neutral detergent fiber |
| ADF | Acid detergent fiber |
| IVDMD | In vitro dry matter digestibility |
| A/P | Acetate to propionate |
| TVFA | Total volatile fatty acids |
| GHG | Greenhouse gas |
| CO2 | Carbon dioxide |
| CH4 | Methane |
| GE | Gross energy |
| OM | Organic matter |
| DM | Dry matter |
| O2 | Oxygen |
| TGP | Total gas production |
| NH3-N | Ammonia–nitrogen |
| EE | Ether extract |
| CF | Crude fiber |
| CA | Crude ash |
| USA | United States of America |
| SAS | Statistical analysis system |
| GLM | General linear model |
| SEM | Standard error of the mean |
| CNU | Chungnam National University |
| IPET | Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry |
| MAFRA | Ministry of Agriculture, Food and Rural Affairs, Korea |
| KOICA | Korea International Cooperation Agency |
| HKNU | Hankyong National University |
References
- Tullo, E.; Finzi, A.; Guarino, M. Review: Environmental Impact of Livestock Farming and Precision Livestock Farming as a Mitigation Strategy. Sci. Total Environ. 2019, 650, 2751–2760. [Google Scholar] [CrossRef]
- Joos, F.; Prentice, I.C.; Sitch, S.; Meyer, R.; Hooss, G.; Plattner, G.-K.; Gerber, S.; Hasselmann, K. Global Warming Feedbacks on Terrestrial Carbon Uptake under the Intergovernmental Panel on Climate Change (IPCC) Emission Scenarios. Glob. Biogeochem. Cycles 2001, 15, 891–907. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane Production by Ruminants: Its Contribution to Global Warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Qin, Z.; Johnson, D.W.; Johnson, R.T. Cooperative Versus Competitive Efforts and Problem Solving. Rev. Educ. Res. 1995, 65, 129–143. [Google Scholar] [CrossRef]
- Hur, S.N.; Lim, K.B.; Kim, D.A. Sustainable Roughage Production in Korea-Review. Asian-Australas. J. Anim. Sci. 1999, 12, 445–448. [Google Scholar] [CrossRef]
- Jeon, S.; Jeong, S.; Lee, M.; Seo, J.; Kam, D.K.; Kim, J.H.; Park, J.; Seo, S. Effects of Reducing Inclusion Rate of Roughages by Changing Roughage Sources and Concentrate Types on Intake, Growth, Rumen Fermentation Characteristics, and Blood Parameters of Hanwoo Growing Cattle (Bos Taurus coreanae). Asian-Australas. J. Anim. Sci. 2019, 32, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Risi, J.; Galwey, N.W. The Chenopodium Grains of the Andes: Inca Crops for Modern Agriculture. In Advances in Applied Biololgy; Coaker, T.H., Ed.; Academic Press: Cambridge, MA, USA, 1984; pp. 145–216. [Google Scholar]
- Han, K.-S.; Jung, T.-H.; Shin, K.-O. Studies on the General Analysis and Antioxidant Component Analysis of Chenopodium album Var. Centrorubrum and Biochemical Analysis of Blood of Mice Administered C. album. Korean J. Food Sci. Technol. 2019, 51, 492–498. [Google Scholar]
- Choo, Y.-S.; Song, S.-D. Ecophysiological Characteristics of Chenopodiaceous Plants-An Approach through Inorganic and Organic Solutes. Korean J. Ecol. 2000, 23, 397–406. [Google Scholar]
- Liu, Y.; Ma, T.; Chen, D.; Zhang, N.; Si, B.; Deng, K.; Tu, Y.; Diao, Q. Effects of Tea Saponin Supplementation on Nutrient Digestibility, Methanogenesis, and Ruminal Microbial Flora in Dorper Crossbred Ewe. Animals 2019, 9, 29. [Google Scholar] [CrossRef]
- Hristov, A.N.; McAllister, T.A.; Van Herk, F.H.; Cheng, K.-J.; Newbold, C.J.; Cheeke, P.R. Effect of Yucca Schidigera on Ruminal Fermentation and Nutrient Digestion in Heifers1. J. Anim. Sci. 1999, 77, 2554–2563. [Google Scholar] [CrossRef]
- Yadav, S.K.; Sehgal, S. In Vitro and in Vivo Availability of Iron from Bathua (Chenopodium album) and Spinach (Spinacia oleracia) Leaves. J. Food Sci. Technol. Mysore 2002, 39, 42–46. [Google Scholar]
- Wallace, R.J.; Arthaud, L.; Newbold, C.J. Influence of Yucca Shidigera Extract on Ruminal Ammonia Concentrations and Ruminal Microorganisms. Appl. Environ. Microbiol. 1994, 60, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Lavaud, C.; Voutquenne, L.; Bal, P.; Pouny, I. Saponins from Chenopodium album. Fitoterapia 2000, 71, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Makkar, H.P.S. Methane Mitigation from Ruminants Using Tannins and Saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef]
- Dębski, B.; Gralak, M.A.; Bertrandt, J.; Kłos, A. Comparison of Antioxidant Potential and Mineral Composition of Quinoa and Lamb’s Quarters Weed (Chenopodium album). Probl. Hig. Epidemiol. 2018, 99, 88–93. [Google Scholar]
- Damdinsuren, G. The Issue of Feed Preparation of Some Species for Undesirable Plants. Ph.D. Dissertation, Mongolian University of Life Sciences, Ulaanbaatar, Mongolia, 2016. [Google Scholar]
- Poonia, A.; Upadhayay, A. Chenopodium album Linn: Review of Nutritive Value and Biological Properties. J. Food Sci. Technol. 2015, 52, 3977–3985. [Google Scholar] [CrossRef] [PubMed]
- Prakash, D.; Pal, M. Chenopodium: Seed Protein, Fractionation and Amino Acid Composition. Int. J. Food Sci. Nutr. 1998, 49, 271–275. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Ohri, D. Genetic Variability and Heritability of Selected Traits during Different Cuttings of Vegetable Chenopodium. Indian J. Genet. Plant Breed. 2003, 63, 359–360. [Google Scholar]
- McDougall, E.I. Studies on Ruminant Saliva. 1. The Composition and Output of Sheep’s Saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 21st ed.; AOAC International: Washington, DC, USA, 2019. [Google Scholar]
- Council, N.R. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Le Bot, M.; Thibault, J.; Pottier, Q.; Boisard, S.; Guilet, D. An Accurate, Cost-Effective and Simple Colorimetric Method for the Quantification of Total Triterpenoid and Steroidal Saponins from Plant Materials. Food Chem. 2022, 383, 132597. [Google Scholar] [CrossRef]
- Beuvink, J.M.W.; Spoelstra, S.F. Interactions between Substrate, Fermentation End-Products, Buffering Systems and Gas Production upon Fermentation of Different Carbohydrates by Mixed Rumen Microorganisms in Vitro. Appl. Microbiol. Biotechnol. 1992, 37, 505–509. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, J.H.; Song, J.Y.; Bae, G.S.; Lee, M.H.; Jung, S.Y.; Kim, E.J. Effects of Energy Feed Sources on Hanwoo Rumen Fermentation Characteristics and Microbial Amino Acids Composition in Vitro. Ann. Anim. Resour. Sci. 2022, 33, 94–104. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified Reagents for Determination of Urea and Ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- SAS/SHARE V9.4 User’s Guide, SAS Institute Inc.: Cary, NC, USA, 2013.
- Fan, W.; Yuan, L.; Qu, X. CFD Simulation of Hydrodynamic Behaviors and Aerobic Sludge Granulation in a Stirred Tank with Lower Ratio of Height to Diameter. Biochem. Eng. J. 2018, 137, 78–94. [Google Scholar] [CrossRef]
- Sikora, M.C.; Hatfield, R.D.; Kalscheur, K.F. Fermentation and Chemical Composition of High-Moisture Lucerne Leaf and Stem Silages Harvested at Different Stages of Development Using a Leaf Stripper. Grass Forage Sci. 2019, 74, 254–263. [Google Scholar] [CrossRef]
- Palmonari, A.; Fustini, M.; Canestrari, G.; Grilli, E.; Formigoni, A. Influence of Maturity on Alfalfa Hay Nutritional Fractions and Indigestible Fiber Content. J. Dairy Sci. 2014, 97, 7729–7734. [Google Scholar] [CrossRef]
- Yari, M.; Valizadeh, R.; Naserian, A.A.; Jonker, A.; Azarfar, A.; Yu, P. Effects of Including Alfalfa Hay Cut in the Afternoon or Morning at Three Stages of Maturity in High Concentrate Rations on Dairy Cows Performance, Diet Digestibility and Feeding Behavior. Anim. Feed Sci. Technol. 2014, 192, 62–72. [Google Scholar] [CrossRef]
- Ebeid, H.M.; Kholif, A.E.; El-Bordeny, N.; Chrenkova, M.; Mlynekova, Z.; Hansen, H.H. Nutritive Value of Quinoa (Chenopodium quinoa) as a Feed for Ruminants: In Sacco Degradability and in Vitro Gas Production. Environ. Sci. Pollut. Res. 2022, 29, 35241–35252. [Google Scholar] [CrossRef]
- Adedapo, A.; Jimoh, F.; Afolayan, A. Comparison of the Nutritive Value and Biological Activities of the Acetone, Methanol and Water Extracts of the Leaves of Bidens pilosa and Chenopodium album. Acta Pol. Pharm. 2011, 68, 83–92. [Google Scholar]
- Singh, L.; Yadav, N.; Kumar, A.R.; Gupta, A.K.; Chacko, J.; Parvin, K.; Tripathi, U. Preparation of Value Added Products from Dehydrated Bathua Leaves (Chenopodium album Linn.). Nat. Prod. Radiance 2007, 6, 45–49. [Google Scholar]
- Odhav, B.; Beekrum, S.; Akula, U.S.; Baijnath, H. Preliminary Assessment of Nutritional Value of Traditional Leafy Vegetables in KwaZulu-Natal, South Africa. J. Food Compos. Anal. 2007, 20, 430–435. [Google Scholar] [CrossRef]
- Peiretti, P.G.; Gai, F.; Tassone, S. Fatty Acid Profile and Nutritive Value of Quinoa (Chenopodium quinoa Willd.) Seeds and Plants at Different Growth Stages. Anim. Feed Sci. Technol. 2013, 183, 56–61. [Google Scholar] [CrossRef]
- Afolayan, A.J.; Jimoh, F.O. Nutritional Quality of Some Wild Leafy Vegetables in South Africa. Int. J. Food Sci. Nutr. 2009, 60, 424–431. [Google Scholar] [CrossRef]
- Yildirim, E.; Dursun, A.; Turan, M. Determination of the Nutrition Contents of the Wild Plants Used as Vegetables in Upper Coruh Valley. Turk. J. Bot. 2001, 25, 367–371. [Google Scholar]
- Oguntona, C.R.; Razaq, M.A.; Akintola, T.T. Pattern of Dietary Intake and Consumption of Street Foods among Nigerian Students. Nutr. Health 1998, 12, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of Environmental Abiotic Factors on the Content of Saponins in Plants. Phytochem. Rev. 2011, 10, 471–491. [Google Scholar] [CrossRef]
- Anseong Climate. Available online: https://data.kma.go.kr/climate/RankState/selectRankStatisticsDivisionList.do?pgmNo=179 (accessed on 2 April 2025).
- Pecetti, L.; Tava, A.; Romani, M.; Benedetto, M.G.D.; Corsi, P. Variety and Environment Effects on the Dynamics of Saponins in Lucerne (Medicago sativa L.). Eur. J. Agron. 2006, 25, 187–192. [Google Scholar] [CrossRef]
- Sylwia, G.; Leszczynski, B.; Wieslaw, O. Effect of Low and High-Saponin Lines of Alfalfa on Pea Aphid. J. Insect Physiol. 2006, 52, 737–743. [Google Scholar] [CrossRef]
- Budan, A.; Tessier, N.; Saunier, M.; Gillmann, L.; Hamelin, J.; Chicoteau, P.; Richomme, P.; Guilet, D. Effect of Several Saponin Containing Plant Extracts on Rumen Fermentation in Vitro, Tetrahymena Pyriformis and Sheep Erythrocytes. J. Food Agric. Environ. 2013, 11, 576–582. [Google Scholar]
- Oakenfull, D. Saponins in Food—A Review. Food Chem. 1981, 7, 19–40. [Google Scholar] [CrossRef]
- Boukrouh, S.; Noutfia, A.; Moula, N.; Avril, C.; Hornick, J.-L.; Chentouf, M.; Cabaraux, J.-F. Effects of Sulla Flexuosa Hay as Alternative Feed Resource on Goat’s Milk Production and Quality. Animals 2023, 13, 709. [Google Scholar] [CrossRef] [PubMed]
- de Ruiter, J.M.; Burns, J.C. Cell Wall Carbohydrates of Flaccidgrass Plant Parts. I. Neutral Sugar Composition of Fermented Residues. Crop. Sci. 1987, 27, 1057–1063. [Google Scholar] [CrossRef]
- Balde, A.T.; Vandersall, J.H.; Erdman, R.A.; Reeves, J.B.; Glenn, B.P. Effect of Stage of Maturity of Alfalfa and Orchard grass on in Situ Dry Matter and Crude Protein Degradability and Amino Acid Composition. Anim. Feed Sci. Technol. 1993, 44, 29–43. [Google Scholar] [CrossRef]
- Yu, P.; Christensen, D.A.; McKinnon, J.J.; Markert, J.D. Effect of Variety and Maturity Stage on Chemical Composition, Carbohydrate and Protein Subfractions, in Vitro Rumen Degradability and Energy Values of Timothy and Alfalfa. Can. J. Anim. Sci. 2003, 83, 279–290. [Google Scholar] [CrossRef]
- Tang, S.X.; Tayo, G.O.; Tan, Z.L.; Sun, Z.H.; Wang, M.; Ren, G.P.; Han, X.F. Use of In Vitro Gas Production Technique to Investigate Interactions between Rice Straw, Wheat Straw, Maize Stover and Alfalfa or Clover. Asian-Australas. J. Anim. Sci. 2008, 21, 1278–1285. [Google Scholar] [CrossRef]
- Soest, P.J.V. Rice Straw, the Role of Silica and Treatments to Improve Quality. Anim. Feed Sci. Technol. 2006, 130, 137–171. [Google Scholar] [CrossRef]
- Mir, P.S.; Mir, Z.; Broersma, K.; Bittman, S.; Hall, J.W. Prediction of Nutrient Composition and in Vitro Dry Matter Digestibility from Physical Characteristics of Forages. Anim. Feed Sci. Technol. 1995, 55, 275–285. [Google Scholar] [CrossRef]
- Agustin, F.; Pazla, R.; Jamarun, N.; Suryadi, H. Exploring the Impact of Processed Cassava Peel on Microbial Dynamics and in Vitro Nutrient Digestibility in Ruminant Diets. Int. J. Vet. Sci. 2023, 13, 463–470. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994; ISBN 978-0-8014-2772-5. [Google Scholar]
- McDonald, P.; Greenhalgh, J.F.D.; Morgan, C.A.; Greenhalgh, J.F.D.; Morgan, C.A.; Edwards, R.; Sinclair, L.; Wilkinson, R. Animal Nutrition, 7th ed.; Benjamin Cummings: San Francisco, CA, USA, 2010; ISBN 978-1-4082-0427-6. [Google Scholar]
- Adams, D.C.; Cochran, R.C.; Currie, P.O. Forage Maturity Effects on Rumen Fermentation, Fluid Flow, and Intake in Grazing Steers. Rangel. Ecol. Manag. Range Manag. Arch. 1987, 40, 404–408. [Google Scholar] [CrossRef]
- Muck, R.E.; Filya, I.; Contreras-Govea, F.E. Inoculant Effects on Alfalfa Silage: In Vitro Gas and Volatile Fatty Acid Production. J. Dairy Sci. 2007, 90, 5115–5125. [Google Scholar] [CrossRef] [PubMed]
- Blümmel, M.; Makkar, H.P.S.; Becker, K. In Vitro Gas Production: A Technique Revisited. J. Anim. Physiol. Anim. Nutr. 1997, 77, 24–34. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane Emissions from Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Wilman, D.; Foulkes, G.R.; Givens, D.I. The Rate and Extent of Cell-Wall Degradation in Vitro for 40 Silages Varying in Composition and Digestibility. Anim. Feed Sci. Technol. 1996, 63, 111–122. [Google Scholar] [CrossRef]
- Topps, J.H.; Reed, W.D.C.; Elliott, R.C. The Effect of Season and of Supplementary Feeding on the Rumen Contents of African Cattle Grazing Subtropical Herbage: II. pH Values and Concentration and Proportions of Volatile Fatty Acids. J. Agric. Sci. 1965, 64, 397–402. [Google Scholar] [CrossRef]
- Hungate, R.E. The Rumen and Its Microbes; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 1-4832-6362-2. [Google Scholar]
- Eskeland, B.; Pfander, W.H.; Preston, R.L. Intravenous Energy Infusion in Lambs: Effects on Nitrogen Retentin, Plasma Free Amino Acids and Plasma Urea Nitrogen. Br. J. Nutr. 1974, 31, 201–211. [Google Scholar] [CrossRef]
- Abbasi, D.; Rouzbehan, Y.; Rezaei, J. Effect of Harvest Date and Nitrogen Fertilization Rate on the Nutritive Value of Amaranth Forage (Amaranthus hypochondriacus). Anim. Feed Sci. Technol. 2012, 171, 6–13. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. The Effect and Mode of Action of Saponins on the Microbial Populations and Fermentation in the Rumen and Ruminant Production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The Impact of Saponins or Saponin-Containing Plant Materials on Ruminant Production A Review. J. Agric. Food Chem. 2005, 53, 8093–8105. [Google Scholar] [CrossRef]
- Wang, X.; Song, J.; Liu, Z.; Zhang, G.; Zhang, Y. Fermentation Quality and Microbial Community of Corn Stover or Rice Straw Silage Mixed With Soybean Curd Residue. Animals 2022, 12, 919. [Google Scholar] [CrossRef]
- Gunun, P.; Gunun, N.; Khejornsart, P.; Ouppamong, T.; Cherdthong, A.; Wanapat, M.; Sirilaophaisan, S.; Yuangklang, C.; Polyorach, S.; Kenchaiwong, W.; et al. Effects of Antidesma thwaitesianum Muell. Arg. Pomace as a Source of Plant Secondary Compounds on Digestibility, Rumen Environment, Hematology, and Milk Production in Dairy Cows. Anim. Sci. J. 2018, 90, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and Mode of Action of Chitosan and Ivy Fruit Saponins on the Microbiome, Fermentation and Methanogenesis in the Rumen Simulation Technique. Fems Microbiol. Ecol. 2016, 92, fiv160. [Google Scholar] [CrossRef]
- Archer, K.A.; Decker, A.M. Relationship Between Fibrous Components and in Vitro Dry Matter Digestibility of Autumn-saved Grasses1. Agron. J. 1977, 69, 610–612. [Google Scholar] [CrossRef]
- Mir, M.A.; Sharma, R.K.; Rastogi, A.; Barman, K. Effect of Incorporation of Walnut Cake (Juglans regia) in Concentrate Mixture on Degradation of Dry Matter, Organic Matter and Production of Microbial Biomass in Vitro in Goat. Vet. World 2015, 8, 1172–1176. [Google Scholar] [CrossRef]
- Li, M.; Zi, X.; Zhou, H.; Lv, R.; Tang, J.; Cai, Y. Silage Fermentation and Ruminal Degradation of Cassava Foliage Prepared With Microbial Additive. Amb. Express 2019, 9, 180. [Google Scholar] [CrossRef] [PubMed]
- Poornachndra, K.T.; Malik, P.K.; Trivedi, S.; Thirumalaisamy, G.; Kolte, A.P.; Dhali, A.; Bhatta, R. Effect of Individual vs. Combined Supplementation of Tamarind Seed Husk and Soapnut on Methane Production, Feed Fermentation and Protozoal Population in Vitro. Approaches Poult. Dairy Vet. Sci. 2019, 6, 572–581. [Google Scholar] [CrossRef]
- Ampapon, T.; Phesatcha, K.; Wanapat, M. Effects of Phytonutrients on Ruminal Fermentation, Digestibility, and Microorganisms in Swamp Buffaloes. Animals 2019, 9, 671. [Google Scholar] [CrossRef]
- Shokryzadan, P.; Rajion, M.A.; Goh, Y.M.; Ishak, I.; Ramlee, M.F.; Jahromi, M.F.; Ebrahimi, M. Mangosteen Peel Can Reduce Methane Production and Rumen Biohydrogenation in Vitro. S. Afr. J. Anim. Sci. 2016, 46, 419. [Google Scholar] [CrossRef]
- Miguel, M.; Lee, S.S.; Mamuad, L.L.; Choi, Y.J.; Jeong, C.D.; Son, A.-R.; Cho, K.K.; Kim, E.T.; Kim, S.B.; Lee, S. Enhancing Butyrate Production, Ruminal Fermentation and Microbial Population Through Supplementation with Clostridium Saccharobutylicum. J. Microbiol. Biotechnol. 2019, 29, 1083–1095. [Google Scholar] [CrossRef]
- Wang, P.; Tan, Z.; Guan, L.; Tang, S.; Zhou, C.; Han, X.; Kang, J.; He, Z. Ammonia and Amino Acids Modulates Enzymes Associated With Ammonia Assimilation Pathway by Ruminal Microbiota in Vitro. Livest. Sci. 2015, 178, 130–139. [Google Scholar] [CrossRef]
- Dong, R.; Guoqiang, S.; Yu, G. Estimating in Vitro Ruminal Ammonia-N Using Multiple Linear Models and Artificial Neural Networks Based on the CNCPS Nitrogenous Fractions of Cattle Rations with Low Concentrate/Roughage Ratios. J. Anim. Physiol. Anim. Nutr. 2021, 106, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Goel, G.; Makkar, H.P.S.; Becker, K. Effects of Sesbania sesban and Carduus pycnocephalus Leaves and Fenugreek (Trigonella foenum-graecum L.) Seeds and Their Extracts on Partitioning of Nutrients from Roughage- and Concentrate-Based Feeds to Methane. Anim. Feed Sci. Technol. 2008, 147, 72–89. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Liu, J.-X.; Lu, Y.; Zhu, W.Y.; Denman, S.E.; McSweeney, C.S. Effect of Tea Saponin on Methanogenesis, Microbial Community Structure and Expression of mcrA Gene, in Cultures of Rumen Micro-organisms. Lett. Appl. Microbiol. 2008, 47, 421–426. [Google Scholar] [CrossRef]
- Jayanegara, A.; Yogianto, Y.; Wina, E.; Sudarman, A.; Kondo, M.; Obitsu, T.; Kreuzer, M. Combination Effects of Plant Extracts Rich in Tannins and Saponins as Feed Additives for Mitigating in Vitro Ruminal Methane and Ammonia Formation. Animals 2020, 10, 1531. [Google Scholar] [CrossRef]
- Yuliana, P.; Laconi, E.B.; Jayanegara, A.; Achmadi, S.S.; Samsudin, A.A. Effect of Napier Grass Supplemented With Gliricidia Sepium, Sapindus Rarak or Hibiscus Rosa-Sinensis on in Vitro Rumen Fermentation Profiles and Methanogenesis. J. Indones. Trop. Anim. Agric. 2019, 44, 167–176. [Google Scholar] [CrossRef]
- Wu, H.; Meng, Q.; Zhou, Z.; Yu, Z. Ferric Citrate, Nitrate, Saponin and Their Combinations Affect in Vitro Ruminal Fermentation, Production of Sulphide and Methane and Abundance of Select Microbial Populations. J. Appl. Microbiol. 2019, 127, 150–158. [Google Scholar] [CrossRef]
- Kholif, A.E. A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants. Vet. Sci. 2023, 10, 450. [Google Scholar] [CrossRef]
- Hundal, J.S.; Wadhwa, M.; Bakshi, M.P.S. Effect of Herbal Feed Additives Containing Saponins on Rumen Fermentation Pattern. Indian J. Anim. Sci. 2020, 90, 237–243. [Google Scholar] [CrossRef]
- Chamkhi, I.; Charfi, S.; El Hachlafi, N.; Mechchate, H.; Guaouguaou, F.-E.; El Omari, N.; Bakrim, S.; Balahbib, A.; Zengin, G.; Bouyahya, A. Genetic Diversity, Antimicrobial, Nutritional, and Phytochemical Properties of Chenopodium album: A Comprehensive Review. Food Res. Int. 2022, 154, 110979. [Google Scholar] [CrossRef]
- Al Rharad, A.; El Aayadi, S.; Avril, C.; Souradjou, A.; Sow, F.; Camara, Y.; Hornick, J.-L.; Boukrouh, S. Meta-Analysis of Dietary Tannins in Small Ruminant Diets: Effects on Growth Performance, Serum Metabolites, Antioxidant Status, Ruminal Fermentation, Meat Quality, and Fatty Acid Profile. Animals 2025, 15, 596. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment/Feed | Ingredients (%) | Chemical Composition (% of DM) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Rice Straw | CAL | Concentrate | NFC 1 | Crude Protein | Ether Extract | Crude Ash | NDF 2 | ADF 3 | |
| Control (C) | 30 | 0 | 70 | 38.47 | 13.96 | 6.20 | 6.65 | 41.80 | 21.92 |
| T1 | 25 | 5 | 70 | 38.90 | 14.40 | 6.21 | 6.50 | 40.83 | 21.25 |
| T2 | 20 | 10 | 70 | 39.33 | 14.83 | 6.23 | 6.34 | 39.87 | 20.59 |
| T3 | 15 | 15 | 70 | 39.77 | 15.26 | 6.25 | 6.19 | 38.90 | 19.92 |
| T4 | 10 | 20 | 70 | 40.20 | 15.69 | 6.27 | 6.03 | 37.93 | 19.25 |
| Rice straw | 18.58 | 3.87 | 1.82 | 7.50 | 73.34 | 47.22 | |||
| CAL (August) | 27.24 | 12.49 | 2.18 | 4.57 | 54.01 | 33.87 | |||
| Concentrate | 46.99 | 18.29 | 8.07 | 6.29 | 28.28 | 11.08 | |||
| Harvested Time | Moisture (%) | NFC 1 | Crude Protein | Ether Extract | Crude Ash | Crude Fiber | NDF 2 | ADF 3 |
|---|---|---|---|---|---|---|---|---|
| (% of DM) | ||||||||
| June | 80.83 a | 30.44 b | 18.73 a | 4.92 a | 4.57 a | 17.36 c | 43.04 c | 21.69 c |
| July | 79.52 b | 38.45 a | 13.24 b | 1.96 b | 3.71 b | 18.78 b | 43.95 b | 27.53 b |
| August | 62.50 c | 27.24 c | 12.49 b | 2.18 b | 4.40 a | 26.09 a | 54.01 a | 33.87 a |
| SEM 4 | 0.36 | 0.49 | 0.36 | 0.13 | 0.13 | 0.24 | 0.22 | 0.21 |
| p-Value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.01 | <0.0001 | <0.0001 | <0.0001 |
| Harvest Time | Saponins (%) |
|---|---|
| June | 6.60 |
| July | 6.62 |
| August | 7.05 |
| Treatment 1 | |
| Control | 0.00 |
| T1 | 0.21 |
| T2 | 0.42 |
| T3 | 0.63 |
| T4 | 0.85 |
| Incubation Time (h) | Treatments 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| June | July | August | |||
| pH | |||||
| 0 | 7.01 a | 7.01 a | 6.99 b | 0.005 | 0.037 |
| 3 | 6.77 | 6.77 | 6.79 | 0.008 | 0.185 |
| 6 | 6.83 a | 6.73 b | 6.71 b | 0.014 | 0.002 |
| 9 | 6.69 | 6.68 | 6.70 | 0.006 | 0.010 |
| 12 | 6.72 a | 6.67 b | 6.65 b | 0.011 | 0.010 |
| 24 | 6.65 b | 6.63 b | 6.67 a | 0.004 | 0.003 |
| 48 | 6.70 b | 6.65 c | 6.75 a | 0.006 | <0.0001 |
| 72 | 6.78 a | 6.71 b | 6.74 b | 0.011 | 0.008 |
| IVDMD (%) | |||||
| 3 | 38.09 a | 37.31 b | 31.92 c | 0.055 | <0.0001 |
| 6 | 41.06 a | 38.82 b | 30.87 c | 0.160 | <0.0001 |
| 9 | 46.50 a | 41.76 b | 35.76 c | 0.181 | <0.0001 |
| 12 | 55.32 a | 45.93 b | 40.49 c | 0.916 | <0.0001 |
| 24 | 72.87 a | 68.48 b | 52.35 c | 0.157 | <0.0001 |
| 48 | 77.64 a | 73.59 b | 62.00 c | 0.186 | <0.0001 |
| 72 | 82.62 a | 72.14 b | 60.99 c | 0.209 | <0.0001 |
| NH3-N (mg/100 mL) | |||||
| 0 | 2.47 a | 1.57 c | 1.83 b | 0.058 | <0.0001 |
| 3 | 3.8 | 3.7 | 3.53 | 0.090 | 0.189 |
| 6 | 3.4 | 3.47 | 3.27 | 0.079 | 0.269 |
| 9 | 2.8 | 2.6 | 2.9 | 0.094 | 0.152 |
| 12 | 2.40 b | 2.60 a | 2.23 b | 0.050 | 0.007 |
| 24 | 4.70 a | 3.56 b | 4.96 a | 0.250 | 0.015 |
| 48 | 11.00 a | 11.00 a | 8.97 b | 0.366 | 0.012 |
| 72 | 17.50 a | 15.30 b | 14.10 c | 0.111 | <0.0001 |
| Incubation Time (h) | Treatments 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| June | July | August | |||
| TGP (mL/g DM 3) | |||||
| 3 | 10.59 b | 12.22 a | 10.22 b | 0.321 | 0.010 |
| 6 | 18.96 | 19.96 | 19.46 | 0.415 | 0.303 |
| 9 | 24.13 | 22.18 | 21.34 | 0.925 | 0.171 |
| 12 | 40.06 b | 43.89 a | 42.11 ab | 0.643 | 0.016 |
| 24 | 79.91 b | 91.68 a | 75.78 b | 1.490 | 0.001 |
| 48 | 104.44 b | 122.60 a | 81.24 c | 1.668 | <0.0001 |
| 72 | 119.74 b | 136.52 a | 107.17 c | 1.518 | <0.0001 |
| CH4, (mL/g DM 3) | |||||
| 3 | 0.22 c | 0.38 a | 0.27 b | 0.009 | <0.0001 |
| 6 | 0.57 c | 0.79 b | 0.93 a | 0.029 | 0.0003 |
| 9 | 1.16 b | 1.23 b | 1.58 a | 0.045 | 0.0011 |
| 12 | 2.89 c | 3.57 a | 3.26 b | 0.050 | 0.0002 |
| 24 | 8.86 c | 10.68 a | 9.46 b | 0.053 | <0.0001 |
| 48 | 13.06 b | 18.34 a | 11.27 c | 0.250 | <0.0001 |
| 72 | 20.07 b | 21.03 a | 16.25 c | 0.165 | <0.0001 |
| CH4, (mL/g DMD 4) | |||||
| 3 | 0.61 c | 1.00 a | 0.88 b | 0.009 | <0.0001 |
| 6 | 1.35 c | 2.03 b | 2.93 a | 0.076 | <0.0001 |
| 9 | 2.33 c | 3.01 b | 3.97 a | 0.074 | <0.0001 |
| 12 | 5.46 c | 6.54 b | 8.04 a | 0.154 | <0.0001 |
| 24 | 12.33 c | 15.88 b | 18.31 a | 0.285 | <0.0001 |
| 48 | 16.64 c | 24.84 a | 19.04 b | 0.112 | <0.0001 |
| 72 | 23.49 b | 29.57 a | 26.53 ab | 1.074 | 0.020 |
| Incubation Time (h) | Treatment 1 | SEM 2 | p-Value | ||
|---|---|---|---|---|---|
| June | July | August | |||
| Total VFA (mM) | |||||
| 0 | 33.01 a | 33.16 a | 32.20 b | 0.089 | 0.0006 |
| 3 | 41.89 a | 41.58 ab | 41.19 b | 0.129 | 0.0242 |
| 6 | 39.14 a | 39.10 a | 37.65 b | 0.394 | 0.0414 |
| 9 | 38.77 c | 43.29 b | 44.28 a | 0.261 | <0.0001 |
| 12 | 52.46 b | 60.66 a | 53.10 b | 0.215 | <0.0001 |
| 24 | 75.06 a | 73.57 a | 67.55 b | 0.449 | <0.0001 |
| 48 | 99.28 a | 99.67 a | 86.93 b | 0.262 | <0.0001 |
| 72 | 102.65 b | 103.87 a | 94.82 c | 0.239 | <0.0001 |
| Acetate (% of mM) | |||||
| 0 | 61.21 a | 60.93 a | 60.32 b | 0.088 | 0.0010 |
| 3 | 62.92 a | 61.45 b | 61.80 b | 0.146 | 0.0009 |
| 6 | 61.99 | 61.00 | 62.17 | 0.391 | 0.1527 |
| 9 | 63.74 a | 62.66 b | 62.24 c | 0.061 | <0.0001 |
| 12 | 65.15 | 65.01 | 64.86 | 0.116 | 0.2669 |
| 24 | 67.35 a | 65.70 b | 66.18 b | 0.245 | 0.0080 |
| 48 | 66.58 a | 66.50 a | 65.18 b | 0.097 | <0.0001 |
| 72 | 66.75 a | 65.87 b | 65.89 b | 0.073 | 0.0002 |
| Propionate (% of mM) | |||||
| 0 | 17.69 | 17.80 | 18.14 | 0.049 | 0.0016 |
| 3 | 19.25 | 20.24 | 19.76 | 0.103 | 0.0015 |
| 6 | 18.97 | 20.18 | 18.47 | 0.151 | 0.0005 |
| 9 | 18.67 b | 19.37 a | 18.12 c | 0.129 | 0.0014 |
| 12 | 19.31 b | 19.77 a | 19.27 b | 0.081 | 0.0079 |
| 24 | 18.84 b | 20.10 a | 18.29 b | 0.264 | 0.0073 |
| 48 | 18.81 a | 18.52 b | 18.94 a | 0.078 | 0.0241 |
| 72 | 18.53 a | 18.59 a | 17.98 b | 0.099 | 0.0090 |
| Iso-butyrate (% of mM) | |||||
| 0 | 3.36 b | 3.37 b | 3.45 a | 0.012 | 0.0027 |
| 3 | 2.75 b | 2.77 ab | 2.79 a | 0.010 | 0.1106 |
| 6 | 2.96 | 2.97 | 3.05 | 0.031 | 0.1557 |
| 9 | 2.92 a | 2.71 b | 2.42 c | 0.020 | <0.0001 |
| 12 | 2.26 a | 2.00 b | 2.24 a | 0.020 | 0.0001 |
| 24 | 1.76 b | 1.81 b | 1.94 a | 0.014 | 0.0002 |
| 48 | 1.78 b | 1.71 c | 1.89 a | 0.003 | <0.0001 |
| 72 | 1.88 a | 1.81 b | 1.89 a | 0.014 | 0.0142 |
| Butyrate (% of mM) | |||||
| 0 | 11.32 b | 11.50 a | 11.55 a | 0.025 | 0.0013 |
| 3 | 9.85 b | 10.21 a | 10.29 a | 0.030 | <0.0001 |
| 6 | 10.36 | 10.07 | 10.42 | 0.172 | 0.3739 |
| 9 | 9.16 b | 9.94 a | 9.19 b | 0.104 | 0.0029 |
| 12 | 8.79 b | 9.02 a | 9.10 a | 0.057 | 0.0201 |
| 24 | 8.04 c | 8.30 b | 9.02 a | 0.043 | <0.0001 |
| 48 | 8.32 c | 8.93 b | 9.51 a | 0.101 | 0.0005 |
| 72 | 8.14 b | 9.22 a | 9.31 a | 0.045 | <0.0001 |
| Iso-valerate (% of mM) | |||||
| 0 | 3.57 b | 3.58 b | 3.65 a | 0.008 | 0.0011 |
| 3 | 2.97 | 3.00 | 3.01 | 0.014 | 0.1394 |
| 6 | 3.19 | 3.15 | 3.24 | 0.043 | 0.4181 |
| 9 | 3.01 a | 2.89 b | 2.54 c | 0.024 | <0.0001 |
| 12 | 2.40 a | 2.21 b | 2.40 a | 0.009 | <0.0001 |
| 24 | 2.13 b | 2.11 b | 2.49 a | 0.028 | 0.0001 |
| 48 | 2.59 b | 2.49 c | 2.66 a | 0.016 | 0.0008 |
| 72 | 2.88 a | 2.80 b | 2.92 a | 0.016 | 0.0023 |
| Valerate (% of mM) | |||||
| 0 | 2.86 b | 2.83 c | 2.89 a | 0.005 | 0.0003 |
| 3 | 2.3 | 2.34 | 2.35 | 0.015 | 0.1207 |
| 6 | 2.54 | 2.54 | 2.65 | 0.035 | 0.0989 |
| 9 | 2.48 a | 2.42 b | 2.19 c | 0.015 | <0.0001 |
| 12 | 2.08 b | 1.95 c | 2.13 a | 0.006 | <0.0001 |
| 24 | 1.92 b | 1.96 b | 2.10 a | 0.022 | 0.0027 |
| 48 | 1.83 b | 1.83 b | 1.97 a | 0.006 | <0.0001 |
| 72 | 1.87 b | 1.90 b | 1.98 a | 0.011 | 0.0011 |
| A/P 3 ratio | |||||
| 0 | 3.46 a | 3.42 a | 3.33 b | 0.014 | 0.0014 |
| 3 | 3.27 a | 3.04 c | 3.13 b | 0.023 | 0.0011 |
| 6 | 3.27 a | 3.02 b | 3.37 a | 0.047 | 0.0051 |
| 9 | 3.41 b | 3.24 b | 3.24 b | 0.019 | 0.0009 |
| 12 | 3.38 a | 3.29 b | 3.37 a | 0.019 | 0.0345 |
| 24 | 3.58 a | 3.27 b | 3.62 a | 0.063 | 0.015 |
| 48 | 3.54 a | 3.59 a | 3.44 b | 0.018 | 0.0034 |
| 72 | 3.60 ab | 3.54 b | 3.66 a | 0.022 | 0.0213 |
| Treatments 1 | Incubation time (h) | |||||||
| 3 | 6 | 9 | 12 | 24 | 48 | 72 | ||
| pH | ||||||||
| Control | 6.76 b | 6.75 c | 6.67 d | 6.74 a | 6.55 a | 6.54 a | 6.45 | |
| T1 | 6.78 b | 7.04 a | 6.69 c | 6.63 b | 6.53 a | 6.48 d | 6.48 | |
| T2 | 6.83 a | 6.85 b | 6.74 b | 6.58 c | 6.54 a | 6.50 c | 6.47 | |
| T3 | 6.82 a | 6.86 b | 6.83 a | 6.59 c | 6.53 a | 6.50 c | 6.46 | |
| T4 | 6.83 a | 6.81 bc | 6.72 bc | 6.57 c | 6.48 b | 6.53 b | 6.46 | |
| SEM 2 | 0.01 | 0.024 | 0.014 | 0.009 | 0.013 | 0.009 | 0.014 | |
| p value | 0.0006 | <0.0001 | 0.0002 | <0.0001 | 0.0184 | <0.0001 | 0.4068 | |
| L 3 | <0.0001 | 0.5247 | 0.0004 | <0.0001 | 0.0098 | 0.0004 | 0.9704 | |
| Q 3 | 0.0474 | 0.0004 | 0.0038 | <0.0001 | 0.0453 | 0.7337 | 0.2620 | |
| C 3 | 0.7213 | 0.0002 | 0.0007 | 0.0147 | 0.1427 | <0.0001 | 0.1792 | |
| IVDMD (%) | ||||||||
| Control | 24.27 bc | 33.42 b | 47.89 a | 51.38 b | 64.26 b | 76.11 b | 80.22 a | |
| T1 | 28.94 a | 37.13 a | 43.39 bc | 54.28 a | 65.46 ab | 76.70 b | 80.90 a | |
| T2 | 23.57 c | 34.57 ab | 44.74 b | 55.67 a | 67.16 a | 78.12 a | 80.20 a | |
| T3 | 25.85 b | 35.46 ab | 48.50 a | 54.61 a | 67.29 a | 76.54 b | 80.89 a | |
| T4 | 14.71 d | 27.90 c | 42.03 c | 44.43 c | 59.62 c | 71.06 c | 73.66 b | |
| SEM 2 | 0.613 | 0.938 | 0.605 | 0.756 | 0.740 | 0.382 | 0.480 | |
| p value | <0.0001 | 0.0004 | <0.0001 | <0.0001 | 0.0001 | <0.0001 | <0.0001 | |
| L 3 | <0.0001 | 0.0016 | 0.0062 | 0.0002 | 0.0099 | <0.0001 | <0.0001 | |
| Q 3 | <0.0001 | 0.0003 | 0.5137 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| C 3 | 0.1105 | 0.4812 | <0.0001 | 0.0099 | 0.0053 | 0.0028 | 0.0015 | |
| NH3-N (mg/100 mL) | ||||||||
| 0 | 3 | 6 | 9 | 12 | 24 | 48 | 72 | |
| Control | 1.57 c | 1.77 c | 1.18 c | 1.43 a | 1.77 a | 1.75 d | 7.76 a | 14.87 a |
| T1 | 1.51 c | 2.97 a | 1.60 b | 1.26 b | 1.57 c | 1.88 c | 6.09 b | 13.91 a |
| T2 | 1.75 b | 2.14 b | 1.55 b | 0.83 d | 1.69 ab | 1.91 c | 7.97 a | 9.50 c |
| T3 | 1.95 a | 2.88 a | 1.72 ab | 1.18 b | 1.66 b | 2.45 a | 7.55 a | 11.29 b |
| T4 | 1.52 c | 2.37 b | 1.86 a | 1.01 c | 1.43 d | 2.26 b | 7.71 a | 14.95 a |
| SEM 2 | 0.048 | 0.086 | 0.073 | 0.04 | 0.024 | 0.038 | 0.15 | 0.415 |
| p value | 0.0003 | <0.0001 | 0.0008 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| L 3 | 0.0403 | 0.0022 | <0.0001 | 0.0002 | <0.0001 | <0.0001 | 0.0166 | 0.09 |
| Q 3 | 0.0013 | 0.0002 | 0.2464 | 0.115 | 0.0387 | 0.4472 | 0.0373 | <0.0001 |
| C 3 | <0.0001 | 0.0159 | 0.0934 | 0.0843 | <0.0001 | 0.0004 | <0.0001 | 0.0023 |
| Treatments 1 | Incubation time (h) | ||||||
| 3 | 6 | 9 | 12 | 24 | 48 | 72 | |
| TGP (mL/g DM) | |||||||
| Control | 19.14 b | 37.86 b | 46.57 c | 74.57 a | 113.33 a | 138.27 ab | 148.95 a |
| T1 | 18.95 b | 40.06 a | 50.88 a | 75.34 a | 113.53 a | 140.32 a | 149.58 a |
| T2 | 19.27 b | 37.50 bc | 48.92 ab | 76.01 a | 108.38 b | 137.17 b | 147.86 ab |
| T3 | 19.52 b | 35.44 d | 47.04 bc | 71.56 b | 107.88 b | 136.30 b | 146.29 b |
| T4 | 20.22 a | 35.88 cd | 50.29 a | 68.88 b | 108.82 b | 136.10 b | 148.05 ab |
| SEM 2 | 0.204 | 0.565 | 0.674 | 0.954 | 0.646 | 0.761 | 0.646 |
| p value | 0.0109 | 0.0013 | 0.0036 | 0.0017 | 0.0001 | 0.0162 | 0.041 |
| L 3 | 0.0018 | 0.0007 | 0.1232 | 0.0005 | <0.0001 | 0.006 | 0.0316 |
| Q 3 | 0.048 | 0.1808 | 0.4379 | 0.0071 | 0.0297 | 0.4571 | 0.3417 |
| C 3 | 0.9138 | 0.0022 | 0.0003 | 0.5515 | 0.0077 | 0.0346 | 0.0193 |
| CH4 (mL/g DM 4) | |||||||
| Control | 0.60 b | 2.33 b | 3.58 d | 7.59 bc | 24.84 a | 42.55 a | 51.12 ab |
| T1 | 0.57 b | 2.47 a | 4.17 a | 8.01 a | 22.79 b | 41.21 bc | 52.58 a |
| T2 | 0.64 a | 2.42 ab | 3.71 c | 7.46 bc | 22.19 b | 41.95 ab | 51.26 ab |
| T3 | 0.58 b | 2.01 c | 3.63 cd | 7.25 c | 21.61 b | 40.43 c | 47.09 c |
| T4 | 0.63 a | 2.47 a | 4.03 b | 7.64 b | 22.44 b | 40.37 c | 50.63 b |
| SEM 2 | 0.0086 | 0.037 | 0.034 | 0.116 | 0.425 | 0.410 | 0.569 |
| p value | 0.0005 | <0.0001 | <0.0001 | 0.0113 | 0.003 | 0.0142 | 0.0005 |
| L 3 | 0.0515 | 0.1117 | 0.0075 | 0.1044 | 0.0012 | 0.0027 | 0.0048 |
| Q 3 | 0.4342 | 0.0753 | 0.817 | 0.5347 | 0.0047 | 0.8579 | 0.5487 |
| C 3 | 0.3912 | <0.0001 | <0.0001 | 0.0018 | 0.9718 | 0.6440 | 0.0002 |
| CH4 (mL/g DMD 5) | |||||||
| Control | 2.48 b | 6.99 b | 7.53 d | 14.33 b | 36.09 b | 55.84 a | 63.86 a |
| T1 | 1.97 c | 7.24 b | 8.54 b | 14.93 b | 35.11 b | 54.11 ab | 65.09 a |
| T2 | 2.44 b | 7.15 b | 8.09 c | 13.42 c | 33.05 c | 54.69 ab | 60.46 bc |
| T3 | 2.13 bc | 5.52 c | 7.42 d | 14.69 b | 32.10 c | 52.70 b | 59.23 c |
| T4 | 3.34 a | 8.84 a | 9.13 a | 16.28 a | 38.63 a | 55.63 a | 62.67 ab |
| SEM 2 | 0.141 | 0.168 | 0.09 | 0.277 | 0.495 | 0.72 | 0.945 |
| p value | 0.0004 | <0.0001 | <0.0001 | 0.0004 | <0.0001 | 0.0659 | 0.0077 |
| L 3 | 0.0018 | 0.0039 | <0.0001 | 0.0019 | 0.2176 | 0.4428 | 0.0203 |
| Q 3 | 0.0005 | <0.0001 | 0.0058 | 0.001 | <0.0001 | 0.0309 | 0.0515 |
| C 3 | 0.2586 | <0.0001 | <0.0001 | 0.0196 | 0.0003 | 0.2756 | 0.0055 |
| Treatments 1 | Incubation time (h) | |||||||
| 0 | 3 | 6 | 9 | 12 | 24 | 48 | 72 | |
| Total VFA (mM) | ||||||||
| Control | 20.52 a | 34.08 a | 40.88 c | 46.67 c | 63.47 a | 88.52 b | 81.20 c | 102.91 b |
| T1 | 20.86 a | 32.37 b | 46.44 ab | 51.39 ab | 56.75 b | 85.60 b | 78.77 c | 100.86 b |
| T2 | 20.29 ab | 32.14 b | 42.35 bc | 52.38 a | 59.33 ab | 98.28 a | 85.72 b | 104.04 ab |
| T3 | 19.46 b | 31.85 b | 42.34 bc | 47.79 bc | 62.20 a | 86.66 b | 78.86 c | 100.17 b |
| T4 | 20.67 a | 33.92 a | 50.35 a | 52.68 a | 62.32 a | 82.66 b | 91.35 a | 109.05 a |
| SEM 2 | 0.27 | 0.455 | 1.343 | 1.346 | 1.336 | 3.075 | 1.003 | 1.776 |
| p value | 0.0336 | 0.0147 | 0.0031 | 0.0293 | 0.0301 | 0.0413 | <0.0001 | 0.0364 |
| L 3 | 0.2198 | 0.5802 | 0.0057 | 0.0759 | 0.4738 | 0.2996 | <0.0001 | 0.066 |
| Q 3 | 0.1797 | 0.0013 | 0.1039 | 0.3226 | 0.0191 | 0.0443 | 0.0016 | 0.0501 |
| C 3 | 0.0063 | 0.5583 | 0.0019 | 0.011 | 0.0173 | 0.4305 | 0.0104 | 0.2108 |
| Acetate (% of mM) | ||||||||
| Control | 59.50 a | 58.89 d | 57.13 ab | 56.50 c | 58.04 c | 58.53 a | 55.61 bc | 54.41 a |
| T1 | 59.39 a | 58.93 d | 56.65 ab | 57.68 bc | 59.05 a | 58.33 a | 56.26 a | 54.18 ab |
| T2 | 59.38 a | 59.23 c | 57.72 a | 59.27 ab | 59.12 a | 58.25 ab | 55.98 ab | 53.90 b |
| T3 | 59.31 a | 59.66 b | 57.62 a | 59.41 a | 58.71 ab | 57.40 c | 55.82 abc | 52.58 c |
| T4 | 58.69 b | 60.04 a | 56.37 b | 58.35 ab | 58.46 bc | 57.52 bc | 55.45 c | 52.30 c |
| SEM 2 | 0.126 | 0.096 | 0.346 | 0.54 | 0.153 | 0.24 | 0.165 | 0.154 |
| p value | 0.0076 | <0.0001 | 0.0786 | 0.0182 | 0.0029 | 0.024 | 0.0436 | <0.0001 |
| L 3 | 0.0017 | <0.0001 | 0.6291 | 0.0098 | 0.3232 | 0.0031 | 0.1707 | <0.0001 |
| Q 3 | 0.0449 | 0.0482 | 0.0623 | 0.015 | 0.0003 | 0.8981 | 0.0108 | 0.0754 |
| C 3 | 0.1329 | 0.331 | 0.0328 | 0.3706 | 0.0443 | 0.2778 | 0.2097 | 0.0459 |
| Propionate (% of mM) | ||||||||
| Control | 15.93 b | 20.31 a | 23.60 b | 24.73 ab | 25.39 a | 25.76 c | 27.75 b | 31.65 b |
| T1 | 16.30 a | 19.74 b | 24.36 ab | 24.83 ab | 24.25 c | 25.62 c | 26.85 c | 32.02 b |
| T2 | 16.00 b | 19.54 b | 23.57 b | 24.40 bc | 24.51 bc | 26.36 b | 27.90 b | 32.34 b |
| T3 | 15.86 b | 19.40 b | 23.78 b | 24.25 c | 25.08 ab | 27.21 a | 27.73 b | 32.52 ab |
| T4 | 16.19 a | 19.62 b | 25.14 a | 25.13 a | 25.06 ab | 27.51 a | 29.54 a | 33.46 a |
| SEM 2 | 0.06 | 0.125 | 0.291 | 0.138 | 0.233 | 0.168 | 0.227 | 0.326 |
| p value | 0.002 | 0.004 | 0.0149 | 0.008 | 0.035 | <0.0001 | 0.0001 | 0.027 |
| L 3 | 0.6369 | 0.0015 | 0.0212 | 0.6341 | 0.8193 | <0.0001 | <0.0001 | 0.0025 |
| Q 3 | 0.7014 | 0.0058 | 0.071 | 0.0052 | 0.0146 | 0.1522 | 0.0006 | 0.4342 |
| C 3 | 0.0001 | 0.9847 | 0.015 | 0.0051 | 0.0224 | 0.0215 | 0.9717 | 0.4481 |
| Iso-butyrate (% of mM) | ||||||||
| Control | 5.13 b | 3.33 bc | 2.86 a | 2.57 a | 1.98 b | 1.55 c | 1.87 a | 1.62 ab |
| T1 | 5.15 b | 3.45 ab | 2.59 b | 2.32 b | 2.15 a | 1.67 a | 1.92 a | 1.65 a |
| T2 | 5.11 b | 3.46 a | 2.76 ab | 2.27 b | 2.03 b | 1.46 d | 1.80 b | 1.62 ab |
| T3 | 5.29 a | 3.47 a | 2.75 ab | 2.35 b | 2.01 b | 1.61 b | 1.92 a | 1.68 a |
| T4 | 5.17 b | 3.29 c | 2.38 c | 2.27 b | 2.04 b | 1.61 b | 1.67 c | 1.57 b |
| SEM 2 | 0.027 | 0.041 | 0.058 | 0.058 | 0.027 | 0.013 | 0.023 | 0.019 |
| p value | 0.0057 | 0.0283 | 0.0012 | 0.0225 | 0.0134 | <0.0001 | <0.0001 | 0.0278 |
| L 3 | 0.04 | 0.7124 | 0.0013 | 0.0104 | 0.8631 | 0.2798 | 0.0003 | 0.2942 |
| Q 3 | 0.5856 | 0.0026 | 0.1135 | 0.0659 | 0.1248 | 0.0238 | 0.0021 | 0.0173 |
| C 3 | 0.014 | 0.4884 | 0.0012 | 0.0827 | 0.0017 | 0.0022 | 0.0143 | 0.0688 |
| Butyrate (% of mM) | ||||||||
| Control | 9.97 bc | 11.07 a | 10.73 ab | 10.97 a | 10.22 a | 10.26 a | 9.81 a | 8.04 a |
| T1 | 10.34 ab | 11.07 a | 10.58 ab | 10.22 ab | 9.96 c | 10.21 a | 9.93 a | 7.89 b |
| T2 | 9.59 c | 11.03 ab | 10.47 b | 9.56 b | 9.78 d | 10.01 b | 9.73 a | 7.49 d |
| T3 | 10.14 b | 10.87 bc | 10.40 b | 9.28 b | 9.78 d | 9.90 c | 9.39 b | 7.58 c |
| T4 | 10.65 a | 10.84 c | 10.98 a | 9.69 b | 10.09 b | 9.69 d | 8.94 c | 7.59 c |
| SEM 2 | 0.147 | 0.058 | 0.146 | 0.305 | 0.027 | 0.03 | 0.066 | 0.02 |
| p value | 0.0051 | 0.0421 | 0.1074 | 0.0209 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| L 3 | 0.0314 | 0.0046 | 0.4949 | 0.0047 | 0.0004 | <0.0001 | <0.0001 | <0.0001 |
| Q 3 | 0.0167 | 0.3786 | 0.0198 | 0.0405 | <0.0001 | 0.0495 | 0.0005 | <0.0001 |
| C 3 | 0.0443 | 0.3382 | 0.2069 | 0.5521 | 0.0337 | 0.5571 | 0.3162 | 0.0289 |
| Isovalerate (% of mM) | ||||||||
| Control | 5.06 c | 3.55 c | 3.15 a | 2.92 a | 2.36 bc | 1.98 b | 2.63 c | 2.36 c |
| T1 | 5.13 bc | 3.72 ab | 2.95 c | 2.61 b | 2.46 a | 2.07 a | 2.71 a | 2.36 c |
| T2 | 5.05 c | 3.76 a | 3.06 b | 2.54 b | 2.35 c | 1.90 c | 2.48 d | 2.39 b |
| T3 | 5.22 a | 3.68 b | 3.08 ab | 2.57 b | 2.29 d | 2.01 b | 2.67 b | 2.42 a |
| T4 | 5.17 ab | 3.55 c | 2.84 d | 2.51 b | 2.39 d | 2.01 b | 2.32 e | 2.32 d |
| SEM 2 | 0.03 | 0.016 | 0.026 | 0.085 | 0.011 | 0.013 | 0.012 | 0.007 |
| p value | 0.0092 | <0.0001 | <0.0001 | 0.0397 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| L 3 | 0.007 | 0.6046 | 0.0001 | 0.01 | 0.0042 | 0.8061 | <0.0001 | 0.3144 |
| Q 3 | 0.9893 | <0.0001 | 0.1423 | 0.0862 | 0.227 | 0.1156 | <0.0001 | <0.0001 |
| C 3 | 0.4121 | 0.1715 | <0.0001 | 0.2497 | <0.0001 | 0.0119 | <0.0001 | <0.0001 |
| Valerate (% of mM) | ||||||||
| Control | 3.99 d | 2.71 c | 2.48 a | 1.88 b | 2.00 d | 1.79 d | 2.30 c | 2.07 c |
| T1 | 4.03 c | 2.85 ab | 2.35 b | 2.11 a | 2.23 a | 2.00 a | 2.49 a | 2.08 c |
| T2 | 4.01 cd | 2.89 a | 2.43 a | 2.07 a | 2.07 c | 1.87 c | 2.35 b | 2.10 c |
| T3 | 4.16 a | 2.84 b | 2.44 a | 2.11 a | 2.00 d | 1.92 b | 2.49 a | 2.57 a |
| T4 | 4.07 b | 2.71 c | 2.06 c | 2.05 ab | 2.11 b | 2.03 a | 2.11 d | 2.26 b |
| SEM 2 | 0.009 | 0.014 | 0.018 | 0.056 | 0.011 | 0.013 | 0.006 | 0.03 |
| p value | <0.0001 | <0.0001 | <0.0001 | 0.0754 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| L 3 | <0.0001 | 0.9185 | <0.0001 | 0.0858 | 0.7714 | <0.0001 | <0.0001 | <0.0001 |
| Q 3 | 0.0176 | <0.0001 | <0.0001 | 0.0362 | 0.0038 | 0.5233 | <0.0001 | 0.1133 |
| C 3 | <0.0001 | 0.5334 | <0.0001 | 0.3659 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| A/P 4 ratio | ||||||||
| Control | 3.76 a | 2.90 e | 2.42 ab | 2.29 b | 2.31 c | 2.27 a | 2.00 b | 1.72 a |
| T1 | 3.64 c | 2.99 d | 2.36 b | 2.31 b | 2.44 a | 2.26 a | 2.07 a | 1.69 a |
| T2 | 3.71 ab | 3.03 c | 2.45 a | 2.39 ab | 2.42 ab | 2.21 a | 2.01 ab | 1.66 a |
| T3 | 3.71 ab | 3.12 a | 2.42 ab | 2.42 ab | 2.31 c | 2.11 b | 2.01 ab | 1.65 a |
| T4 | 3.69 b | 3.06 b | 2.47 a | 2.79 a | 2.36 bc | 2.08 b | 1.87 c | 1.55 b |
| SEM 2 | 0.017 | 0.01 | 0.025 | 0.129 | 0.021 | 0.022 | 0.02 | 0.028 |
| p value | 0.0064 | <0.0001 | 0.0752 | 0.1094 | 0.0036 | 0.0002 | 0.0006 | 0.0169 |
| L 3 | 0.201 | <0.0001 | 0.0399 | 0.0196 | 0.7801 | <0.0001 | 0.0004 | 0.0013 |
| Q 3 | 0.0806 | <0.0001 | 0.4375 | 0.2159 | 0.0158 | 0.3203 | 0.001 | 0.2602 |
| C 3 | 0.003 | 0.0073 | 0.3634 | 0.5213 | 0.001 | 0.1512 | 0.7817 | 0.4031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batburged, N.; Bae, G.-S.; Damdinsuren, G.; Kim, S.-Y.; Lee, H.-A.; Jung, S.-Y.; Kang, I.-K.; Choi, D.-H.; Kim, C.-H. Effects of Chenopodium album L. Substitution Levels and Harvest Time on In Vitro Rumen Fermentation and Methane Production in Early-Fattening Hanwoo Steers. Animals 2025, 15, 1372. https://doi.org/10.3390/ani15101372
Batburged N, Bae G-S, Damdinsuren G, Kim S-Y, Lee H-A, Jung S-Y, Kang I-K, Choi D-H, Kim C-H. Effects of Chenopodium album L. Substitution Levels and Harvest Time on In Vitro Rumen Fermentation and Methane Production in Early-Fattening Hanwoo Steers. Animals. 2025; 15(10):1372. https://doi.org/10.3390/ani15101372
Chicago/Turabian StyleBatburged, Narantuya, Gui-Seck Bae, Gurbazar Damdinsuren, Sang-Yoon Kim, Hye-An Lee, Soo-Yeon Jung, In-Ki Kang, Da-Hyun Choi, and Chang-Hyun Kim. 2025. "Effects of Chenopodium album L. Substitution Levels and Harvest Time on In Vitro Rumen Fermentation and Methane Production in Early-Fattening Hanwoo Steers" Animals 15, no. 10: 1372. https://doi.org/10.3390/ani15101372
APA StyleBatburged, N., Bae, G.-S., Damdinsuren, G., Kim, S.-Y., Lee, H.-A., Jung, S.-Y., Kang, I.-K., Choi, D.-H., & Kim, C.-H. (2025). Effects of Chenopodium album L. Substitution Levels and Harvest Time on In Vitro Rumen Fermentation and Methane Production in Early-Fattening Hanwoo Steers. Animals, 15(10), 1372. https://doi.org/10.3390/ani15101372