
Streptococcus equi subsp. zooepidemicus: Epidemiological and Genomic Findings of an Emerging Pathogen in Central Italy

 , , , , , , ,
, , , , , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Target Population
2.2. Sampling Design and Collection
2.3. Data Management and Statistical Analysis
2.4. Isolation, PCR, and Sequencing of SEZ
3. Results
3.1. SEZ Prevalence
3.2. Genomic Characterisation and Cluster Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SEZ | Streptococcus equi subsp. zooepidemicus |
| IZSAM | Istituto Zooprofilattico Sperimentale dell’Abruzzo e Molise |
| WGS | Whole Genome Sequencing |
References
- Kuusi, M.; Lahti, E.; Virolainen, A.; Hatakka, M.; Vuento, R.; Rantala, L.; Vuopio-Varkila, J.; Seuna, E.; Karppelin, M.; Hakkinen, M.; et al. An outbreak of Streptococcus equi subspecies zooepidemicus associated with consumption of fresh goat cheese. BMC Infect. Dis. 2006, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Balter, S.; Benin, A.; Pinto, S.W.; Teixeira, L.M.; Alvim, G.G.; Luna, E.; Jackson, D.; LaClaire, L.; Elliott, J.; Facklam, R.; et al. Epidemic nephritis in Nova Serrana, Brazil. Lancet 2000, 355, 1776–1780. [Google Scholar] [CrossRef]
- Torres Rosângela, S.L.A.; Santos Talita, Z.; Bernardes Andre, F.L.; Soares Patricia, A.; Soares Ana, C.C.; Dias Ricardo, S. Outbreak of Glomerulonephritis Caused by Streptococcus zooepidemicus SzPHV5 Type in Monte Santo de Minas, Minas Gerais, Brazil. J. Clin. Microbiol. 2018, 56, 10-1128. [Google Scholar] [CrossRef]
- Madzar, D.; Hagge, M.; Moller, S.; Regensburger, M.; Lee, D.; Schwab, S.; Jantsch, J. Endogenous endophthalmitis complicating Streptococcus equi subspecies zooepidemicus meningitis: A case report. BMC Res. Notes 2015, 8, 184. [Google Scholar] [CrossRef]
- Franceschi, G.; Soffritti, A.; Mantovani, M.; Digaetano, M.; Prandini, F.; Sarti, M.; Bedini, A.; Meschiari, M.; Mussini, C. Streptococcus equi Subspecies zooepidemicus Endocarditis and Meningitis in a 62-Year-Old Horse Rider Patient: A Case Report and Literature Review. Microorganisms 2024, 12, 2201. [Google Scholar] [CrossRef]
- Minces, L.R.; Brown, P.J.; Veldkamp, P.J. Human meningitis from Streptococcus equi subsp. zooepidemicus acquired as zoonoses. Epidemiol. Infect. 2011, 139, 406–410. [Google Scholar] [CrossRef]
- Pelkonen, S.; Lindahl, S.B.; Suomala, P.; Karhukorpi, J.; Vuorinen, S.; Koivula, I.; Vaisanen, T.; Pentikainen, J.; Autio, T.; Tuuminen, T. Transmission of Streptococcus equi subspecies zooepidemicus infection from horses to humans. Emerg. Infect. Dis. 2013, 19, 1041–1048. [Google Scholar] [CrossRef]
- Bosica, S.; Chiaverini, A.; De Angelis, M.E.; Petrini, A.; Averaimo, D.; Martino, M.; Rulli, M.; Saletti, M.A.; Cantelmi, M.C.; Ruggeri, F.; et al. Severe Streptococcus equi Subspecies zooepidemicus Outbreak from Unpasteurized Dairy Product Consumption, Italy. Emerg. Infect. Dis. 2023, 29, 1020–1024. [Google Scholar] [CrossRef]
- Cantelmi, M.C.; Merola, C.; Averaimo, D.; Chiaverini, A.; Cito, F.; Cocco, A.; Di Teodoro, G.; De Angelis, M.E.; Di Bernardo, D.; Auzino, D.; et al. Identification of the Novel Streptococcus equi subsp. zooepidemicus Sequence Type 525 in Donkeys of Abruzzo Region, Italy. Pathogens 2023, 12, 750. [Google Scholar] [CrossRef]
- Centers for Epidemiology and Animal Health; USDA; Animal and Plant Health Inspective Service (APHIS). Infectious Upper Respiratory Disease in U.S. Horses: Laboratory Results for Influenza Serology and Nasal Swab Culture for Streptococcus Isolation. 2001. Available online: https://www.aphis.usda.gov/sites/default/files/equine98_is_iurd.pdf (accessed on 6 January 2024).
- European Parliament and of the Council. Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’). 2016. Available online: http://data.europa.eu/eli/reg/2016/429/oj (accessed on 10 January 2024).
- Ministero della Salute di concerto con il Ministero delle Politiche Agricole, Alimentari e Forestali. DECRETO 30 Settembre 2021 Gestione e Funzionamento Dell’anagrafe Degli Equini. (21A07453) (GU Serie Generale n.302 del 21-12-2021). 2021. Available online: https://www.gazzettaufficiale.it/eli/id/2021/12/21/21A07453/sg (accessed on 15 January 2024).
- Il Presidente della Repubblica. Decreto Legislativo 5 Agosto 2022, n.134, Disposizioni in Materia di Sistema di Identificazione e Registrazione Degli Operatori, Degli Stabilimenti e Degli Animali per L’adeguamento della Normativa Nazionale alle Disposizioni del Regolamento (UE) 2016/429, ai Sensi Dell’articolo 14, Comma 2, Lettere a), b), g), h), i) e p), della Legge 22 Aprile 2021, n. 53. 2022. Available online: https://www.gazzettaufficiale.it/eli/id/2022/09/12/22G00142/sg (accessed on 18 January 2024).
- Mcgowan, C. Welfare of Aged Horses. Animals 2011, 1, 366–376. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef]
- Davis, S.; Pettengill, J.B.; Luo, Y.; Payne, J.; Shpuntoff, A.; Rand, H.; Strain, E. CFSAN SNP Pipeline: An Automated Method for Constructing SNP Matrices from Next-Generation Sequence Data. PeerJ Comput. Sci. 2015, 1, e20. [Google Scholar] [CrossRef]
- Libardoni, F.; Machado, G.; Gressler, L.T.; Kowalski, A.P.; Diehl, G.N.; dos Santos, L.C.; Corbellini, L.G.; de Vargas, A.C. Prevalence of Streptococcus equi subsp. equi in horses and associated risk factors in the State of Rio Grande do Sul, Brazil. Res. Vet. Sci. 2016, 104, 53–57. [Google Scholar] [CrossRef]
- Nocera, F.P.; Capozzi, L.; Simone, D.; Pizzano, F.; Iovane, V.; Bianco, A.; Parisi, A.; De Martino, L. Multi-locus sequence typing and in vitro antimicrobial resistance of equine Streptococcus equi subspecies zooepidemicus strains. Vet. Res. Commun. 2024, 48, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Chalker, V.J.; Waller, A.; Webb, K.; Spearing, E.; Crosse, P.; Brownlie, J.; Erles, K. Genetic diversity of Streptococcus equi subsp. zooepidemicus and doxycycline resistance in kennelled dogs. J. Clin. Microbiol. 2012, 50, 2134–2136. [Google Scholar] [CrossRef]
- Mangano, E.R.; Jones, G.M.C.; Suarez-Bonnet, A.; Waller, A.S.; Priestnall, S.L. Streptococcus zooepidemicus in dogs: Exploring a canine pathogen through multilocus sequence typing. Vet. Microbiol. 2024, 292, 110059. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, Z.; Wang, L.; Zhang, B.; Su, L. Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China. Microorganisms 2024, 12, 110059. [Google Scholar] [CrossRef]
- Retamar, G.C.; Bustos, C.P.; Guillemi, E.C.; Becu, T.; Ivanissevich, A.; Mesplet, M.; Munoz, A.J. Streptococcus equi subsp. zooepidemicus: High molecular diversity of Argentinian strains isolated from mares with endometritis. Res. Vet. Sci. 2024, 173, 105242. [Google Scholar] [CrossRef]
- Webb, K.; Jolley, K.A.; Mitchell, Z.; Robinson, C.; Newton, J.R.; Maiden, M.C.J.; Waller, A. Development of an unambiguous and discriminatory multilocus sequence typing scheme for the Streptococcus zooepidemicus group. Microbiology 2008, 154, 3016–3024. [Google Scholar] [CrossRef]
- Rasmussen, C.D.; Haugaard, M.M.; Petersen, M.R.; Nielsen, J.M.; Pedersen, H.G.; Bojesen, A.M. Streptococcus equi subsp. zooepidemicus isolates from equine infectious endometritis belong to a distinct genetic group. Vet. Res. 2013, 44, 26. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, Y.; Zhang, X.; Tian, X.; Liang, Y.; Pan, F.; Song, H.; Xia, L.; Wu, Z.; Wang, W.; et al. The clinical characteristics of Streptococcus equi ssp. zooepidemicus causing acute death in pigs and its prevention with chimeric monoclonal antibodies. Vet. Microbiol. 2025, 302, 110420. [Google Scholar] [CrossRef]
- Chen, X.; Mou, K.; Lu, W.; Schumacher, L.; Resende-De-Macedo, N.; Sitthicharoenchai, P.; Derscheid, R.; Burrough, E.; Li, G. Genomic characterization of Streptococcus equi subspecies zooepidemicus from a 2021 outbreak in Indiana with increased sow mortality. mSphere 2023, 8, e00404-23. [Google Scholar] [CrossRef] [PubMed]
- Courtney, H.S.; Dale, J.B.; Hasty, D.L. Host cell specific adhesins of group A streptococci. Adv. Exp. Med. Biol. 1997, 418, 605–606. [Google Scholar] [CrossRef] [PubMed]
- Courtney, H.S.; Dale, J.B.; Hasty, D.I. Differential effects of the streptococcal fibronectin-binding protein, FBP54, on adhesion of group A streptococci to human buccal cells and HEp-2 tissue culture cells. Infect. Immun. 1996, 64, 2415–2419. [Google Scholar] [CrossRef]
- Wen, Y.; Tsou, C.; Kuo, H.; Wang, J.; Wu, J.; Liao, P. Differential secretomics of Streptococcus pyogenes reveals a novel peroxide regulator (PerR)-regulated extracellular virulence factor mitogen factor 3 (MF3). Mol. Cell. Proteom. 2011, 10, M110.007013. [Google Scholar] [CrossRef]
- Alber, J.; El-Sayed, A.; Estoepangestie, S.; Lammler, C.; Zschock, M. Dissemination of the superantigen encoding genes seeL, seeM, szeL and szeM in Streptococcus equi subsp. equi and Streptococcus equi subsp. zooepidemicus. Vet. Microbiol. 2005, 109, 135–141. [Google Scholar] [CrossRef]
- Morris, E.R.A.; Wu, J.; Bordin, A.I.; Lawhon, S.D.; Cohen, N.D. Differences in the Accessory Genomes and Methylomes of Strains of Streptococcus equi subsp. equi and of Streptococcus equi subsp. zooepidemicus Obtained from the Respiratory Tract of Horses from Texas. Microbiol. Spectr. 2022, 10, e00764-21. [Google Scholar] [CrossRef]
- Blank, L.M.; Hugenholtz, P.; Nielsen, L.K. Evolution of the hyaluronic acid synthesis (has) operon in Streptococcus zooepidemicus and other pathogenic streptococci. J. Mol. Evol. 2008, 67, 13–22. [Google Scholar] [CrossRef]
- Crater, D.L.; van de Rijn, I. Hyaluronic Acid Synthesis Operon (has) Expression in Group A Streptococci. J. Biol. Chem. 1995, 270, 18452–18458. [Google Scholar] [CrossRef]

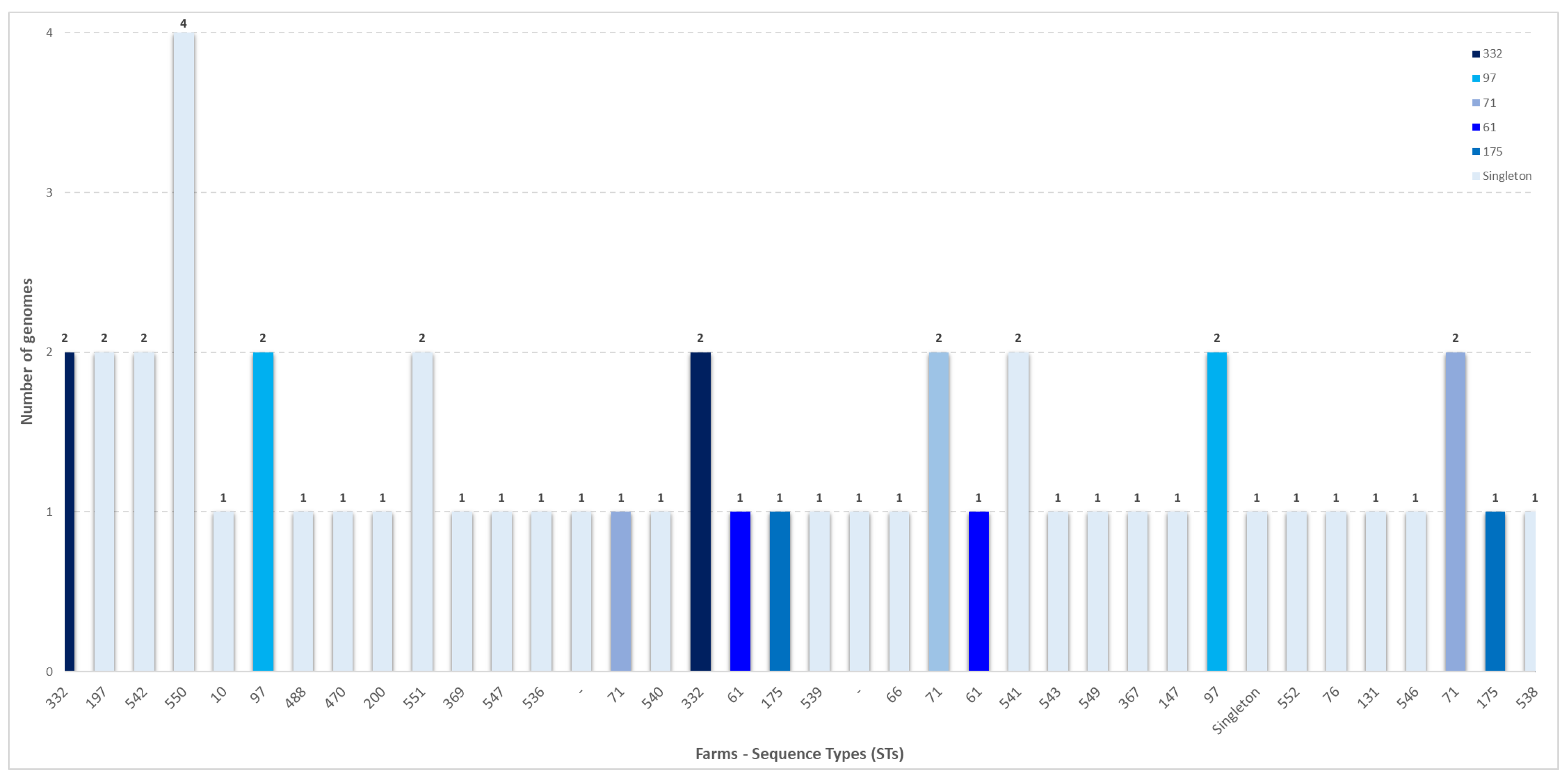
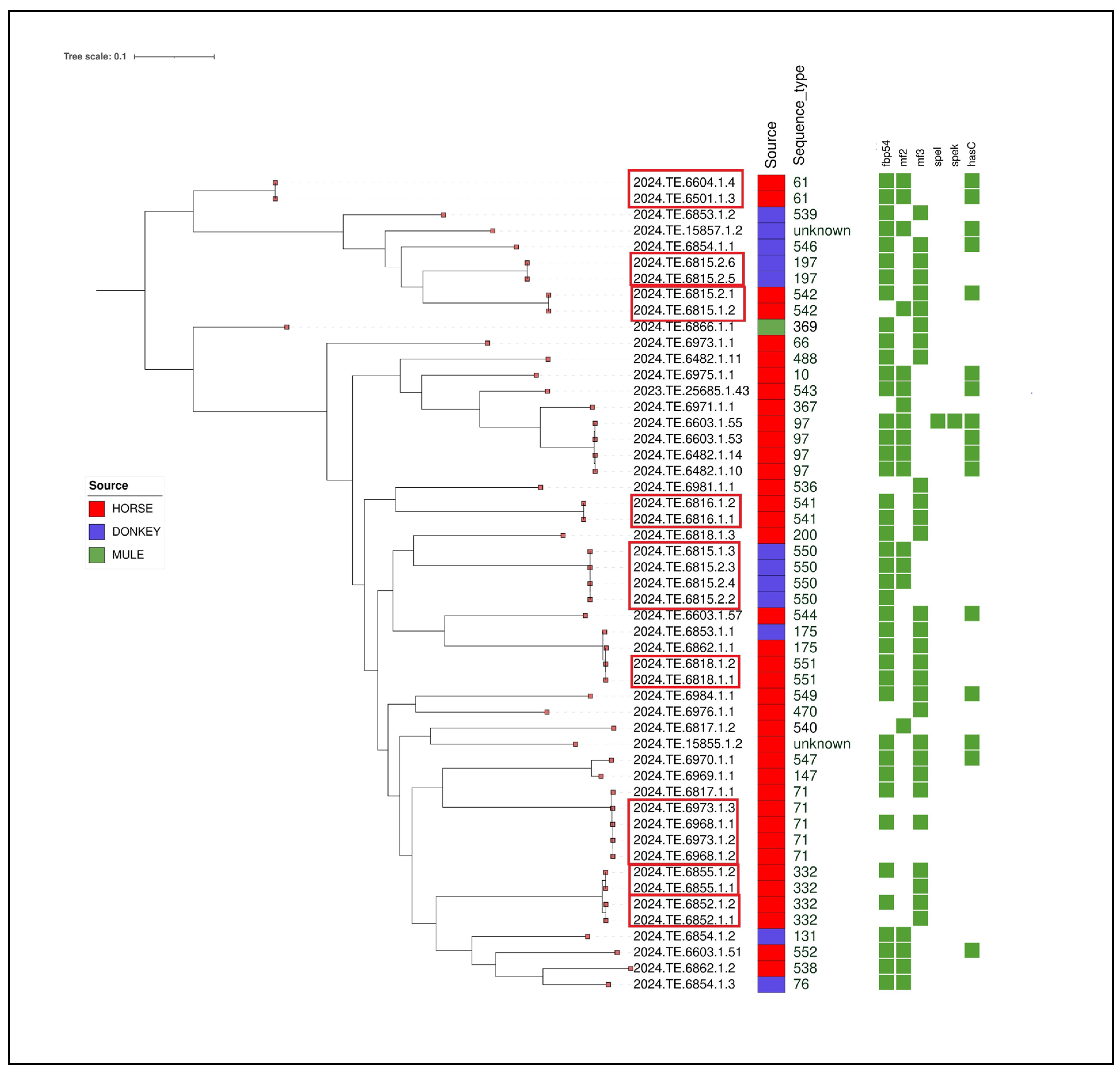

{kind=link}
{kind=link}
{kind=link}
| Province | Total Herds | Total Animals | Positive Herds (Number of Positive Herds/ Number of Herds Tested) | Positive Animals (Number of Positive Animals/ Number of Animals Tested) * |
|---|---|---|---|---|
| AQ | 1866 (41%) | 12,142 (58%) | 64% (9/14) | 36% (52/145) |
| CH | 624 (14%) | 1950 (9%) | 26% (10/38) | 17% (13/78) |
| PE | 552 (12%) | 2043 (10%) | 40% (6/15) | 28% (14/50) |
| TE | 802 (18%) | 2796 (13%) | 89% (8/9) | 29% (45/155) |
| CB | 725 (16%) | 2065 (10%) | 55% (11/20) | 40% (16/40) |
| IS | 481 (11%) | 2592 (12%) | 33% (1/3) | 50% (3/6) |
| Total | 5050 (100%) | 23,588 (100%) | 46% (45/99) | 30% (143/474) |
| Coefficients | Estimate | Std. Error | z Value | Pr (>|z|) |
|---|---|---|---|---|
| (Intercept) | −0.43583 | 0.35862 | −1.215 | 0.224 |
| Sex M | −0.11497 | 0.22638 | −0.508 | 0.612 |
| Species Equine | −0.43152 | 0.26911 | −1.603 | 0.109 |
| Breed YES | 0.17963 | 0.32104 | 0.560 | 0.576 |
| Symptoms YES | −0.41085 | 0.53616 | −0.766 | 0.444 |
| Dairy species YES | 0.32565 | 0.22357 | 1.457 | 0.145 |
| Intended for food production YES | −0.08249 | 0.24038 | −0.343 | 0.731 |
| Age_category YOUNG | −0.47630 | 0.28268 | −1.685 | 0.092 |
| ID | Source | Province | ST | Fbp54 | mf2 | mf3 | spel | spek | hasC | Biosample |
|---|---|---|---|---|---|---|---|---|---|---|
| 2024.TE.6815.2.1 | Horse | Teramo | 542 | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192347 |
| 2024.TE.6815.1.2 | Horse | Teramo | 542 | Absence | Presence | Presence | Absence | Absence | Absence | SAMN48192348 |
| 2024.TE.6815.2.2 | Donkey | Teramo | 550 | Presence | Absence | Absence | Absence | Absence | Absence | SAMN48192349 |
| 2024.TE.6815.1.3 | Donkey | Teramo | 550 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192350 |
| 2024.TE.6815.2.3 | Donkey | Teramo | 550 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192351 |
| 2024.TE.6815.2.4 | Donkey | Teramo | 550 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192352 |
| 2024.TE.6815.2.5 | Donkey | Teramo | 197 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192353 |
| 2024.TE.6815.2.6 | Donkey | Teramo | 197 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192354 |
| 2024.TE.6816.1.1 | Horse | Teramo | 541 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192355 |
| 2024.TE.6816.1.2 | Horse | Teramo | 541 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192356 |
| 2023.TE.25685.1.43 | Horse | Teramo | 543 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192357 |
| 2024.TE.6817.1.1 | Horse | L’Aquila | 71 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192358 |
| 2024.TE.6817.1.2 | Horse | L’Aquila | 540 | Absence | Presence | Absence | Absence | Absence | Absence | SAMN48192359 |
| 2024.TE.6818.1.1 | Horse | Teramo | 551 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192360 |
| 2024.TE.6818.1.2 | Horse | Teramo | 551 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192361 |
| 2024.TE.6818.1.3 | Horse | Teramo | 200 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192362 |
| 2024.TE.6852.1.1 | Horse | Campobasso | 332 | Absence | Absence | Presence | Absence | Absence | Absence | SAMN48192363 |
| 2024.TE.6852.1.2 | Horse | Campobasso | 332 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192364 |
| 2024.TE.6853.1.1 | Donkey | Pescara | 175 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192365 |
| 2024.TE.6853.1.2 | Donkey | Pescara | 539 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192366 |
| 2024.TE.6854.1.1 | Donkey | Teramo | 546 | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192367 |
| 2024.TE.6854.1.2 | Donkey | Teramo | 131 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192368 |
| 2024.TE.6854.1.3 | Donkey | Teramo | 76 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192369 |
| 2024.TE.6855.1.1 | Horse | Isernia | 332 | Absence | Absence | Presence | Absence | Absence | Absence | SAMN48192370 |
| 2024.TE.6855.1.2 | Horse | Isernia | 332 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192371 |
| 2024.TE.6970.1.1 | Horse | Pescara | 547 | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192372 |
| 2024.TE.6862.1.1 | Horse | Teramo | 175 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192373 |
| 2024.TE.6862.1.2 | Horse | Teramo | 538 | Presence | Presence | Absence | Absence | Absence | Absence | SAMN48192374 |
| 2024.TE.6866.1.1 | Mule | Pescara | 369 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192375 |
| 2024.TE.6968.1.1 | Horse | Chieti | 71 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192376 |
| 2024.TE.6968.1.2 | Horse | Chieti | 71 | Absence | Absence | Absence | Absence | Absence | Absence | SAMN48192377 |
| 2024.TE.6969.1.1 | Horse | Chieti | 147 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192378 |
| 2024.TE.6971.1.1 | Horse | Teramo | 367 | Absence | Presence | Absence | Absence | Absence | Absence | SAMN48192379 |
| 2024.TE.6973.1.1 | Horse | L’Aquila | 66 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192380 |
| 2024.TE.6973.1.2 | Horse | L’Aquila | 71 | Absence | Absence | Absence | Absence | Absence | Absence | SAMN48192381 |
| 2024.TE.6973.1.3 | Horse | L’Aquila | 71 | Absence | Absence | Absence | Absence | Absence | Absence | SAMN48192382 |
| 2024.TE.6975.1.1 | Horse | Chieti | 10 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192383 |
| 2024.TE.6976.1.1 | Horse | Campobasso | 470 | Absence | Absence | Presence | Absence | Absence | Absence | SAMN48192384 |
| 2024.TE.6981.1.1 | Horse | Campobasso | 536 | Absence | Absence | Presence | Absence | Absence | Absence | SAMN48192385 |
| 2024.TE.6984.1.1 | Horse | Teramo | 549 | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192386 |
| 2024.TE.15857.1.2 | Donkey | L’Aquila | unknown | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192387 |
| 2024.TE.6603.1.51 | Horse | L’Aquila | 552 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192388 |
| 2024.TE.6482.1.11 | Horse | Campobasso | 488 | Presence | Absence | Presence | Absence | Absence | Absence | SAMN48192389 |
| 2024.TE.6482.1.10 | Horse | Campobasso | 97 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192390 |
| 2024.TE.6482.1.14 | Horse | Campobasso | 97 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192391 |
| 2024.TE.6603.1.57 | Horse | L’Aquila | 544 | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192392 |
| 2024.TE.6603.1.53 | Horse | L’Aquila | 97 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192393 |
| 2024.TE.15855.1.2 | Horse | L’Aquila | unknown | Presence | Absence | Presence | Absence | Absence | Presence | SAMN48192394 |
| 2024.TE.6603.1.55 | Horse | L’Aquila | 553 | Presence | Presence | Absence | Presence | Presence | Presence | SAMN48192395 |
| 2024.TE.6501.1.3 | Horse | Campobasso | 61 | Presence | Presence | Absence | Absence | Absence | Presence | SAMN48192396 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cito, F.; Di Francesco, C.E.; Averaimo, D.; Chiaverini, A.; Alessiani, A.; Di Domenico, M.; Cresci, M.; Rulli, M.; Cantelmi, M.C.; Di Bernardo, M.D.; et al. Streptococcus equi subsp. zooepidemicus: Epidemiological and Genomic Findings of an Emerging Pathogen in Central Italy. Animals 2025, 15, 1351. https://doi.org/10.3390/ani15101351
Cito F, Di Francesco CE, Averaimo D, Chiaverini A, Alessiani A, Di Domenico M, Cresci M, Rulli M, Cantelmi MC, Di Bernardo MD, et al. Streptococcus equi subsp. zooepidemicus: Epidemiological and Genomic Findings of an Emerging Pathogen in Central Italy. Animals. 2025; 15(10):1351. https://doi.org/10.3390/ani15101351
Chicago/Turabian StyleCito, Francesca, Cristina Esmeralda Di Francesco, Daniela Averaimo, Alexandra Chiaverini, Alessandra Alessiani, Marco Di Domenico, Marta Cresci, Marco Rulli, Maria Chiara Cantelmi, Maria Daniela Di Bernardo, and et al. 2025. "Streptococcus equi subsp. zooepidemicus: Epidemiological and Genomic Findings of an Emerging Pathogen in Central Italy" Animals 15, no. 10: 1351. https://doi.org/10.3390/ani15101351
APA StyleCito, F., Di Francesco, C. E., Averaimo, D., Chiaverini, A., Alessiani, A., Di Domenico, M., Cresci, M., Rulli, M., Cantelmi, M. C., Di Bernardo, M. D., Giammarino, A., Vincifori, G., & Petrini, A. (2025). Streptococcus equi subsp. zooepidemicus: Epidemiological and Genomic Findings of an Emerging Pathogen in Central Italy. Animals, 15(10), 1351. https://doi.org/10.3390/ani15101351