
Improving Human Diets and Welfare through Using Herbivore-Based Foods: 2. Environmental Consequences and Mitigations


Abstract
Simple Summary
Abstract
1. Introduction
2. Land Use
- Traditional extensive grazing in southern Europe and mountainous ecosystems created a mosaic of habitats resulting in biodiversity;
- Short-term abandonment following overgrazing had positive effects in central Europe and lowlands but not in southern European mountains;
- The abandonment of long-term traditional grazing activities in mountainous ecosystems resulted in the extinction of populations of species tightly linked with open habitats; and
- In lowlands, the abandonment of some grazing patches augments habitat diversification and creates new habitats for more species, but overgrazing caused a significant decrease in biodiversity.

{kind=link}
{kind=link}
{kind=link}
| Production System | Comment | Reference | |||
|---|---|---|---|---|---|
| Meat from Feedlot-Fed Animals | Meat from Grazed-Pasture Animals | Plant-Based Meat | Lab-Grown Meat | ||
| Beef +*22 | Western Canada—lifetime GHG emissions | [50] | |||
| Dairy +3.8 to +6.2 | Mycoprotein based +2.4 to +2.6 | [51,52] | |||
| +1.8 to +2.3 | [45] | ||||
| Beef +9 to +42; Dairy +1 to +2 | Suckler herds +23 to +52 Extensive pastoral +12 to +129 | +1 to +2 | Review of numerous studies | [53] | |
| +31 | +7.5 | Feedlot beef in upper midwest USA | [44] | ||
| Dairy +4.3 to +4.9 Chicken meat +5.2 to +5.8 | Soymeal-based +2.6 to +2.8 Mycoprotein-based +5.5 to +6.1 | +23.9 to +24.6 | Cradle-to-plate life cycle | [43] | |
| +150 | +35 | [54] | |||
| +33—US feedlot | +3.5 per kg beef | Total cradle-to-distribution impacts Beyond Burger and U.S. beef in feedlot | [55] | ||
| Feedlot from +6.09 to +6.12 due to soil erosion | Rotationally grazed systems moved from +9.62 to −6.65 due to soil erosion | Change due to inclusion of soil organic matter accumulation in analysis | [56] | ||
| +48.4 | Full LCA for USA beef—includes feed production and feedlot | [57] | |||
| +33—US feedlot beef | −3.5 for grazed pastures | Rotational grazed beef can in some circumstances have a negative carbon impact due to soil C sequestration | [58] | ||
| +21.3 | Full LCA for USA beef for feedlot and processing, etc. | [59] | |||
| Beef +48 to +210 Dairy +35 to +45 Sheep +80 to +190 | +5 to +35 | +15 to +40 | [60] | ||
| +11 (feedlot finished beef) | −3.5 (grazed) | +3.5 (soy-based) +3 (pea-based) | Cradle-to-distribution LCA, but excludes GHGE potential of retail, restaurant, or at-home use, and end-of-life stages | [61] | |
| +42 to +235 | +21 to +55 | Range depends on functional unit and allocation method | [62] | ||
| +6.01 for sheep and +8.97 for beef cattle | “Cradle-to-grave” for average NZ sheep and beef (weighted for traditional and dairy beef)—grazed pasture | [63] | |||
| +4.9 to +25.2 | [64] | ||||
| Traditional beef +10.09 Dairy beef +6.88 Sheep +6.01 | NZ—Cradle-to-farm-gate GHG emissions per kg live-weight sold (kg CO2e kg−1 LW) for grazed pasture | [65] | |||
| +11 (dairy +40 (beef) | +2.2 to +24.8 | Current benchmark; best and worst case for cultured meat | [66] | ||
3. Greenhouse Gas Emissions
- Feed type and quality: Improving the nutritive value of the grazed feed through replacing low-quality native pasture with improved higher-quality pasture increases the enteric methane emission (g/day) produced by ruminants but reduces the methane yield per unit of meat or wool produced [86,87]. The increase in dietary lipids that improves nutritive value through balancing the ratios of energy to protein in diets has been shown to reduce greenhouse gas emissions [88]. This has led to programs seeking to increase lipid levels in ryegrass [89,90]. It is also well known that the incorporation of species into pasture that express condensed tannins, which protect protein in the rumen will reduce greenhouse gas emissions [91,92]. Other forage species have been shown to reduce methane emissions when eaten such as biserrula (Biserrula pelecinus) [93,94], sulla (Hedysarum coronarium) [91,95], Lotus corniculatus [91,96], L. pedunculatus [97], and sainfoin (Onobrychis viciifolia) [98]. However, these species are agronomically inferior to the forages currently used and their management under grazing is a challenge. A program in white clover (Trifolium repens), the most used pasture legume in temperate areas, is set to achieve condensed tannin expression in leaf tissue through the use of a transcription factor taken from a closely related Trifolium species [99,100].
- Dietary additives such as oils, microalgae, macroalgae, nitrate, ionophores, protozoal control, phytochemicals from plant extracts, and 3-nitrooxypropanol have shown differing levels of efficacy in reducing methane production per kg dry matter consumed [101,102,103,104]. Macroalgae and 3-nitrooxypropanol have shown the greatest efficacy in reducing methane yield. The seaweeds Asparagopsis taxiformis and A. armata, when included at low concentrations in the feed of cattle and sheep, inhibit methanogenesis by up to 98% [105,106]. The active ingredient from these macroalgae are bromoforms (organic compounds that are classified as a probable human carcinogen by the US EPA, but can be found in chlorinated drinking water [107]). Bromoform inhibits an enzyme in the methanogenesis pathway [108]. Studies are mixed on whether there are negative impacts on animal health or food quality [109]. However, because bromoforms are rapidly metabolized by rumen microbes [110], to be effective they need to be included with feed at a rate of 0.4–1.0 mg/kg animal/day of bromoform [111]. For animals in pasture, the major difficulty is longevity of action. Further, to be cost effective the expense associated with wild harvest and indeed aquaculture production will need to be reduced [109]. Canola oil has been shown to reduce methane losses from cattle, but animal performance may be compromised due to lower feed intake and decreased fiber digestibility [112] Fumaric acid, which can utilize hydrogen (instead of it combining with carbon to form methane), has been disappointing as an additive [112,113,114], but when encapsulated in partially hydrogenated vegetable oil it suppressed methane formation by 19% [115]. The main ways that many of these additives reduce methane production is through reducing the number of rumen protozoa and inhibiting methanogen activity, increasing propionic acid production, which competes with methanogens for hydrogen, and inhibiting the activity of enzymes involved in methanogen activity [116].
- The breeding of animals with higher growth rates and increased fecundity [86,87,117]. Breeding ruminants with lower methane production has been shown to be a feasible option [118,119,120] with heritability of g methane/day of 0.29 ± 0.05, and for g methane/kg DMI of 0.13 ± 0.03 [121]. Breeding for animals with low methane production per unit of dry matter intake is unlikely to negatively affect fecal egg counts, adult ewe fertility, and litter survival traits, with no evidence for significant genetic correlations, but may reduce wool, live weight, and fat deposition traits [122].
- Rumen microbial manipulations through the use of vaccines [87,123,124,125]. A recent review has concluded that it is complicated to evaluate the real effectiveness of this strategy with few published studies that have directly assessed the complete approach from vaccination to enteric animal methane emission measurement [126]. Similarly, the antibiotic monensin as a rumen additive has shown some success in vitro but results from in vivo trials have been disappointing [113].
- Pasture management, which ensures grazing occurs when fiber content is low (e.g., prior to grasses maturing and flowering) has been proposed as a method of reducing methane emissions [91].
4. Water Use and Quality
- N fertilizer used in conjunction with urease inhibitors such as N-(n-butyl)-thiophosphoric triamide and N-(n-propyl)-thiophosphoric triamide with an ability to reduce N2O and ammonia emissions while preserving yield [168]. However, caution has been called for from a meta-analysis that concluded that urease inhibitors applied with 20–30 kg N/ha per application in the spring and autumn are unlikely to increase plant dry matter yields and lead to improved NUE [169].
- Supplementary feed formulations including essential oils [170].
- Plant breeding [174] to exploit genetic variation among and within species in traits that have the potential to improve NUE (such as condensed tannin content as discussed above [100]), internal and external critical N concentrations [175], protein degradability [176,177], and biological nitrification inhibition [178].
- Combining traits in complementary forage species mixtures [179] rather than monoculture grass or simple two-species mixtures could substantially reduce N leakage to the environment. For example, in New Zealand, a combination of a N-fixing legume (e.g., white clover) with a N-demanding grass (e.g., perennial ryegrass, which has a relatively high critical internal N content) and a herb that inhibits nitrification in the soil and/or dilutes the N concentration of urine (e.g., plantain, [178,180,181]), has been shown to reduce N leaching by up to 80% in lysimeter studies [182] and 40% in field studies [183]. Proof of practice for this approach is currently underway in whole-farm systems experiments over multiple years [184].
5. Carbon Sequestration
- Fertilizer application: N inputs (10 to 20 kg N/ha/year) to low-fertility grasslands can increase soil carbon [197]. However, whether or not N inputs are associated with increased soil carbon depends on grazing intensity [198,199]. A process-based model of the dynamics of carbon and N cycling between plants, soils, and animals in grazed temperate pastures indicated that the optimal N input for balancing food production, carbon sequestration, N loss to the environment, and greenhouse gas emissions in New Zealand is approximately 150 kg/ha N fertilizer [154]. Alternatively, P fertilizer application appears to have little effect on soil carbon accumulation following conversion from native vegetation to grasslands for grazing [196,200].
- Irrigation can result in variable and contradictory impacts on soil carbon. In desert and semi-arid areas, irrigation can increase soil carbon substantially, while in humid environments, no consistent effects have been observed [201]. Whitehead et al. [192] concluded that “no change or decreases in soil carbon stocks in response to irrigation in humid climates but increases could be expected at more arid sites where plant productivity is very low prior to irrigation”. In New Zealand, irrigation has been shown to decrease soil C [196,202] due possibly to effects on soil N levels in different soil types and management systems [203], whereas in arid and semi-arid environments, irrigation might be expected to increase soil C stocks due to increased plant growth and inputs to soils [201]. However, under irrigation it is likely that N2O emissions will increase by up to 140% [201].
- Refraining from draining peaty soils, which contain high quantities of soil C (and which can lose soil organic matter through the oxidation of organic matter after drainage) [204].
- Use of supplementary feed, such as hay or silage, which may result in small increases in soil carbon on paddocks where it is used but may also result in a small decrease in soil carbon at locations where it is produced due to the “length of time between harvest and re-establishment of the new crop, maximizing returns of organic residues, and adopting minimum tillage and direct-drill methods to reduce disturbance” [192].
- Increased forage production also tends to increase soil carbon [207]. The Conant et al. review [207] indicated that the main drivers were use of more permanent pasture, improved grazing management, use of legumes, and increasing earthworm numbers. Grazing intensity can also impact soil carbon loss or accumulation. Overgrazing is generally considered to result in reduced soil carbon [197,198,208,209]. However, some studies have shown decreased soil carbon at both high and low grazing frequencies but most often the maximum accumulation occurred at a moderate grazing intensity [210,211]. This depends on balance—Parsons et al. [198]. A comparison of C4 and C3 gases in a meta-analysis has shown that higher grazing intensity results in increases in soil carbon in C4 grasslands but decreases in C3 grasslands [212]. This difference could be due to the high lignin levels in C4 grasses, which slow their decomposition and subsequent carbon release [213]. However, it is generally accepted that soil organic matter is greater in grazed pastures than non-grazed grasslands or land used for row crops or hay production [214].
- Species and diversity of species used: Whitehead et al. [192] concluded that forage species with deeper rooting and higher fine-root density at greater depths could increase soil carbon stocks. However, evidence that increased species diversity would increase soil carbon is inconclusive. The Jena Experiment setup in 2002 in Germany to investigate the effects of plant diversity on element cycling and trophic interactions do support increased soil carbon with increased pasture diversity, but the research involved mowing 2–4 times per year, i.e., there was no food production aspect [215].
- Use of biochar as a soil amendment may lead to an increase in soil carbon levels but its use as a widespread amendment to pasture soils is in many cases impractical [192].
- Full inversion tillage, which seeks to bury topsoil with high carbon levels to depths below 40 cm while bringing to the surface soil with a high carbon saturation deficit [192]. This would be achievable only on flat to moderately contoured sites and would be useful only where the soil carbon value for the topsoil is at least twofold greater than that of the subsoil [216].
6. Concluding Comments and Looking to the Future
- Increased use of white clover and plantain in pasture seed mixtures;
- Ensuring that pasture is composed of forage species that are highly digestible with high protein and high energy, and low fiber content;
- Using ruminant animals with higher growth rates and increased fecundity;
- Reduced tillage when resowing and use of a nitrification inhibitor when using N fertilizer on intensive pastures;
- Including forages with levels of condensed tannins and possibly other phytochemicals that reduce methane emissions and do not affect palatability;
- Matching the use of fertilizer N with the demand driven by the number of animals per hectare to ensure the efficient conversion of N and feed inputs to milk and meat;
- Direct injection of animal slurry into soil where this is feasible;
- Maximizing carbon sequestration in soil where and when this is possible through reducing soil disturbance and fallowing; and
- Avoiding overgrazing, which can negatively affect persistence and result in soil disturbance through resowing.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caradus, J.R.; Chapman, D.F.; Rowarth, J.S. Improving human diets and welfare through using herbivore-based foods: 1. Human and animal perspectives. Animals, 2024; in press. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2023. In Urbanization, Agrifood Systems Transformation and Healthy Diets Across the Rural–Urban Continuum; FAO: Rome, Italy, 2023. [Google Scholar] [CrossRef]
- Mottet, A.; de Haan, C.; Falcuccia, A.; Tempioa, G.; Opioa, C.; Gerbera, P. Livestock: On our plates or eating at our table? A new analysis of the feed/food debate. Glob. Food Secur. 2017, 14, 1–8. [Google Scholar] [CrossRef]
- Mathur, M.B.; Peacock, J.; Reichling, D.B.; Nadler, J.; Bain, P.A.; Gardner, C.D.; Robinson, T.N. Interventions to reduce meat consumption by appealing to animal welfare: Meta-analysis and evidence-based recommendations. Appetite 2021, 164, 105277. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.; Aiking, H. Considering how farm animal welfare concerns may contribute to more sustainable diets. Appetite 2022, 168, 105786. [Google Scholar] [CrossRef]
- Woods, M. Rural-urban linkages. In Routledge Handbook of Sustainable and Regenerative Food Systems; Duncan, J., Wiskerke, J.S.C., Carolan, M., Eds.; Taylor & Francis Group: Abingdon, UK, 2020; Chapter 26; pp. 363–375. [Google Scholar]
- Braimoh, A.K.; Vlek, P.L. Impact of land use on soil resources. In Land Use and Soil Resources; Braimoh, A.K., Vlek, P.L.G., Eds.; Springer: Dordrecht, Netherlands, 2008; pp. 1–7. [Google Scholar] [CrossRef]
- Houghton, R.A. The worldwide extent of land-use change. BioScience 1994, 44, 305–313. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef]
- Borrelli, P.; Robinson, D.A.; Fleischer, L.R.; Lugato, E.; Ballabio, C.; Alewell, C.; Meusburger, K.; Modugno, S.; Schütt, B.; Ferro, V.; et al. An assessment of the global impact of 21st century land use change on soil erosion. Nat. Commun. 2017, 8, 2013. [Google Scholar] [CrossRef]
- Richardson, B.F. Finance, food, and future urban zones: The failure of flexible development in Auckland, New Zealand. Land Use Policy 2022, 119, 106203. [Google Scholar] [CrossRef]
- Feldt, T.; Karg, H.; Kadaouré, I.; Bessert, L.; Schlecht, E. Growing struggle over rising demand: How land use change and complex farmer-grazier conflicts impact grazing management in the Western Highlands of Cameroon. Land Use Policy 2020, 95, 104579. [Google Scholar] [CrossRef]
- Bergius, M.; Benjaminsen, T.A.; Maganga, F.; Buhaug, H. Green economy, degradation narratives, and land-use conflicts in Tanzania. World Dev. 2020, 129, 104850. [Google Scholar] [CrossRef]
- Jenberu, A.A.; Admasu, T.G. Urbanization and land use pattern in Arba Minch town, Ethiopia: Driving forces and challenges. GeoJournal 2020, 85, 761–778. [Google Scholar] [CrossRef]
- Zaman, H.M.; Saqib, Z.; Bokhari, A.S.; Akhtar, N.; Amir, S. The dynamics of urbanizations and concomitant land use land cover transformations in planned and quasi-planned urban settlements of Pakistan. Geogr. Environ. Sustain. 2020, 13, 107–120. [Google Scholar] [CrossRef]
- Sencébé, Y.; Pinton, F.; Cazell, A.A. On the unequal coexistence of agrifood systems in Brazil. Rev. Agric. Food Environ. Stud. 2020, 101, 191–212. [Google Scholar] [CrossRef]
- Seifollahi-Aghmiuni, S.; Kalantari, Z.; Egidi, G.; Gaburova, L.; Salvati, L. Urbanisation-driven land degradation and socioeconomic challenges in peri-urban areas: Insights from Southern Europe. Ambio 2022, 51, 1446–1458. [Google Scholar] [CrossRef] [PubMed]
- Kanowski, P.; Edwards, P. Forests under the Southern Cross: The forest environmental frontier in Australia and New Zealand. Ambio 2021, 50, 2183–2198. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.S.; Richardson, S.J.; Carswell, F.E.; Mason, N.W.; Burrows, L.E. Knowing when native regeneration is for you, and what you should do about it. The Aotearoa New Zealand context. N. Z. J. Ecol. 2023, 47, 3524. [Google Scholar] [CrossRef]
- de Jong, L.; De Bruin, S.; Knoop, J.; van Vliet, J. Understanding land-use change conflict: A systematic review of case studies. J. Land Use Sci. 2021, 16, 223–239. [Google Scholar] [CrossRef]
- Laurance, W.F. Switch to corn promotes Amazon deforestation. Science 2007, 318, 1721. [Google Scholar] [CrossRef] [PubMed]
- Phalan, B.; Bertzky, M.; Butchart, S.H.M.; Donald, P.F.; Scharlemann, J.P.W.; Stattersfield, A.J.; Balmford, A. Crop expansion and conservation priorities in tropical countries. PLoS ONE 2013, 8, e51759. [Google Scholar] [CrossRef]
- Bodo, T.; Gimah, B.G.; Seomoni, K.J. Deforestation and habitat loss: Human causes, consequences and possible solutions. J. Geogr. Res. 2021, 4, 22–30. [Google Scholar] [CrossRef]
- Symes, W.S.; Edwards, D.P.; Miettinen, J.; Rheindt, F.E.; Carrasco, L.R. Combined impacts of deforestation and wildlife trade on tropical biodiversity are severely underestimated. Nat. Commun. 2018, 9, 4052. [Google Scholar] [CrossRef] [PubMed]
- Paiva, P.F.; de Lourdes Pinheiro Ruivo, M.; da Silva Júnior, O.M.; de Nazaré Martins Maciel, M.; Braga, T.G.; de Andrade, M.M.; dos Santos Junior, P.C.; da Rocha, E.S.; de Freitas, T.P.; da Silva Leite, T.V.; et al. Deforestation in protect areas in the Amazon: A threat to biodiversity. Biodivers. Conserv. 2020, 29, 19–38. [Google Scholar] [CrossRef]
- Levers, C.; Romero-Muñoz, A.; Baumann, M.; De Marzo, T.; Fernández, P.D.; Gasparri, N.I.; Gavier-Pizarro, G.I.; Waroux, Y.L.; Piquer-Rodríguez, M.; Semper-Pascual, A.; et al. Agricultural expansion and the ecological marginalization of forest-dependent people. Proc. Natl. Acad. Sci. USA 2021, 118, e2100436118. [Google Scholar] [CrossRef]
- Sartorello, Y.; Pastorino, A.; Bogliani, G.; Ghidotti, S.; Viterbi, R.; Cerrato, C. The impact of pastoral activities on animal biodiversity in Europe: A systematic review and meta-analysis. J. Nat. Conserv. 2020, 56, 125863. [Google Scholar] [CrossRef]
- Rockström, J.; Williams, J.; Daily, G.; Noble, A.; Matthews, N.; Gordon, L.; Wetterstrand, H.; DeClerck, F.; Shah, M.; Steduto, P.; et al. Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio 2017, 46, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Pretty, J. Intensification for redesigned and sustainable agriculture. Science 2018, 362, 6417. [Google Scholar] [CrossRef]
- Rudel, T.K. The variable paths to sustainable intensification in agriculture. Reg. Environ. Change 2020, 20, 126. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.-O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; van Diemen, R.; et al. (Eds.) IPCC Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems. 2019. Available online: https://www.ipcc.ch/site/assets/uploads/2019/11/SRCCL-Full-Report-Compiled-191128.pdf (accessed on 31 October 2023).
- McKinsey Report. Striking the Balance: Catalyzing a Sustainable Land-Use Transition. 2023, 16p. Available online: https://www.mckinsey.com/industries/agriculture/our-insights/striking-the-balance-catalyzing-a-sustainable-land-use-transition (accessed on 18 November 2023).
- Eisler, M.C.; Lee, M.R.; Tarlton, J.F.; Martin, G.B.; Beddington, J.; Dungait, J.A.; Greathead, H.; Liu, J.; Mathew, S.; Miller, H.; et al. Agriculture: Steps to sustainable livestock. Nature 2014, 507, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, G. Alternative Cereals in Animal Feed. Veterinaria Digital. 2019. Available online: https://www.veterinariadigital.com/en/articulos/alternative-cereals-in-animal-feed/#:~:text=Globally%2C%2037%25%20of%20cereal%20production%20goes%20to%20animal%20protein%20production.,-Graphic%201%3A%20Percentage (accessed on 18 November 2023).
- Pradhan, P.; Lüdeke, M.K.; Reusser, D.E.; Kropp, J.P. Embodied crop calories in animal products. Environ. Res. Lett. 2013, 8, 044044. [Google Scholar] [CrossRef]
- Eisen, M.B.; Brown, P.O. Rapid global phaseout of animal agriculture has the potential to stabilize greenhouse gas levels for 30 years and offset 68 percent of CO2 emissions this century. PLoS Clim. 2022, 1, e0000010. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Aveyard, P.; Garnett, T.; Hall, J.W.; Key, T.J.; Lorimer, J.; Pierrehumbert, R.T.; Scarborough, P.; Springmann, M.; Jebb, S.A. Meat consumption, health, and the environment. Science 2018, 361, eaam5324. [Google Scholar] [CrossRef]
- Graça, J.; Godinho, C.A.; Truninger, M. Reducing meat consumption and following plant-based diets: Current evidence and future directions to inform integrated transitions. Trends Food Sci. Technol. 2019, 91, 380–390. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the anthropocene: The EAT–lancet commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Manzano, P.; Rowntree, J.; Thompson, L.; del Prado, A.; Ederer, P.; Windisch, W.; Lee, M.R.F. Challenges for the balanced attribution of livestock’s environmental impacts: The art of conveying simple messages around complex realities. Anim. Front. 2023, 13, 35–44. [Google Scholar] [CrossRef]
- Cain, M.; Lynch, J.; Allen, M.R.; Fuglestveldt, J.S.; Frame, D.J.; Macey, A.H. Improved calculation of warming-equivalent emissions for short-lived climate pollutants. npj Clim. Atmos. Sci. 2019, 2, 29. [Google Scholar] [CrossRef]
- Smith, M.A.; Cain, M.; Allen, M.R. Further improvement of warming-equivalent emissions calculation. npj Clim. Atmos. Sci. 2021, 4, 19. [Google Scholar] [CrossRef]
- Smetana, S.; Mathys, A.; Knoch, A.; Heinz, V. Meat alternatives: Life cycle assessment of most known meat substitutes. Int. J. Life Cycle Assess. 2015, 20, 1254–1267. [Google Scholar] [CrossRef]
- Mattick, C.S.; Landis, A.E.; Allenby, B.R.; Genovese, N.J. Anticipatory life cycle analysis of in vitro biomass cultivation for cultured meat production in the United States. Environ. Sci. Technol. 2015, 49, 11941–11949. [Google Scholar] [CrossRef]
- Tuomisto, H.L.; Teixeira de Mattos, M.J. Environmental impacts of cultured meat production. Environ. Sci. Technol. 2011, 45, 6117–6123. [Google Scholar] [CrossRef]
- Risner, D.; Kim, Y.; Nguyen, C.; Siegel, J.B.; Spang, E.S. Environmental Impacts of Cultured Meat: A Cradle-to-Gate Life Cycle Assessment. bioXiv 2023. [Google Scholar] [CrossRef]
- Rodríguez Escobar, M.I.; Cadena, E.; Nhu, T.T.; Cooreman-Algoed, M.; De Smet, S.; Dewulf, J. Analysis of the Cultured Meat Production System in Function of Its Environmental Footprint: Current Status, Gaps and Recommendations. Foods 2021, 10, 2941. [Google Scholar] [CrossRef]
- UNEP United Nations Environment Programme. What’s Cooking? An Assessment of the Potential Impacts of Selected Novel Alternatives to Conventional Animal Products; Frontiers 2023; United Nations Environment Programme: Nairobi, Kenya, 2023; p. 72. [Google Scholar]
- Smetana, S.; Ristic, D.; Pleissner, D.; Tuomisto, H.L.; Parniakov, O.; Heinz, V. Meat substitutes: Resource demands and environmental footprints. Resour. Conserv. Recycl. 2023, 190, 106831. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Janzen, H.H.; Little, S.M.; McAllister, T.A.; McGinn, S.M. Life cycle assessment of greenhouse gas emissions from beef production in western Canada: A case study. Agric. Syst. 2010, 103, 371–379. [Google Scholar] [CrossRef]
- Blonk, H.; Kool, A.; Luske, B. Milieueffecten van Nederlandse Consumptie van Eiwitrijke Producten. Gevolgen van Vervanging van Dierlijke Eiwitten Anno 2008. 166p. Available online: https://edepot.wur.nl/117665#:~:text=Gezonder%20eten%20ten%20aanzien%20van,visconsumptie%20buiten%20beschouwing%20is%20gebleven (accessed on 19 January 2024).
- Head, M.; Sevenster, M.; Croezen, H. Life Cycle Impacts of Protein Rich Foods for Superwijzer 2011, 70p. Available online: https://cedelft.eu/wp-content/uploads/sites/2/2021/04/2329_finalreportMHSD_1339077247.pdf (accessed on 19 January 2024).
- Nijdam, D.; Rood, T.; Westhoek, H. The price of protein: Review of land use and carbon footprints from life cycle assessments of animal food products and their substitutes. Food Policy 2012, 37, 760–770. [Google Scholar] [CrossRef]
- Goldstein, B.; Moses, R.; Sammons, N.; Birkved, M. Potential to curb the environmental burdens of American beef consumption using a novel plant-based beef substitute. PLoS ONE 2017, 12, e0189029. [Google Scholar] [CrossRef]
- Heller, M.; Keoleian, G. “Beyond Meat’s Beyond Burger Life Cycle Assessment: A Detailed Comparison between a Plant-Based and an Animal-Based Protein Source. Report No. CSS18-10. Center for Sustainable Systems, University of Michigan 2018, 45 p. Available online: https://css.umich.edu/sites/default/files/publication/CSS18-10.pdf (accessed on 17 July 2023).
- Stanley, P.L.; Rowntree, J.E.; Beede, D.K.; Delonge, M.S.; Hamm, M.W. Impacts of soil carbon sequestration on life cycle greenhouse gas emissions in Midwestern USA beef finishing systems. Agric. Syst. 2018, 162, 249–258. [Google Scholar] [CrossRef]
- Asem-Hiablie, S.; Battagliese, T.; Stackhouse-Lawson, K.R.; Rotz, C.A. A life cycle assessment of the environmental impacts of a beef system in the United States. Int. J. Life Cycle Assess. 2019, 24, 441–455. [Google Scholar] [CrossRef]
- Quantis International. Carbon Footprint Evaluation of Regenerative Grazing at White Oak Pastures. Prepared for General Mills and White Oak Pastures 2019, 34p. Available online: https://blog.whiteoakpastures.com/hubfs/WOP-LCA-Quantis-2019.pdf (accessed on 17 July 2023).
- Rotz, C.A.; Asem-Hiablie, S.; Place, S.; Thoma, G. Environmental footprints of beef cattle production in the United States. Agric. Syst. 2019, 169, 1–13. [Google Scholar] [CrossRef]
- Tuomisto, H.L. The eco-friendly burger: Could cultured meat improve the environmental sustainability of meat products? EMBO Rep. 2019, 20, e47395. [Google Scholar] [CrossRef]
- van Vliet, S.; Kronberg, S.L.; Provenza, F.D. Plant-based meats, human health, and climate change. Front. Sustain. Food Syst. 2020, 4, 128. [Google Scholar] [CrossRef]
- Saget, S.; Port Costa, M.; Santos, C.S.; Vasconcelos, M.; Styles, D.; Williams, M. Comparative life cycle assessment of plant and beef based patties, including carbon opportunity costs. Sustain. Prod. Consum. 2021, 28, 936–952. [Google Scholar] [CrossRef]
- Ledgard, S.; Falconer, S.; Mazzetto, A. Carbon Footprint of New Zealand Beef and Sheep Exported to Different Markets. Report for the Meat Industry Association and Beef+ Lamb New Zealand, RE450/2021/006 2021, 81p. Available online: https://beeflambnz.com/sites/default/files/levies/files/BLNZ_review_report.pdf (accessed on 19 January 2024).
- Tuomisto, H.L.; Allan, S.J.; Ellis, M.J. Prospective life cycle assessment of a bioprocess design for cultured meat production in hollow fiber bioreactors. Sci. Total Environ. 2022, 851, 158051. [Google Scholar] [CrossRef]
- Mazzetto, A.M.; Falconer, S.; Ledgard, S. Carbon footprint of New Zealand beef and sheep meat exported to different markets. Environ. Impact Assess. Rev. 2023, 98, 106946. [Google Scholar] [CrossRef]
- Sinke, P.; Swartz, E.; Sanctorum, H.; van der Giesen, C.; Odegard, I. Ex-ante life cycle assessment of commercial-scale cultivated meat production in 2030. Int. J. Life Cycle Assess. 2023, 28, 234–254. [Google Scholar] [CrossRef]
- Mazzetto, A.; Falconer, S.; Ledgard, S. Mapping the Carbon Footprint of Milk for Dairy Cows; Report for DairyNZ (No. RE450/2020/081); AgResearch: Hamilton, New Zealand, 2021; 22p. [Google Scholar]
- Mazzetto, A.M.; Falconer, S.; Ledgard, S. Mapping the carbon footprint of milk production from cattle: A systematic review. J. Dairy Sci. 2022, 105, 9713–9725. [Google Scholar] [CrossRef]
- Herron, J.; Hennessy, D.; Curran, T.P.; Moloney, A.; O’Brien, D. The simulated environmental impact of incorporating white clover into pasture-based dairy production systems. J. Dairy Sci. 2021, 104, 7902–7918. [Google Scholar] [CrossRef]
- Harris, S.L.; Clark, D.A.; Auldist, M.J.; Waugh, C.D.; Laboyrie, P.G. Optimum white clover content for dairy pastures. Proc. N. Z. Grassl. Assoc. 1997, 59, 29–33. [Google Scholar] [CrossRef]
- McAuliffe, S.; Mee, J.F.; Lewis, E.; Galvin, N.; Hennessy, D. Feeding system effects on dairy cow rumen function and milk production. Animal 2022, 12, 523. [Google Scholar] [CrossRef]
- Clark, D.A.; Harris, S.L. White clover or nitrogen fertiliser for dairying? In White Clover: NZ’s Competitive Edge; Woodfield, D.R., Ed.; NZ Grassland Association Research and Practice Series 6; New Zealand Grassland Association: Dunedin, New Zealand, 1995; pp. 107–114. [Google Scholar] [CrossRef]
- O’Neill, B.; Deighton, M.; O’Loughlin, B.; Mulligan, F.; Boland, T.; O’Donovan, M.; Lewis, E. Effects of a perennial ryegrass diet or total mixed ration diet offered to spring-calving Holstein-Friesian dairy cows on methane emissions, dry matter intake, and milk production. J. Dairy Sci. 2011, 94, 1941–1951. [Google Scholar] [CrossRef]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change Through Livestock: A Global Assessment of Emissions and Mitigation Opportunities. Food and Agriculture Organization of the United Nations (FAO) 2013, 115p. Available online: https://www.fao.org/3/i3437e/i3437e.pdf (accessed on 2 November 2023).
- Gerber, P.J.; Vellinga, T.; Opio, C.; Steinfeld, H. Productivity gains and greenhouse gas intensity in dairy systems. Livest. Sci. 2011, 139, 100–108. [Google Scholar] [CrossRef]
- Lee, M.A.; Davis, A.P.; Chagunda, M.G.; Manning, P. Forage quality declines with rising temperatures, with implications for livestock production and methane emissions. Biogeosciences 2017, 14, 1403–1417. [Google Scholar] [CrossRef]
- Berhanu, Y.; Olav, L.; Nurfeta, A.; Angassa, A.; Aune, J.B. Methane Emissions from Ruminant Livestock in Ethiopia: Promising Forage Species to Reduce CH4 Emissions. Agriculture 2019, 9, 130. [Google Scholar] [CrossRef]
- Pierrehumbert, R.T.; Eshel, G. Climate impact of beef: An analysis considering multiple time scales and production methods without use of global warming potentials. Environ. Res. Lett. 2015, 10, 085002. [Google Scholar] [CrossRef]
- Marlow, H.J.; Hayes, W.K.; Soret, S.; Carter, R.L.; Schwab, E.R.; Sabate, J. Diet and the environment: Does what you eat matter? Am. J. Clin. Nutr. 2009, 89, 1699S–1703S. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, M.; Hoekstra, A.Y. The Green, Blue and Grey Water Footprint of Animals and Animal Products. (Value of Water Research Report 48 2022, No. 48). Unesco-IHE Institute for Water Education. Available online: http://www.unesco-ihe.org/Value-of-Water-Research-Report-Series/Research-Papers (accessed on 28 October 2023).
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J. Availability of disaggregated greenhouse gas emissions from beef cattle production: A systematic review. Environ. Impact Assess. Rev. 2019, 76, 69–78. [Google Scholar] [CrossRef]
- Cusworth, G.; Lorimer, J.; Brice, J.; Garnett, T. Green rebranding: Regenerative agriculture, future-pasts, and the naturalisation of livestock. Trans. Inst. Br. Geogr. 2022, 47, 1009–1027. [Google Scholar] [CrossRef] [PubMed]
- Sexton, A.E.; Garnett, T.; Lorimer, J. Vegan food geographies and the rise of big veganism. Prog. Hum. Geogr. 2022, 46, 605–628. [Google Scholar] [CrossRef]
- Seré, C.; Steinfeld, H. World Livestock Production Systems. FAO Animal Production and Health Paper 127. Food and Agriculture Organization of the United Nations 1996, 58p. Available online: https://hdl.handle.net/10568/21111 (accessed on 3 November 2023).
- Alcock, D.; Hegarty, R.S. Effects of pasture improvement on productivity, gross margin and methane emissions of a grazing sheep enterprise. Int. Congr. Ser. 2006, 1293, 103–106. [Google Scholar] [CrossRef]
- Smith, P.E.; Kelly, A.K.; Kenny, D.A.; Waters, S.M. Enteric methane research and mitigation strategies for pastoral-based beef cattle production systems. Front. Vet. Sci. 2022, 9, 958340. [Google Scholar] [CrossRef]
- Grainger, C.; Clarke, T.; Beauchemin, K.A.; McGinn, S.M.; Eckard, R.J. Supplementation with white cottonseed reduces methane emissions and can profitably increase milk production of dairy cows offered a forage and cereal grain diet. Aust. J. Exp. Agric. 2008, 48, 73–76. [Google Scholar] [CrossRef]
- Winichayakul, S.; Beechey-Gradwell, Z.; Muetzel, S.; Molano, G.; Crowther, T.; Lewis, S.; Xue, H.; Burke, J.; Bryan, G.; Roberts, N.J. In vitro gas production and rumen fermentation profile of fresh and ensiled genetically modified high–metabolizable energy ryegrass. J. Dairy Sci. 2020, 103, 2405–2418. [Google Scholar] [CrossRef] [PubMed]
- Beechey-Gradwell, Z.; Kadam, S.; Bryan, G.; Cooney, L.; Nelson, K.; Richardson, K.; Cookson, R.; Winichayakul, S.; Reid, M.; Anderson, P.; et al. Lolium perenne engineered for elevated leaf lipids exhibits greater energy density in field canopies under defoliation. Field Crops Res. 2022, 275, 108340. [Google Scholar] [CrossRef]
- Waghorn, G.C.; Tavendale, M.H.; Woodfield, D.R. Methanogenesis from forages fed to sheep. Proc. New Zealand Grassl. Assoc. 2002, 64, 167–171. [Google Scholar] [CrossRef]
- Min, B.R.; Solaiman, S.; Waldrip, H.M.; Parker, D.; Todd, R.W.; Brauer, D. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannin mitigation options. Anim. Nutr. 2020, 6, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Ghamkhar, K.; Rochfort, S.; Banik, B.K.; Revell, C. Candidate metabolites for methane mitigation in the forage legume biserrula. Agron. Sustain. Dev. 2018, 38, 30. [Google Scholar] [CrossRef]
- Banik, B.K.; Durmic, Z.; Erskine, W.; Revell, C. Anti-methanogenic advantage of biserrula (Biserrula pelecinus) over subterranean clover (Trifolium subterraneum) from in vitro fermentation is maintained across growth stages and cutting treatments. Crop Pasture Sci. 2019, 70, 263–272. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technology. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Woodward, S.L.; Waghorn, G.C.; Laboyrie, P.G. Condensed tannins in birdsfoot trefoil (Lotus corniculatus) reduce methane emissions from dairy cows. Proc. N. Z. Soc. Anim. Prod. 2004, 64, 160–164. [Google Scholar]
- Aerts, R.J.; McNabb, W.C.; Molan, A.; Brand, A.; Barry, T.N.; Peters, J.S. Condensed tannins from Lotus corniculatus and Lotus pedunculatus exert different effects on the in vitro rumen degradation of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) protein. J. Sci. Food Agric. 1999, 79, 79–85. [Google Scholar] [CrossRef]
- Lees, G.L.; Gruber, M.Y.; Suttill, N.H. Condensed tannins in sainfoin. II. Occurrence and changes during leaf development. Can. J. Bot. 1995, 73, 1540–1547. [Google Scholar] [CrossRef]
- Caradus, J.R.; Voisey, C.R.; Cousin, G.R.; Kaur, R.; Woodfield, D.R.; Blanc, A.; Roldan, M.B. The hunt for the “holy grail”: Condensed tannins in the perennial forage legume white clover (Trifolium repens L.). Grass Forage Sci. 2022, 77, 111–123. [Google Scholar] [CrossRef]
- Roldan, M.B.; Cousins, G.; Muetzel, S.; Zeller, W.E.; Fraser, K.; Salminen, J.-P.; Blanca, A.; Kaur, R.; Richardson, K.; Maher, D.; et al. Condensed tannins in white clover (Trifolium repens) foliar tissues expressing the transcription factor TaMYB14-1 bind to forage protein and reduce ammonia and methane emissions in vitro. Front. Plant Sci. 2022, 12, 777354. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.K.; Hegarty, R.S.; Cowie, A. Meta-analysis quantifying the potential of dietary additives and rumen modifiers for methane mitigation in ruminant production systems. Anim. Nutr. 2021, 7, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Honan, M.; Feng, X.; Tricarico, J.M.; Kebreab, E. Feed additives as a strategic approach to reduce enteric methane production in cattle: Modes of action, effectiveness and safety. Anim. Prod. Sci. 2021, 62, 1303–1317. [Google Scholar] [CrossRef]
- Króliczewska, B.; Pecka-Kiełb, E.; Bujok, J. Strategies used to reduce methane emissions from ruminants: Controversies and issues. Agriculture 2023, 13, 602. [Google Scholar] [CrossRef]
- Roque, B.M.; Venegas, M.; Kinley, R.D.; de Nys, R.; Duarte, T.L.; Yang, X.; Kebreab, E. Red seaweed (Asparagopsis taxiformis) supplementation reduces enteric methane by over 80 percent in beef steers. PLoS ONE 2021, 16, e0247820. [Google Scholar] [CrossRef] [PubMed]
- Glasson, C.R.; Kinley, R.D.; de Nys, R.; King, N.; Adams, S.L.; Packer, M.A.; Svenson, J.; Eason, C.T.; Magnusson, M. Benefits and risks of including the bromoform containing seaweed Asparagopsis in feed for the reduction of methane production from ruminants. Algal Res. 2022, 64, 102673. [Google Scholar] [CrossRef]
- Faust, R.A. Toxicity Summary for Bromoform. Prepared for Oak Ridge Reservation Environmental Restoration Program 1995. 11p. Available online: https://web.archive.org/web/20060929185136/http://cira.ornl.gov/documents/BROMOFRM.pdf (accessed on 4 November 2023).
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Hayes, M. Potential of seaweeds to mitigate production of greenhouse gases during production of ruminant proteins. Glob. Chall. 2023, 7, 2200145. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Belanche, A.; Hueso, R.; Ramos-Morales, E.; Salwen, J.K.; Kebreab, E.; Yáñez-Ruiz, D.R. In vivo rumen microbial degradation of bromoform and the impact on rumen fermentation. In Proceedings of the 8th Annual Global Greenhouse Gas & Animal Agriculture Conference, Orlando, FL, USA, 5–10 June 2022; p. 291. [Google Scholar]
- Eason, C.T.; Fennessy, P. Methane reduction, health and regulatory considerations regarding Asparagopsis and bromoform for ruminants. N. Z. J. Agric. Res. 2023, 30, 2248948. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006, 84, 1489–1496. [Google Scholar] [CrossRef]
- Clark, H.; Kelliher, F.; Pinares-Patiño, C. Reducing CH4 Emissions from Grazing Ruminants in New Zealand: Challenges and Opportunities. Anim. Biosci. 2011, 24, 295–302. [Google Scholar] [CrossRef]
- Riede, S.; Boguhn, J.; Breves, G. Studies on potential effects of fumaric acid on rumen microbial fermentation, methane production and microbial community. Arch. Anim. Nutr. 2013, 67, 368–380. [Google Scholar] [CrossRef]
- Wood, T.A.; Wallace, R.J.; Rowe, A.; Price, J.; Yáñez-Ruiz, D.R.; Murray, P.; Newbold, C.J. Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions. Anim. Feed Sci. Technol. 2009, 152, 62–71. [Google Scholar] [CrossRef]
- Sun, K.; Liu, H.; Fan, H.; Liu, T.; Zheng, C. Research progress on the application of feed additives in ruminal methane emission reduction: A review. PeerJ 2021, 9, e11151. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, G.J.; Thomson, B.C.; Muir, P.D. Effect of management change on methane output within a sheep flock. Proc. N. Z. Soc. Anim. Prod. 2009, 69, 170–173. [Google Scholar]
- Pickering, N.K.; Oddy, V.H.; Basarab, J.; Cammack, K.; Hayes, B.; Hegarty, R.S.; Lassen, J.; McEwan, J.C.; Miller, S.; Pinares-Patiño, C.S.; et al. Animal board invited review: Genetic possibilities to reduce enteric methane emissions from ruminants. Animal 2015, 9, 1431–1440. [Google Scholar] [CrossRef]
- Fennessy, P.F.; Byrne, T.J.; Proctor, L.E.; Amer, P.R. The potential impact of breeding strategies to reduce methane output from beef cattle. Anim. Prod. Sci. 2019, 59, 1598–1610. [Google Scholar] [CrossRef]
- Rowe, S.J.; Hickey, S.M.; Johnson, P.L.; Bilton, T.P.; Jonker, A.; Bain, W.; Veenvliet, B.; Pilel, G.; Bryson, B.; Knowler, K. The contribution animal breeding can make to industry carbon neutrality goals. Proc. Assoc. Adv. Anim. Breed. Genet. 2021, 24, 15–18. [Google Scholar]
- Pinares-Patiño, C.S.; Hickey, S.M.; Young, E.A.; Dodds, K.G.; MacLean, S.; Molano, G.; Sandoval, E.; Kjestrup, H.; Harland, R.; Hunt, C.; et al. Heritability estimates of methane emissions from sheep. Animal 2013, 7, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Hickey, S.M.; Bain, W.E.; Bilton, T.P.; Greer, G.J.; Elmes, S.; Bryson, B.; Pinares-Patiño, C.S.; Wing, J.; Jonker, A.; Young, E.A.; et al. Impact of breeding for reduced methane emissions in New Zealand sheep on maternal and health traits. Front. Genet. 2022, 13, 910413. [Google Scholar] [CrossRef] [PubMed]
- Kobayashr, Y. Abatement of methane production from ruminants: Trends in the manipulation of rumen fermentation. Asian-Australas. J. Anim. Sci. 2010, 23, 410–416. [Google Scholar] [CrossRef]
- Wedlock, D.N.; Janssen, P.H.; Leahy, S.C.; Shu, D.; Buddle, B.M. Progress in the development of vaccines against rumen methanogens. Animal 2013, 7, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, 2s–16s. [Google Scholar] [CrossRef]
- Baca-González, V.; Asensio-Calavia, P.; González-Acosta, S.; Pérez de la Lastra, J.M.; Morales de la Nuez, A. Are Vaccines the Solution for Methane Emissions from Ruminants? A Systematic Review. Vaccines 2020, 8, 460. [Google Scholar] [CrossRef]
- Rolfe, J. Economics of reducing methane emissions from beef cattle in extensive grazing systems in Queensland. Rangel. J. 2010, 32, 197–204. [Google Scholar] [CrossRef]
- Moot, D.J.; Davison, R. Changes in New Zealand red meat production over the past 30 yr. Anim. Front. 2021, 11, 26–31. [Google Scholar] [CrossRef]
- Henry, B.; Charmley, E.; Eckard, R.; Gaughan, J.B.; Hegarty, R. Livestock production in a changing climate: Adaptation and mitigation research in Australia. Crop Pasture Sci. 2012, 63, 191–202. [Google Scholar] [CrossRef]
- Harty, M.A.; Forrestal, P.J.; Watson, C.J.; McGeough, K.L.; Carolan, R.; Elliot, C.; Krol, D.; Laughlin, R.J.; Richards, K.G.; Lanigan, G.J. Reducing nitrous oxide emissions by changing N fertiliser use from calcium ammonium nitrate (CAN) to urea based formulations. Sci. Total Environ. 2016, 563, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Ridoutt, B.G.; Sanguansri, P.; Freer, M.; Harper, G.S. Water footprint of livestock: Comparison of six geographically defined beef production systems. Int. J. Life Cycle Assess. 2012, 17, 165–175. [Google Scholar] [CrossRef]
- Heinke, J.; Lannerstad, M.; Gerten, D.; Havlík, P.; Herrero, M.; Notenbaert, A.M.O.; Hoff, H.; Muller, C. Water use in global livestock production—Opportunities and constraints for increasing water productivity. Water Resour. Res. 2020, 56, e2019WR026995. [Google Scholar] [CrossRef]
- Rys, G.J.; Gibbs, J.; Clark, D.A.; Lambert, G.; Clark, H.; Newman, M. Productivity, changes and resilience in New Zealand grassland agriculture over the last three decades. In Resilient Pastures; Douglas, G.B., Ed.; Grassland Research and Practice Series; New Zealand Grassland Association: Dunedin, New Zealand, 2021; Volume 17, pp. 37–54. [Google Scholar] [CrossRef]
- Land Air Water Aotearoa 2007. New Zealand Water Quality—International Comparisons. Available online: https://www.lawa.org.nz/explore-data/water-quantity/water-quantity-national-picture/international-comparisons/ (accessed on 20 December 2023).
- Lawrence, J.; Blackett, P.; Cradock-Henry, N.A. Cascading climate change impacts and implications. Clim. Risk Manag. 2020, 29, 100234. [Google Scholar] [CrossRef]
- Mfe 2021. Fresh Water. Available online: https://environment.govt.nz/publications/statement-of-intent-2008-2011/operating-intentions/fresh-water/ (accessed on 16 December 2023).
- Prosser, I.P.; Chiew, F.H.S.; Stafford Smith, M. Adapting water management to climate change in the Murray–Darling Basin, Australia. Water 2021, 13, 2504. [Google Scholar] [CrossRef]
- Monaghan, R.M.; de Klein, C.A.M.; Muirhead, R.W. Prioritisation of farm scale remediation efforts for reducing losses of nutrients and faecal indicator organisms to waterways: A case study of New Zealand dairy farming. J. Environ. Manag. 2008, 87, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Quin, J.M.; Wilcock, R.J.; Monaghan, R.M.; McDowell, R.W.; Journeaux, P.R. Grassland farming and water quality in New Zealand. Ir. J. Agri-Environ. Res. 2009, 7, 69–88. [Google Scholar]
- Our Land and Water 2023. Study Reveals the Size of the Job to Achieve Water Quality Goals. Available online: https://ourlandandwater.nz/news/study-reveals-the-size-of-the-job-to-achieve-water-quality-goals/ (accessed on 20 December 2023).
- Economist. Dairy Farming Is Polluting New Zealand’s Water. 2017. Available online: http://www.economist.com/news/asia/21731435-government-data-suggests-60-rivers-and-lakes-are-unswimmable-dairy-farming-polluting-new?frsc=dg%7Ce (accessed on 16 December 2023).
- Davies-Colley, R.J. Land use and water quality in New Zealand—An overview. Water Air Soil Pollut. 2009, 162, 32–35. [Google Scholar]
- Leip, A.; Wollgast, J.; Kugelberg, S.; Costa Leite, J.; Maas, R.J.M.; Mason, K.E.; Sutton, M.A. (Eds.) Appetite for Change: Food System Options for Nitrogen, Environment & Health, 2nd ed.; European Nitrogen Assessment Special Report on Nitrogen & Food; UK Centre for Ecology and Hydrology: Edinburgh, UK, 2023; 154p, Available online: https://www.clrtap-tfrn.org (accessed on 16 December 2023).
- de Klein, C.A.M.; Sherlock, R.R.; Cameron, K.C.; van der Weerden, T.J. Nitrous oxide emissions from agricultural soils in New Zealand—A review of current knowledge and directions for future research. J. R. Soc. N. Z. 2001, 31, 543–574. [Google Scholar] [CrossRef]
- Saggar, S. Estimation of nitrous oxide emission from ecosystems and its mitigation technologies. Agric. Ecosyst. Environ. 2010, 136, 189–365. [Google Scholar] [CrossRef]
- Moorby, J.M. Relationship between the composition of fresh grass-based diets and the excretion of dietary nitrogen from dairy cows. In Proceedings of the 25th General Meeting of the European Grassland Federation, Aberystwyth, UK, 7–11 September 2014; Volume 19, pp. 686–689. [Google Scholar]
- Vibart, R.E.; Tavendale, M.; Otter, D.; Schwendel, B.H.; Lowe, K.; Gregorini, P.; Pacheco, D. Milk production and composition, nitrogen utilization, and grazing behavior of late-lactation dairy cows as affected by time of allocation of a fresh strip of pasture. J. Dairy Sci. 2017, 100, 5305–5318. [Google Scholar] [CrossRef] [PubMed]
- van der Weerden, T.J.; Luo, J.F.; de Klein, C.A.M.; Hoogendoorn, C.J.; Littlejohn, R.P.; Rys, G.J. Disaggregating nitrous oxide emission factors for ruminant urine and dung deposited onto pastoral soils. Agric. Ecosyst. Environ. 2011, 141, 426–436. [Google Scholar] [CrossRef]
- Theodoridou, K.; Aufrere, J.; Andueza, D.; Morvan, A.; Picard, F.; Stringano, E.; Pourrat, J.; Mueller-Harvey, I.; Baumont, R. Effect of plant development during first and second growth cycle on chemical composition, condensed tannins and nutritive value of three sainfoin (Onobrychis viciifolia) varieties and lucerne. Grass Forage Sci. 2010, 66, 402–414. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, M.; Jiang, L.; Guan, L. Could natural phytochemicals be used to reduce nitrogen excretion and excreta-derived N2O emissions from ruminants? J. Anim. Sci. Biotechnol. 2023, 14, 140. [Google Scholar] [CrossRef] [PubMed]
- Larned, S.T.; Snelder, T.; Unwin, M.J.; McBride, G.B. Water quality in New Zealand rivers: Current state and trends. N. Z. J. Mar. Freshw. Res. 2016, 50, 389–417. [Google Scholar] [CrossRef]
- Carran, R.A.; Clough, T. Environmental effects of nitrogen in pastoral agriculture. In White Clover: New Zealand’s Competitive Advantage; Woodfield, D.R., Ed.; Agronomy Society of New Zealand Special Publication No. 11 and New Zealand Grassland Association Research and Practice Series 6; New Zealand Grassland Association: Dunedin, New Zealand, 1996; pp. 99–102. [Google Scholar] [CrossRef]
- McInerny, J.P. Economic aspects of grassland production and utilization. In Grass; Hopkins, A., Ed.; Its Production and Utilization; Blackwell Science: Oxford, UK, 2000; pp. 394–428. [Google Scholar]
- Parsons, A.J.; Thornley, J.H.M.; Rasmussen, S.; Rowarth, J.S. Some clarification of the impacts of grassland intensification on food production, nitrogen release, greenhouse gas emissions and carbon sequestration: Using the example of New Zealand. CAB Rev. 2016, 11, 19. [Google Scholar] [CrossRef]
- Chapman, D.; Parsons, T. Doing more for the environment with lower inputs in pasture-based livestock systems: Does this always lead to lower outputs. In Doing more with less. In Proceedings of the 18th Australian Society of Agronomy Conference, Ballarat, Australia, 24–28 September 2017; pp. 24–28. Available online: https://www.agronomyaustraliaproceedings.org/images/sampledata/2017/184_ASA2017_Chapman_David_Final.pdf (accessed on 19 January 2024).
- Beukes, P.C.; Scarsbrook, M.R.; Gregorini, P.; Romera, A.J.; Clark, D.A.; Catto, W. The relationship between milk production and farm-gate nitrogen surplus for the Waikato region, New Zealand. J. Environ. Manag. 2012, 93, 44–51. [Google Scholar] [CrossRef]
- de Klein, C.A.M.; Monaghan RMAlfaro, M.; Gourley, C.J.P.; Oenema, O.; Powell, M. Nitrogen performance indicators for dairy production systems. Soil Res. 2017, 55, 479–488. [Google Scholar] [CrossRef]
- Pinxterhuis, J.B.; Edwards, J.P. Comparing nitrogen management on dairy farms—Canterbury case studies. J. N. Z. Grassl. 2018, 80, 201–206. [Google Scholar] [CrossRef]
- Stott, K.J.; Gourley, C.J.P. Intensification, nitrogen use and recovery in grazing-based dairy systems. Agric. Syst. 2016, 144, 101–112. [Google Scholar] [CrossRef]
- Macdonald, K.A.; Penno, J.W.; Lancaster, J.A.S.; Bryant, A.M.; Kidd, J.M.; Roche, J.R. Production and economic responses to intensification of pasture-based dairy production systems. J. Dairy Sci. 2017, 100, 6602–6619. [Google Scholar] [CrossRef] [PubMed]
- Hedley, P.; Kolver, E.; Glassey, C.; Thorrold, B.; van Bysterveldt, A.; Roche, J.; Macdonald, K. Achieving high performance from a range of farm systems. In Proceedings of the 4th Dairy3 Conference, Hamilton, New Zealand, 3–5 April 2006; pp. 147–166. [Google Scholar]
- Chapman, D.; Macdonald, K.; Glassey, C.; Pinxterhuis, I.; Edwards, P.; Beukes, P. Relationships between nitrogen inputs, outputs in product, and surpluses in New Zealand dairy systems. In Proceedings of the 8th Australasian Dairy Science Symposium; 2018; pp. 187–190. [Google Scholar]
- Clark, D.A.; Macdonald, K.A.; Glassey, C.B.; Roach, C.G.; Woodward, S.L.; Griffiths, W.M.; Neal, M.B.; Shepherd, M.A. Production and profit of current and future dairy systems using differing nitrate leaching mitigation methods: The Pastoral 21 experience in Waikato. N. Z. J. Agric. Res. 2020, 63, 505–528. [Google Scholar] [CrossRef]
- Chapman, D.F.; Dalley, D.E.; Edwards, G.R.; Cameron, K.C.; Malcolm, B.J.; Clement, A.; Romera, A.J.; Pinxterhuis, J.B.; Beukes, P.C.; Di, H.J.; et al. Production, profit and nitrogen flows in irrigated dairy systems representing different industry development pathways: The Pastoral 21 experience in Canterbury. N. Z. J. Agric. Res. 2021, 64, 3–35. [Google Scholar] [CrossRef]
- Huebsch, M.; Horan, B.; Blum, P.; Richards, K.G.; Grant, J.; Fenton, O. Impact of agronomic practices of an intensive dairy farm on nitrogen concentrations in a karst aquifer in Ireland. Agric. Ecosyst. Environ. 2013, 179, 187–199. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Penno, J.W.; Sprosen, M.S. Nitrogen inputs and losses from clover/grass pastures grazed by dairy cows, as affected by nitrogen fertiliser application. J. Agric. Sci. Camb. 1999, 132, 215–225. [Google Scholar] [CrossRef]
- Rotz, C.A.; Taube, F.; Russelle, M.P.; Oenema, J.; Sanderson, M.A.; Wachendorf, M. Whole-farm perspectives of nutrient flows in grassland agriculture. Crop Sci. 2005, 45, 2139–2159. [Google Scholar] [CrossRef]
- Krol, D.J.; Forrestal, P.J.; Wall, D.; Lanigan, G.J.; Sanz-Gomez, J.; Richards, K.G. Nitrogen fertilisers with urease inhibitors reduce nitrous oxide and ammonia losses, while retaining yield in temperate grassland. Sci. Total Environ. 2020, 725, 138329. [Google Scholar] [CrossRef] [PubMed]
- Edmeades, D.; McBride, R. An assessment of the agronomic effectiveness of N-(n-butyl) thiophosphoric triamide (nBTPT)—treated urea on the production of clover-based pastures, pastures, grasses and crops. J. N. Z. Grassl. 2023, 85, 149–153. [Google Scholar] [CrossRef]
- Carrazco, A.V.; Peterson, C.B.; Zhao, Y.; Pan, Y.; McGlone, J.J.; DePeters, E.J.; Mitloehner, F.M. The Impact of Essential Oil Feed Supplementation on Enteric Gas Emissions and Production Parameters from Dairy Cattle. Sustainability 2020, 12, 10347. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Luo, J.; Sprosen, M.S.; Wyatt, J.B.; Balvert, S.F.; Lindsey, S.B. Effects of the nitrification inhibitor dicyandiamide (DCD) on pasture production, nitrous oxide emissions and nitrate leaching in Waikato, New Zealand. N. Z. J. Agric. Res. 2014, 57, 294–315. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Menneer, J.C.; Dexter, M.M.; Kear, M.J.; Lindsey, S.; Peters, J.S.; Pacheco, D. A novel concept to reduce nitrogen losses from grazed pastures by administering soil nitrogen process inhibitors to ruminant animals: A study with sheep. Agric. Ecosyst. Environ. 2008, 125, 148–158. [Google Scholar] [CrossRef]
- Chibuike, G.; Saggar, S.; Palmada, T.; Luo, J. The persistence and efficacy of nitrification inhibitors to mitigate nitrous oxide emissions from New Zealand pasture soils amended with urine. Geoderma Reg. 2022, 30, e00541. [Google Scholar] [CrossRef]
- Abberton, M.T.; Marshall, A.H.; Humphreys, M.W.; Macduff, J.H.; Collins, R.P.; Marley, C.L. Genetic improvement of forage species to reduce the environmental impact of temperate livestock grazing systems. Adv. Agron. 2008, 98, 311–355. [Google Scholar] [CrossRef]
- Hill, J.O.; Simpson, R.J.; Wood, J.T.; Moore, A.D.; Chapman, D.F. The phosphorus and nitrogen requirements of temperate pasture species and their influence on grassland botanical composition. Aust. J. Agric. Res. 2005, 56, 1027–1039. [Google Scholar] [CrossRef]
- Lambert, M.G.; Litherland, A.J. A practitioner’s guide to pasture quality. Proc. N. Z. Grassl. Assoc. 2000, 62, 111–115. [Google Scholar] [CrossRef]
- Gardiner, C.A.; Clough, T.J.; Cameron, K.C.; Di, H.J.; Edwards, G.R.; De Klein, C.A. Potential for forage diet manipulation in New Zealand pasture ecosystems to mitigate ruminant urine derived N2O emissions: A review. N. Z. J. Agric. Res. 2016, 59, 301–317. [Google Scholar] [CrossRef]
- Podolyan, A.; Di, H.J.; Cameron, K.C. Effect of plantain on nitrous oxide emissions and soil nitrification rate in pasture soil under a simulated urine patch in Canterbury, New Zealand. J. Soils Sediments 2020, 20, 1468–1479. [Google Scholar] [CrossRef]
- Pembleton, K.G.; Tozer, K.N.; Edwards, G.R.; Jacobs, J.L.; Turner, L.R. Simple versus diverse pastures: Opportunities and challenges in dairy systems. Anim. Prod. Sci. 2015, 55, 893–901. [Google Scholar] [CrossRef]
- Box, L.A.; Edwards, G.R.; Bryant, R.H. Milk production and urinary nitrogen excretion of dairy cows grazing plantain in early and late lactation. N. Z. J. Agric. Res. 2017, 60, 470–482. [Google Scholar] [CrossRef]
- Judson, H.G.; Fraser, P.M.; Peterson, M.E.; Edwards, G.R. Specific genotypes of plantain (Plantago lanceolata) vary in their impact on sheep urine volume and nitrification in the urine patch. J. N. Z. Grassl. 2018, 17, 125–128. [Google Scholar] [CrossRef]
- Carlton, A.J.; Cameron, K.C.; Di, H.J.; Edwards, G.R.; Clough, T.J. Nitrate leaching losses are lower from ryegrass/white clover forages containing plantain than from ryegrass/white clover forages under different irrigation. N. Z. J. Agric. Res. 2019, 62, 150–172. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Navarrete, S.; Horne, D.J.; Donaghy, D.J.; Kemp, P.D. Effect of plantain content in ryegrass-based dairy pastures on nitrate leaching and key components of the nitrogen cycle. In Adaptive Strategies for Future Farming; Christensen, C.L., Horne, D.J., Singh, R., Eds.; Occasional Report; Farmed Landscapes Research Centre, Massey University: Palmerston North, New Zealand, 2022; Volume 34, 7p, Available online: http://flrc.massey.ac.nz/publications.html (accessed on 19 January 2024).
- Herath, H.M.G.P.; Thomas, C.; McMillan, N.; Woods, R.P.; Bryant, R.H.; Al-Marashdeh, O. Incorporating plantain into ryegrass-white clover mixed sward for and economically and environmentally sustainable dairy system: Year one of a farm system study. J. N. Z. Grassl. 2023, 85, 321–329. [Google Scholar] [CrossRef]
- Allard, V.; Soussana, J.F.; Falcimagne, R.; Berbigier, P.; Bonnefond, J.M.; Ceschia, E.; D’hour, P.; Hénault, C.; Laville, P.; Martin, C.; et al. The role of grazing management for the net biome productivity and greenhouse gas budget (CO2, N2O and CH4) of semi-natural grassland. Agric. Ecosyst. Environ. 2007, 121, 47–58. [Google Scholar] [CrossRef]
- Teague, W.R.; Apfelbaum, S.; Lal, R.; Kreuter, U.P.; Rowntree, J.; Davies, C.A.; Conser, R.; Rasmussen, M.; Hatfield, J.; Wang, T.; et al. The role of ruminants in reducing agriculture’s carbon footprint in North America. J. Soil Water Conserv. 2016, 71, 156–164. [Google Scholar] [CrossRef]
- Delgado, J.A.; Groffman, P.M.; Nearing, M.A.; Goddard, T.; Reicosky, D.; Lal, R.; Kitchen, N.R.; Rice, C.W.; Towery, D.; Salon, P. Conservation practices to mitigate and adapt to climate change. J. Soil Water Conserv. 2011, 66, 118A–129A. [Google Scholar] [CrossRef]
- Briske, D.D.; Bestelmeyer, B.T.; Joel, R. Brown J. Savory’s unsubstantiated claims should not be confused with multi-paddock grazing. Rangelands 2014, 36, 39–42. [Google Scholar] [CrossRef]
- Tate, K.R.; Wilde, R.H.; Giltrap, D.J.; Baisden, W.T.; Saggar, S.; Trustram, N.A.; Scott, N.A.; Barton, J.P. Soil organic carbon stocks and flows in New Zealand: System development, measurement and modelling. Can. J. Soil Sci. 2005, 85, 481–489. [Google Scholar] [CrossRef]
- Goh, K.M. Carbon sequestration and stabilization in soils: Implications for soil productivity and climate change. Soil Sci. Plant Nutr. 2004, 50, 467–476. [Google Scholar] [CrossRef]
- NZAGGRC Soil Carbon Science. 2023. Available online: https://www.agmatters.nz/goals/maintain-soil-carbon/soil-carbon-science/https://www.agmatters.nz/goals/maintain-soil-carbon/soil-carbon-science/ (accessed on 18 November 2023).
- Whitehead, D.; Schipper, L.A.; Pronger, J.; Moinet, G.Y.; Mudge, P.L.; Pereira, R.C.; Kirschbaum, M.U.; McNally, S.R.; Beare, M.H.; Camps-Arbestain, M. Management practices to reduce losses or increase soil carbon stocks in temperate grazed grasslands: New Zealand as a case study. Agric. Ecosyst. Environ. 2018, 265, 432–443. [Google Scholar] [CrossRef]
- Minasny, B.; Malone, B.P.; McBratney, A.B.; Angers, D.A.; Arrouays, D.; Chambers, A.; Chaplot, V.; Chen, Z.S.; Cheng, K.; Das, B.S.; et al. Soil carbon 4 per mille. Geoderma 2017, 292, 59–86. [Google Scholar] [CrossRef]
- Minasny, B.; Arrouays, D.; McBratney, A.B.; Angers, D.A.; Chambers, A.; Chaplot, V.; Chen, Z.S.; Cheng, K.; Das, B.S.; Field, D.J.; et al. Rejoinder to Comments on Minasny et al. Soil carbon 4 per mille. Geoderma 292, 59–86. Geoderma 2018, 309, 124–129. [Google Scholar] [CrossRef]
- Baveye, P.C.; Berthelin, J.; Tessier, D.; Lemaire, G. The “4 per 1000” initiative: A credibility issue for the soil science community? Geoderma 2018, 309, 118–123. [Google Scholar] [CrossRef]
- Schipper, L.A.; Mudge, P.L.; Kirschbaum, M.U.F.; Hedley, C.B.; Golubiewski, N.E.; Smaill, S.E.; Kelliher, F.M. 2017. A review of soil carbon change in New Zealand’s grazed grasslands. N. Z. J. Agric. Res. 2017, 60, 93–118. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D. Soil carbon sequestration in prairie grasslands increased by chronic nitrogen addition. Ecology 2012, 93, 2030–2036. [Google Scholar] [CrossRef] [PubMed]
- Parsons, A.J.; Rowarth, J.S.; Newton, P.C.D. Managing pasture for animals and soil carbon. Proc. N. Z. Grassl. Assoc. 2009, 71, 77–84. [Google Scholar] [CrossRef]
- Parsons, A.J.; Thornley, J.H.M.; Newton, P.C.D.; Rasmussen, S.; Rowarth, J.S. Soil carbon dynamics: The effects of nitrogen input, intake demand and off-take by animals. Sci. Total Environ. 2013, 465, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Schipper, L.A.; Dodd, M.B.; Fisk, L.M.; Power, I.L.; Parenzee, J.; Arnold, G. Trends in soil carbon and nutrients of hill-country pastures receiving different phosphorus fertilizer loadings for 20 years. Biogeochemistry 2011, 104, 35–48. [Google Scholar] [CrossRef]
- Trost, B.; Prochnow, A.; Drastig, K.; Meyer-Aurich, A.; Ellmer, F.; Baumecker, M. Irrigation, soil organic carbon and N2O emissions: A review. Agron. Sustain. Dev. 2013, 33, 733–749. [Google Scholar] [CrossRef]
- Condron, L.M.; Hopkins, D.W.; Gregorich, E.G.; Black, A.; Wakelin, S.A. Long-term irrigation effects on soil organic matter under temperate grazed pasture. Eur. J. Soil Sci. 2014, 65, 741–750. [Google Scholar] [CrossRef]
- Parsons, A.J. Modelling Management Manipulations Using the HPM. New Zealand Agricultural Greenhouse Gas Research Centre Annual Report 2017. pp. 64–66. Available online: https://www.nzagrc.org.nz/assets/Publications/NZAGRC-2017-Annual-Report.pdf (accessed on 18 November 2023).
- Campbell, D.I.; Wall, A.M.; Nieveen, J.P.; Schipper, L.A. Variations in CO2 exchange for dairy farms with year-round rotational grazing in drained peatlands. Agric. Ecosyst. Environ. 2015, 202, 68–78. [Google Scholar] [CrossRef]
- Maillard, E.; Angers, D.A. Animal manure application and soil organic carbon stocks: A meta-analysis. Glob. Change Biol. 2014, 20, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, S.; Wall, A.M.; Mudge, P.L.; Troughton, B.; Campbell, D.I.; Pronger, J.; Joshi, C.; Schipper, L.A. The carbon balance of temperate grasslands part I: The impact of increased species diversity. Agric. Ecosyst. Environ. 2017, 239, 310–323. [Google Scholar] [CrossRef]
- Conant, R.T.; Cerri, C.E.P.; Osborne, B.B.; Paustian, K. Grassland management impacts on soil carbon stocks: A new synthesis. Ecol. Appl. 2017, 27, 662–668. [Google Scholar] [CrossRef]
- Hoglund, J.H. Grazing intensity and soil nitrogen accumulation. Proc. N. Z. Grassl. Assoc. 1985, 46, 65–69. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Chen, W.Q.; Huang, D.; Liu, N.; Zhang, Y.; Badgery, W.B.; Wang, X.; Shen, Y. Improved grazing management may increase soil carbon sequestration in temperate steppe. Sci. Rep. 2015, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.F.; Askari, M.S.; Holden, N.M. Grassland soil carbon and nitrogen stocks under temperate livestock grazing. Soil Res. 2015, 53, 485–493. [Google Scholar] [CrossRef]
- McSherry, M.E.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Change Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- van der Cruijsen, K.; Al Hassan, M.; van Erven, G.; Dolstra, O.; Trindade, L.M. Breeding targets to improve biomass quality in Miscanthus. Molecules 2021, 26, 254. [Google Scholar] [CrossRef]
- Russell, J.R.; Bisinger, J.J. Forages and Pastures Symposium: Improving soil health and productivity on grasslands using managed grazing of livestock. J. Anim. Sci. 2015, 93, 2626–2640. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F.; et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Hedley, M.J.; Beare, M.H.; Calvelo Pereira, R.; McNally, S.R.; Lawrence Smith, E.J.; Tregurtha, C.S.; Osborne, M.A.; Gillespie, R.N.; Van der Klei, G.; Thomas, S.M. Where, when and how—Practise guidelines for successful introduction of full inversion tillage to increase soil carbon stocks under pasture. In Nutrient Management in Farmed Landscapes; Christensen, C.L., Horne, D.J., Singh, R., Eds.; Occasional Report No. 33; Farmed Landscapes Research Centre, Massey University: Palmerston North, New Zealand, 2020; 9p, Available online: https://flrc.massey.ac.nz/workshops/20/Manuscripts/Paper_Hedley_2020.pdf (accessed on 18 November 2023).
- Schon, N.L.; Mackay, A.D.; Gray, R.A.; Dodd, M.B.; van Koten, C. Quantifying dung carbon incorporation by earthworms in pasture soils. Eur. J. Soil Sci. 2015, 66, 348–358. [Google Scholar] [CrossRef]
- Lubbers, I.M.; van Groenigen, K.J.; Fonte, S.J.; Six, J.; Brussaard, L.; van Groenigen, J.W. Greenhouse-gas emissions from soils increased by earthworms. Nat. Clim. Change 2013, 3, 187–194. [Google Scholar] [CrossRef]
- Nervo, B.; Tocco, C.; Caprio, E.; Palestrini, C.; Rolando, A. The effects of body mass on dung removal efficiency in dung beetles. PLoS ONE 2014, 9, e107699. [Google Scholar] [CrossRef]
- Paynter, Q.; Fowler, S.; Forgie, S. Evaluation of the Effectiveness of Dung Beetles in Improving the Environmental Health of Land and Rivers within Tairawhiti (Gisborne District). Contract Report: LC3284. Envirolink Grant: 1828-GSDC147. Landcare Research New Zealand Ltd. and Gisborne District Council, 2018. 45p. Available online: https://envirolink.govt.nz/assets/Envirolink/Reports/1828-GSDC147-Evaluation-of-the-effectiveness-of-dung-beetles-in-improving-the-environmental-health-of-land-and-rivers-within-Tairawhiti.pdf (accessed on 18 November 2023).
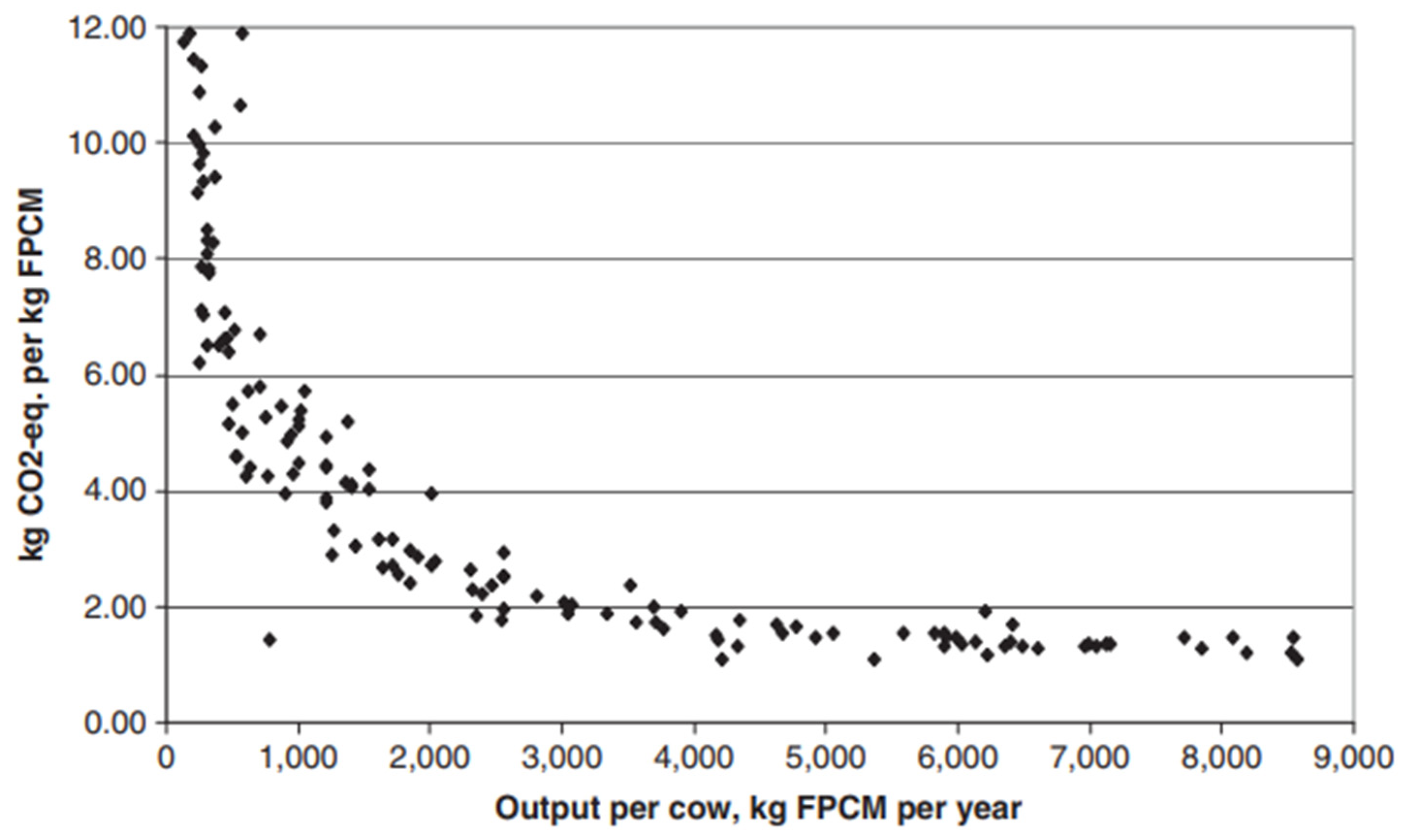
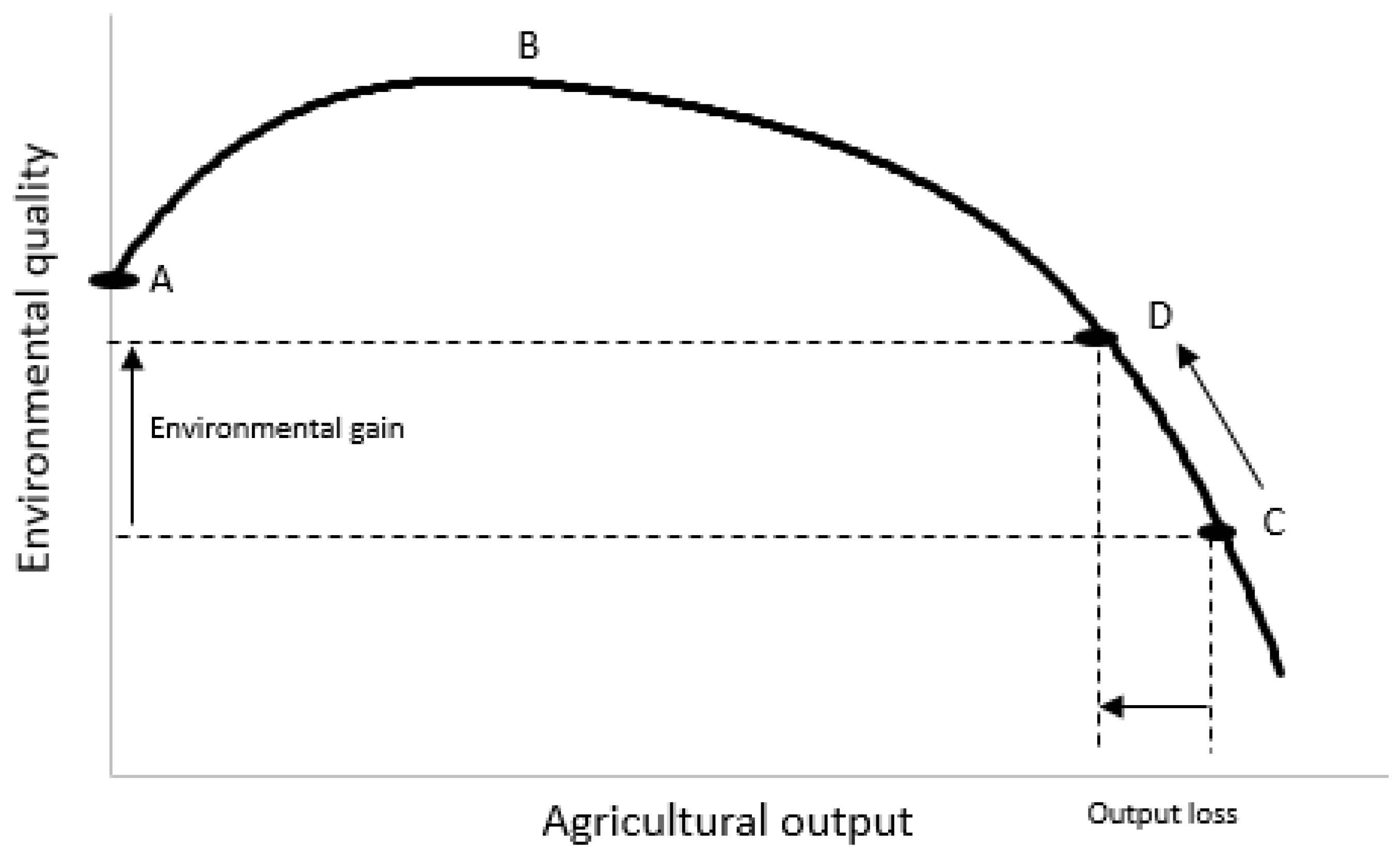

| Species | Feed System | Emissions (Million Tonnes CO2-eq) | Emission Intensity (Kg CO2-eq/kg Product) | ||
|---|---|---|---|---|---|
| Milk | Meat | Milk | Meat | ||
| Dairy | Grazing 1 | 227 | 104 | 2.9 | 21.9 |
| Mixed 2 | 1104 | 382 | 2.6 | 17.4 | |
| Total | 1331 | 486 | 2.6 | 18.2 | |
| Beef | Grazing 1 | - | 875 | - | 102 |
| Mixed 2 | - | 1463 | - | 56 | |
| Total | - | 2338 | - | 67 | |
| Sheep | Grazing1 | 30 | 76 | 9.8 | 23.8 |
| Mixed 2 | 37 | 115 | 7.5 | 23.2 | |
| Total | 67 | 191 | 8.4 | 23.4 | |
| Goat | Grazing 1 | 18 | 27 | 6.1 | 24.2 |
| Mixed 2 | 44 | 84 | 4.9 | 23.1 | |
| Total | 62 | 111 | 5.2 | 23.3 | |
| Total from grazing systems 1 | 275 | 1082 | 6.3 | 43.0 | |
| Total from mixed rations 2 | 1185 | 2044 | 5.0 | 30.0 | |
| Grand total | 1460 | 3126 | 5.4 | 33.0 | |
| Variable | Land Use | ||
|---|---|---|---|
| Pastoral | Cropping and Horticulture | Native Forest | |
| Total N | +0.85 | +0.45 | −0.39 |
| Total P | +0.70 | +0.24 | −0.32 |
| Visual clarity | −0.45 | −0.24 | +0.30 |
| Escherichia coli presence | +0.80 | (0.17) | −0.34 |
| NZ land area (km3) | 107,672 | 4174 | 65,675 |
| Total NZ land area (%) | 39.6 | 1.5 | 24.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caradus, J.R.; Chapman, D.F.; Rowarth, J.S. Improving Human Diets and Welfare through Using Herbivore-Based Foods: 2. Environmental Consequences and Mitigations. Animals 2024, 14, 1353. https://doi.org/10.3390/ani14091353
Caradus JR, Chapman DF, Rowarth JS. Improving Human Diets and Welfare through Using Herbivore-Based Foods: 2. Environmental Consequences and Mitigations. Animals. 2024; 14(9):1353. https://doi.org/10.3390/ani14091353
Chicago/Turabian StyleCaradus, John R., David F. Chapman, and Jacqueline S. Rowarth. 2024. "Improving Human Diets and Welfare through Using Herbivore-Based Foods: 2. Environmental Consequences and Mitigations" Animals 14, no. 9: 1353. https://doi.org/10.3390/ani14091353
APA StyleCaradus, J. R., Chapman, D. F., & Rowarth, J. S. (2024). Improving Human Diets and Welfare through Using Herbivore-Based Foods: 2. Environmental Consequences and Mitigations. Animals, 14(9), 1353. https://doi.org/10.3390/ani14091353