
Preservative Effects of Curcumin on Semen of Hu Sheep
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Reagent Preparation
2.3. Semen Collection
2.4. Determination of Sperm Motility and Viability
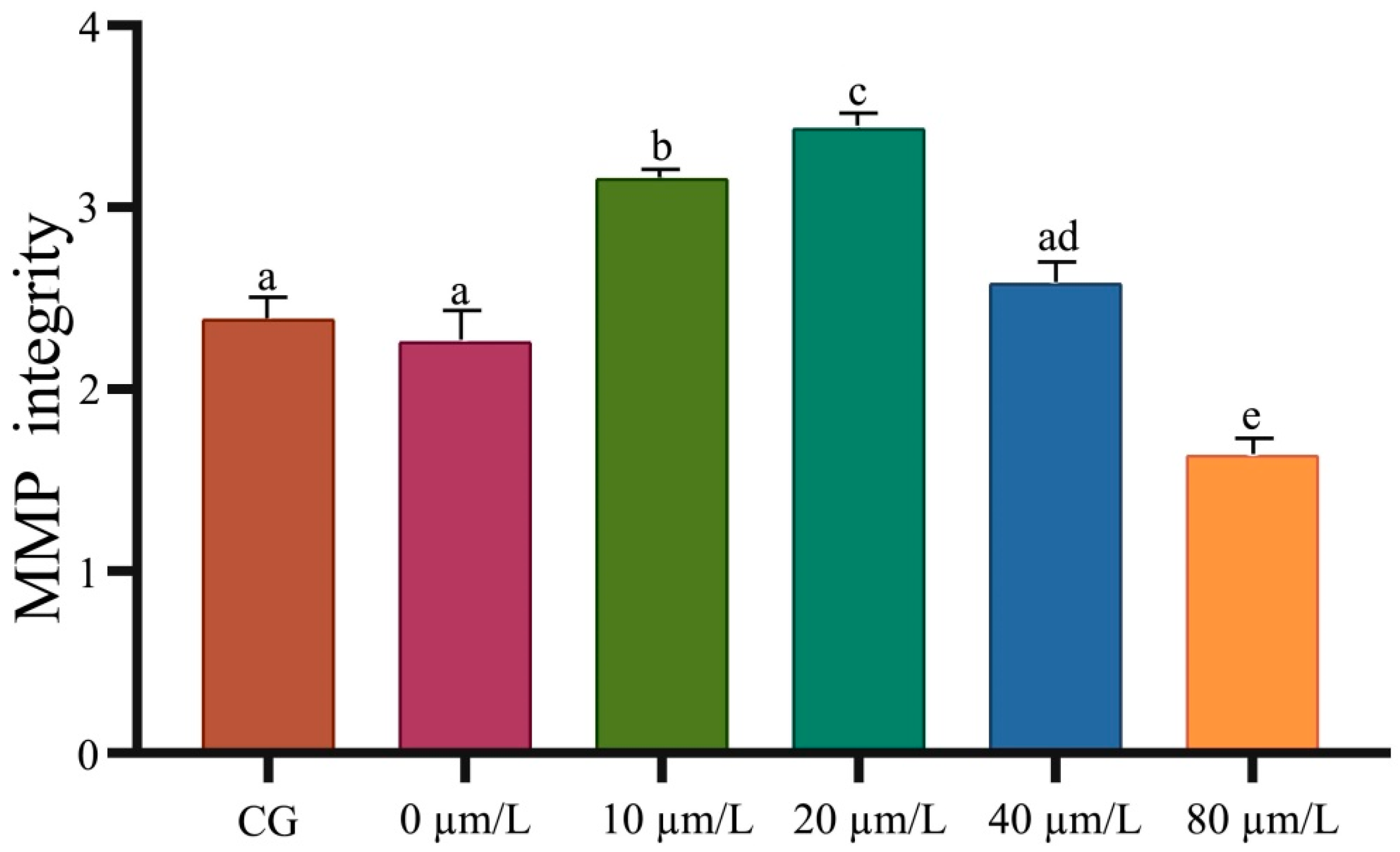
2.5. Determination of Mitochondrial Membrane Potential
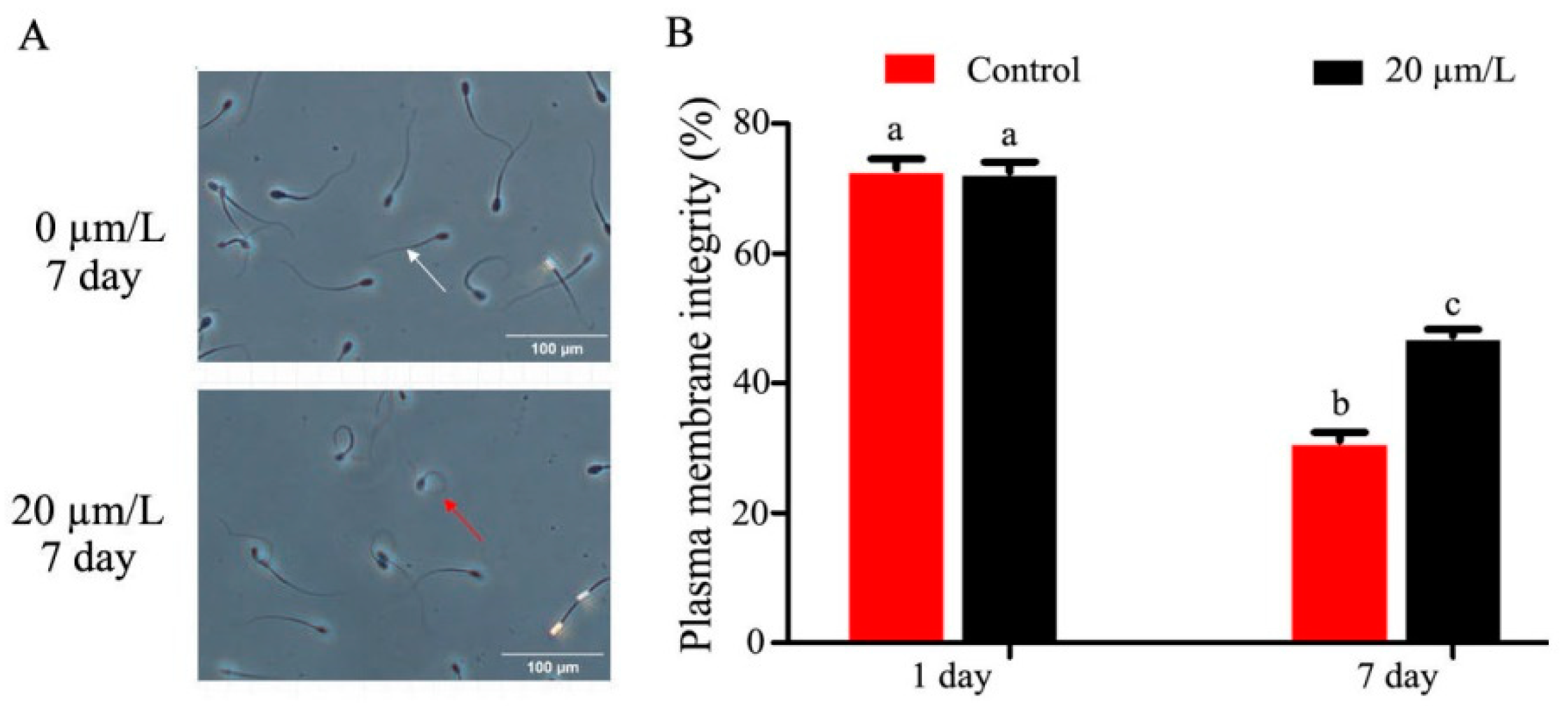
2.6. Determination of Sperm Plasma Membrane Integrity
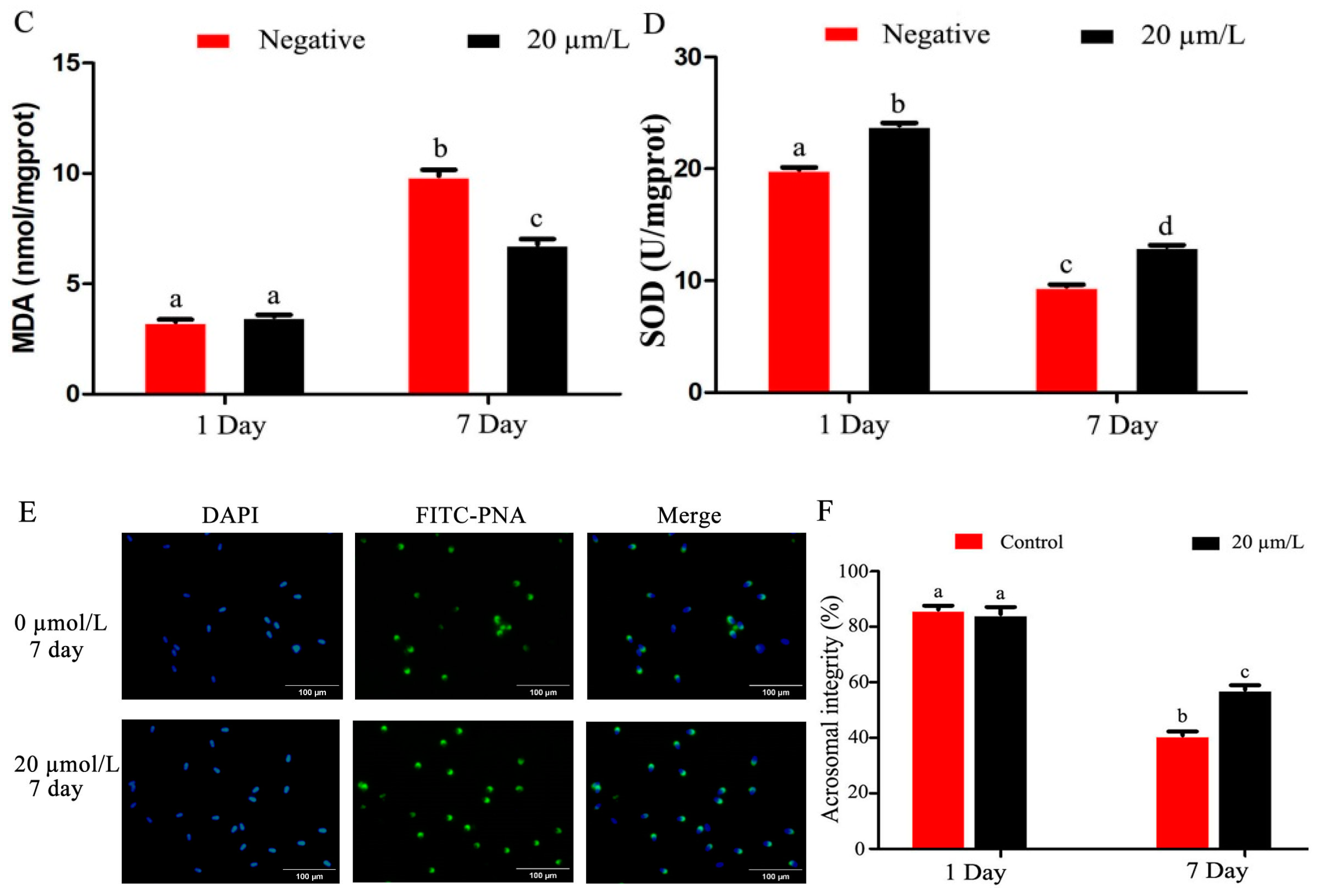
2.7. Determination of Sperm Acrosomal Integrity
2.8. Determination of Sperm Total Antioxidant Capacity
2.9. Determination of Catalase (CAT) Activity
2.10. Determination of Superoxide Dismutase (SOD) Activity
2.11. Determination of Malondialdehyde (MDA) Content
2.12. Determination of ROS
2.13. Purification of Spermatozoa
2.14. Metabolite Extraction
2.15. LC-MS Analysis
2.16. Metabolomics Data Analysis
2.17. Heatmap and KEGG Analysis
2.18. Statistical Analysis
3. Results
3.1. Effects of Curcumin on Sperm Motility
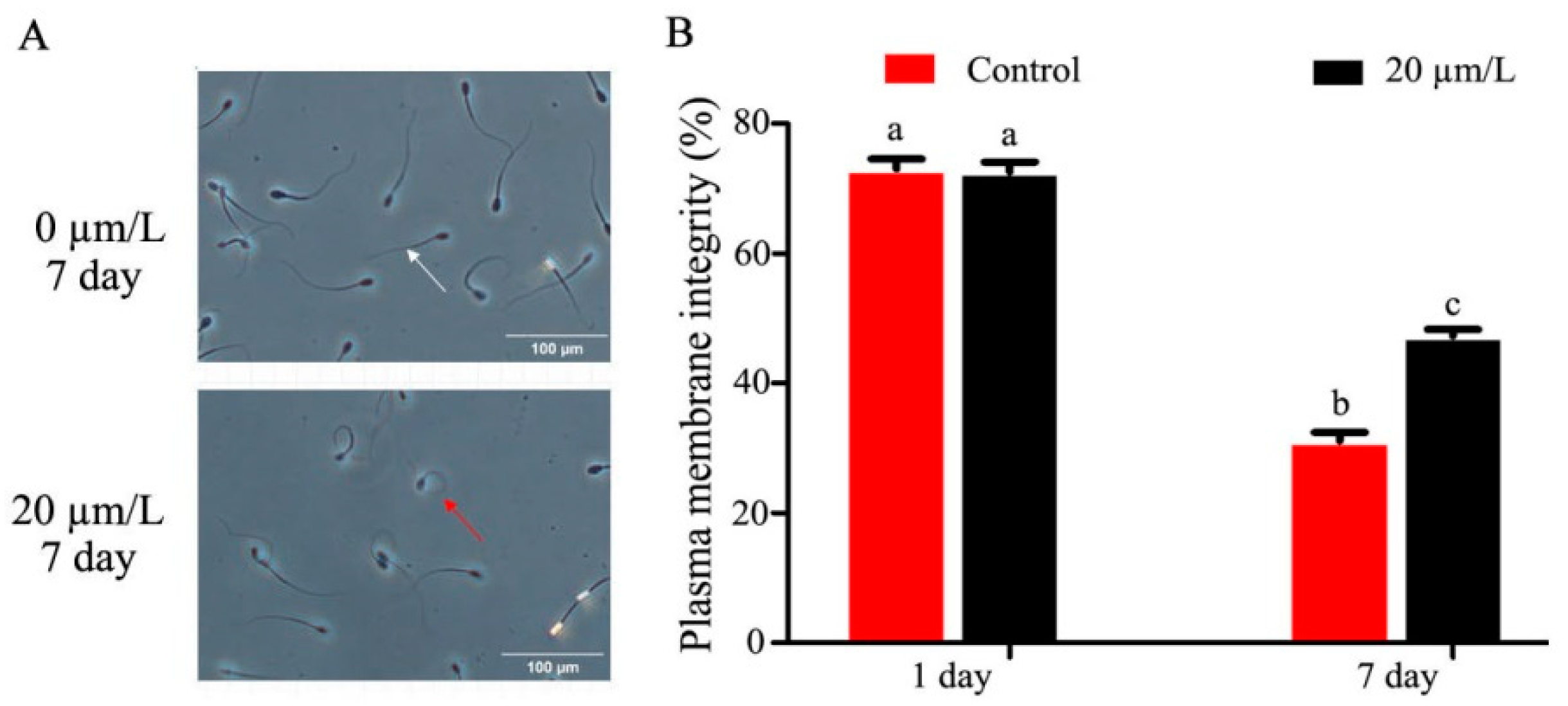
3.2. Effect of Curcumin on Sperm Membrane and Acrosome Integrity
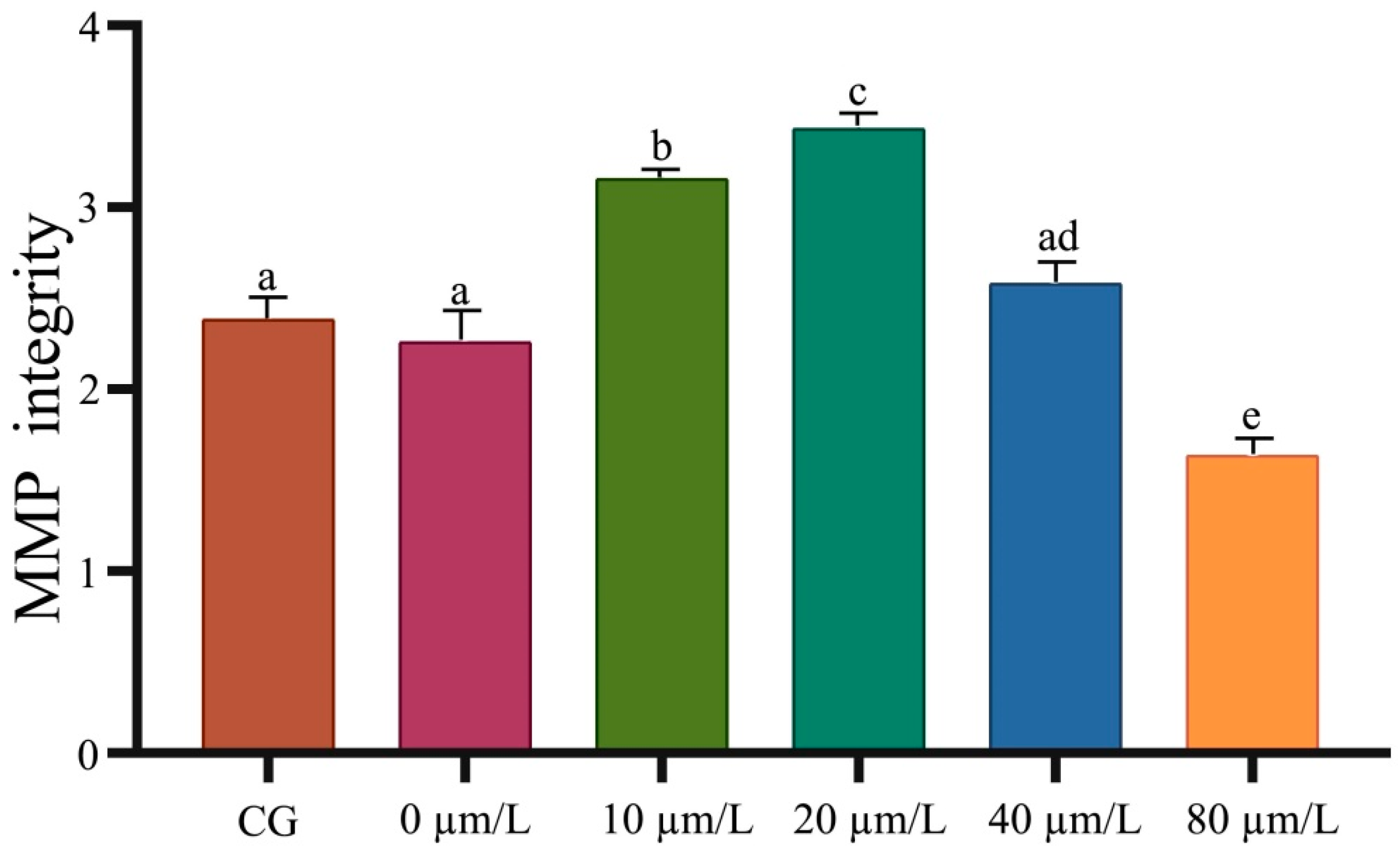
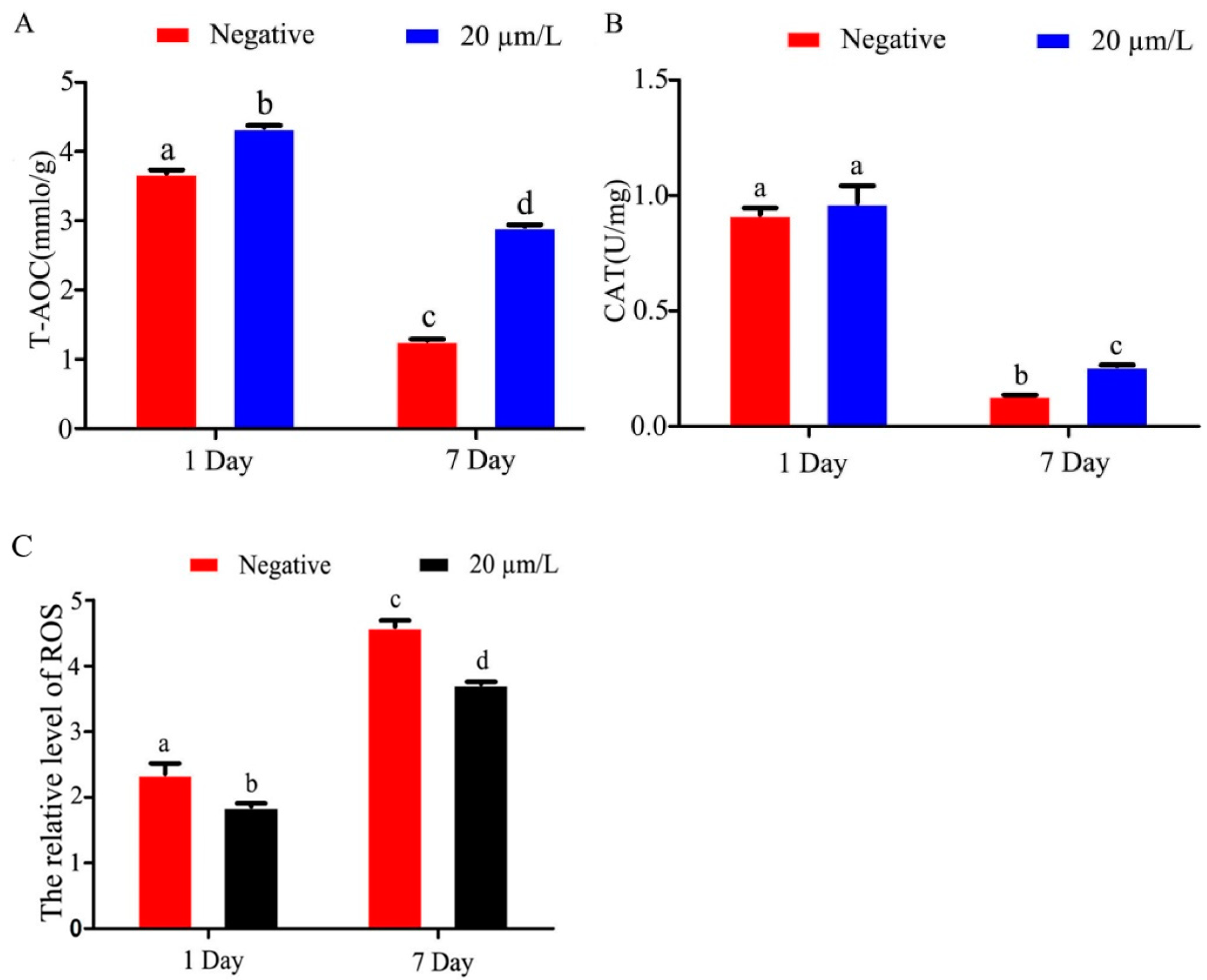
3.3. Effects of Curcumin on Antioxidant Indexes of Sperm
3.4. Metabolite Detection Quality Control
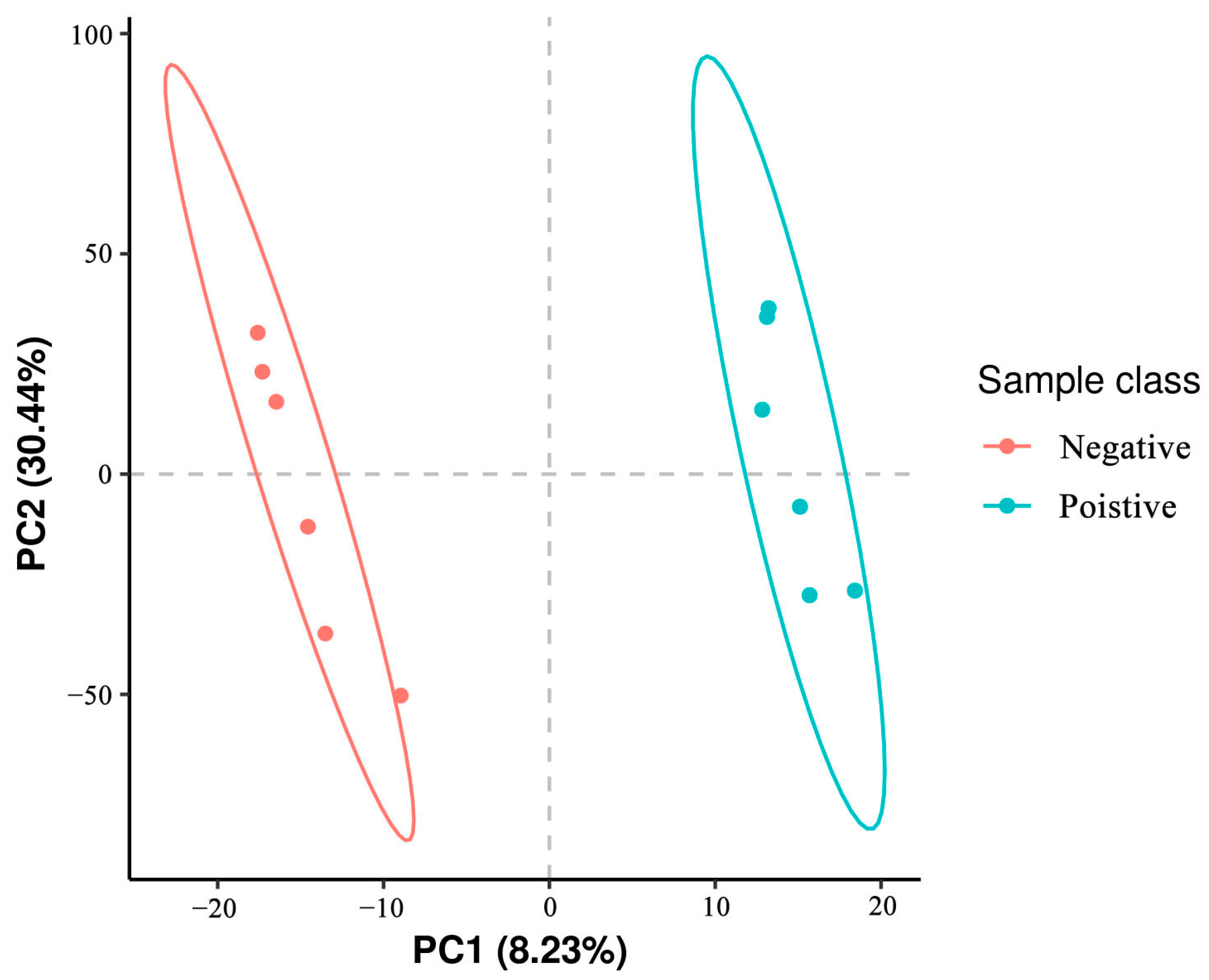
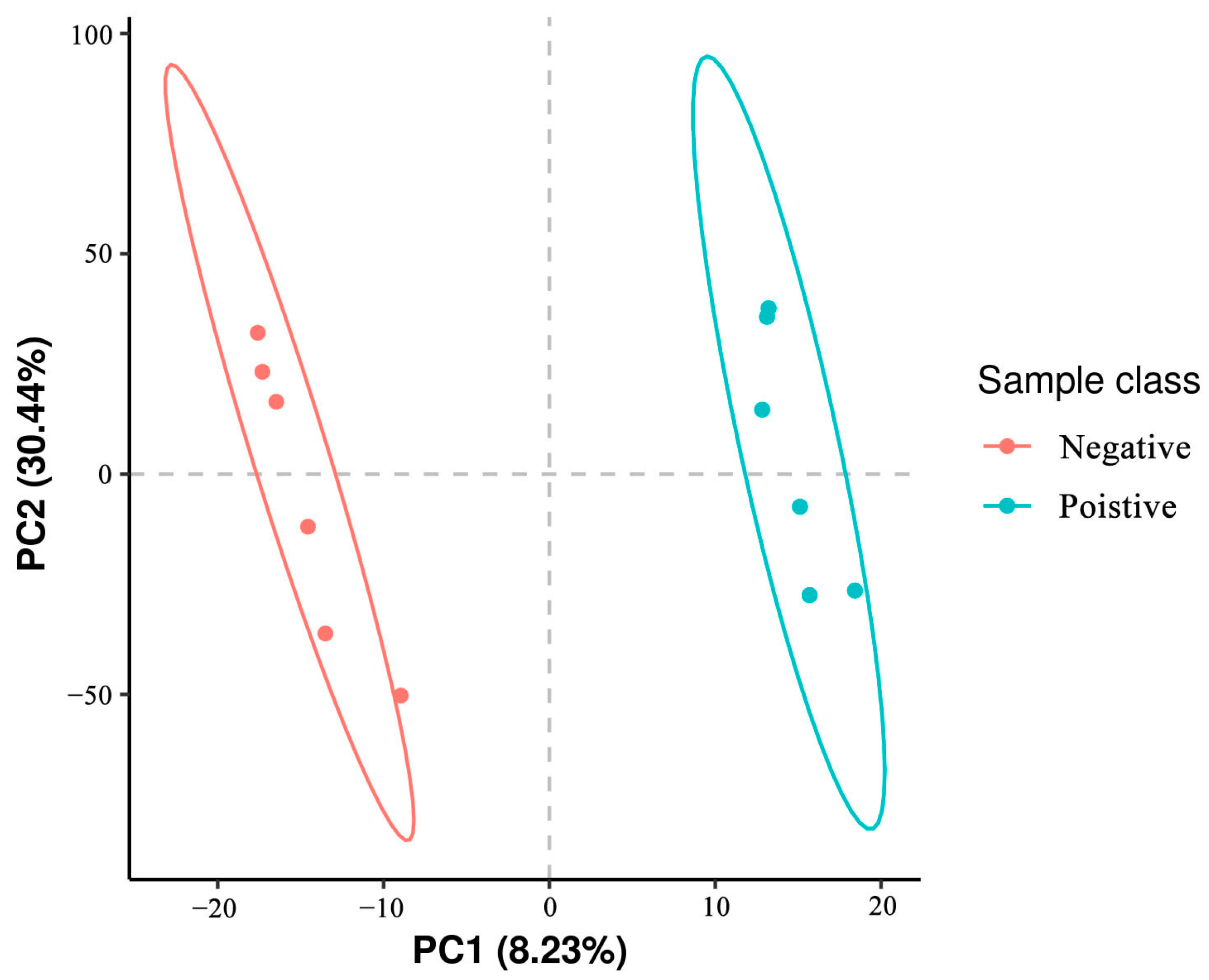
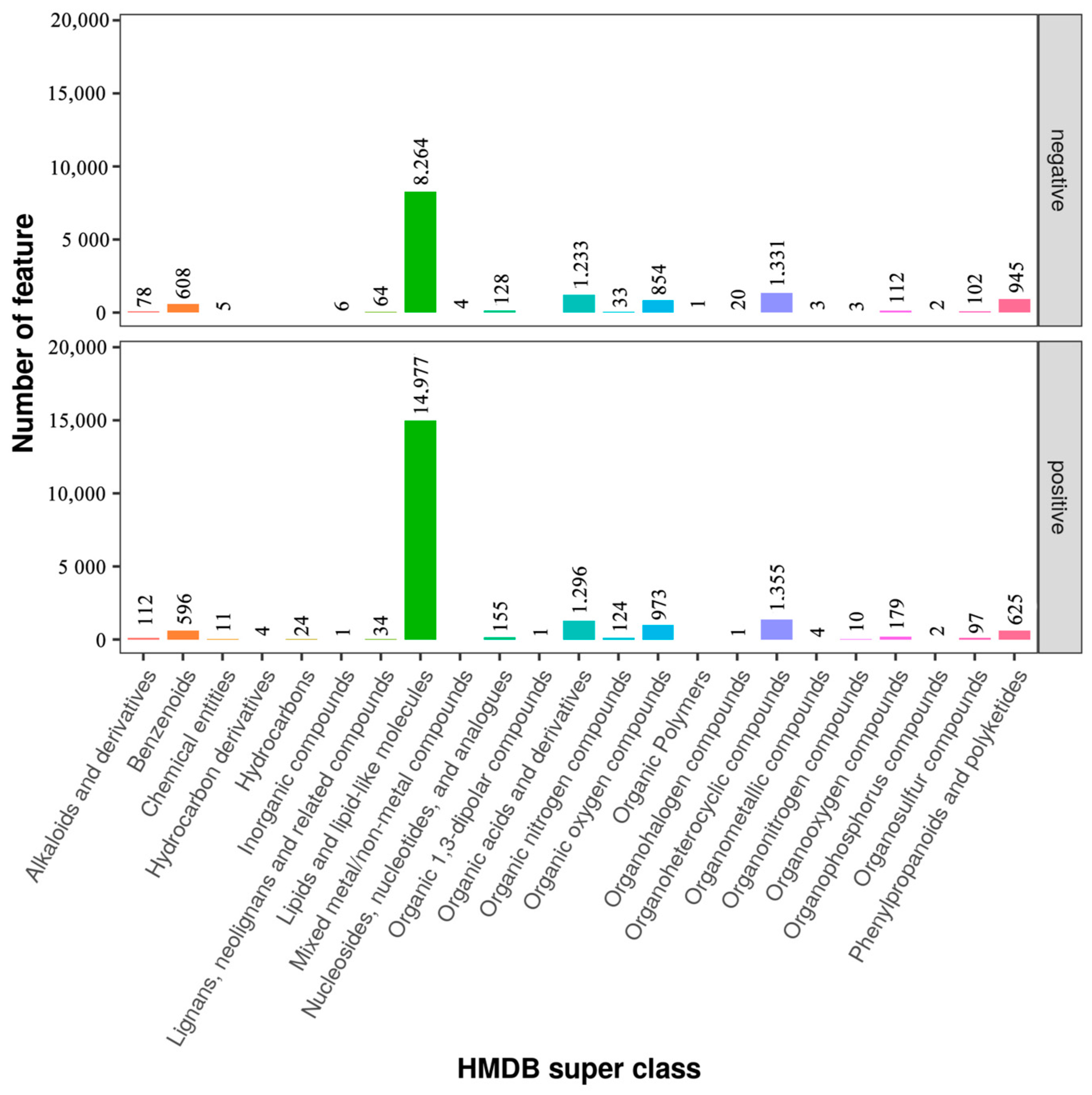
3.5. Classification of Metabolites in Sperm between Negative Groups and Added Curcumin Groups
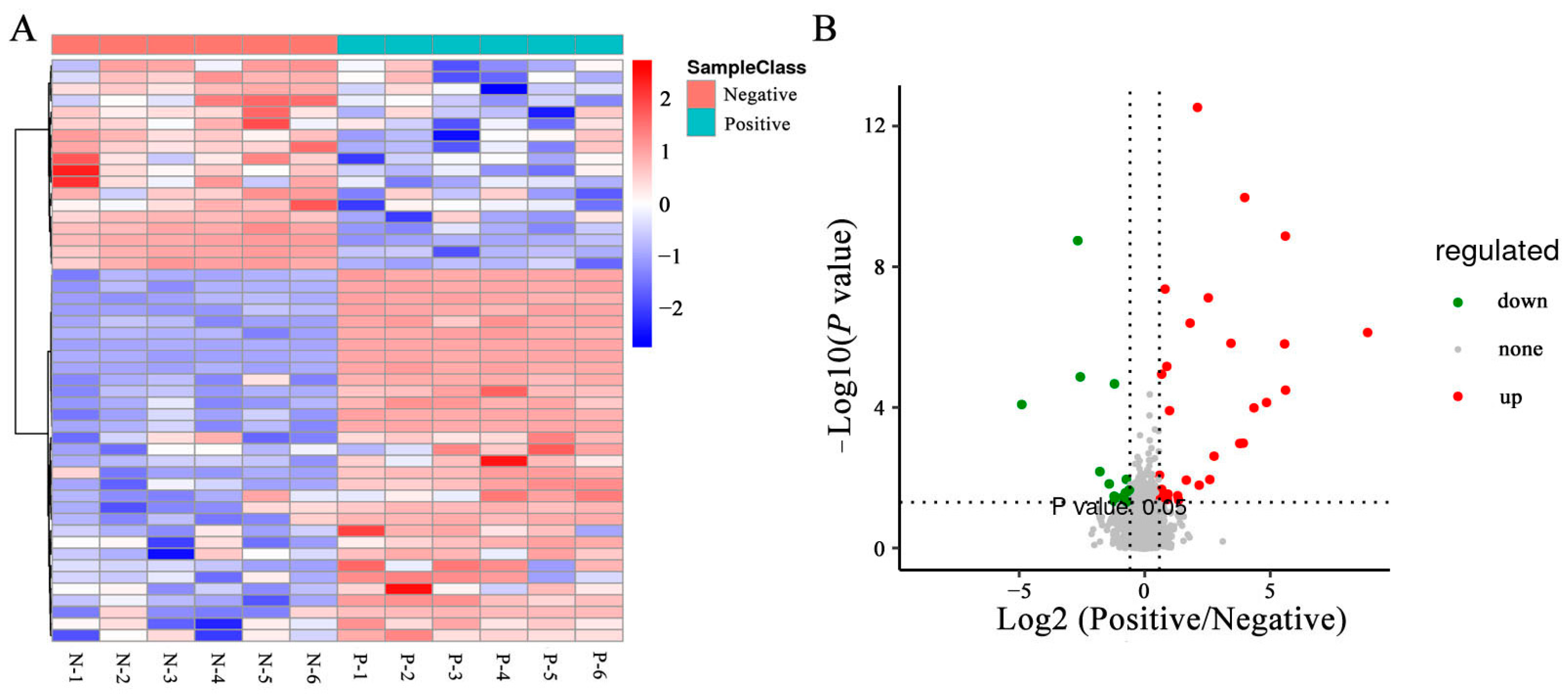
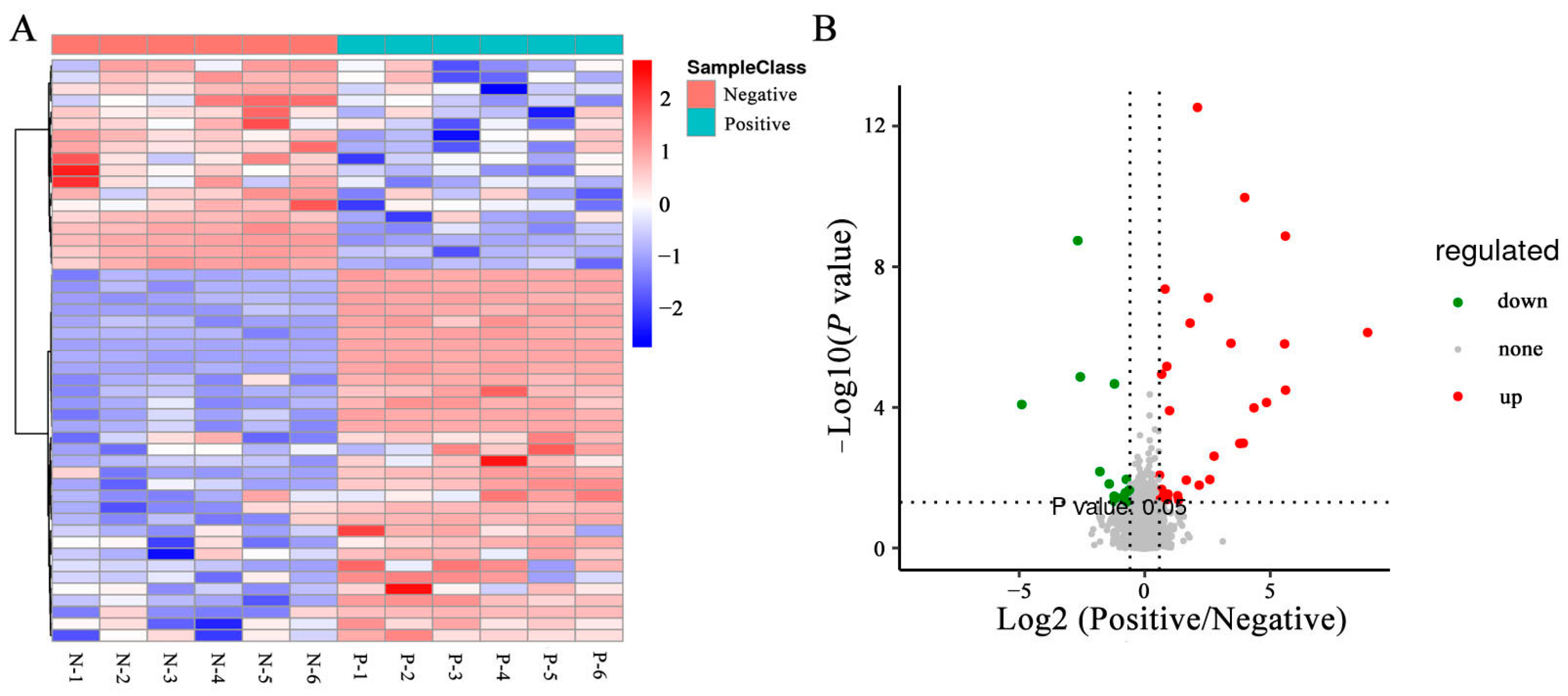
3.6. Effects of Curcumin on Sperm Metabolites in Negative and Added-Curcumin Groups
3.7. Cluster Analyses of DEMs in Sperm between Negative and Added-Curcumin Groups
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barbas, J.P.; Mascarenhas, R.D. Cryopreservation of domestic animal sperm cells. Cell Tissue Bank. 2009, 10, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Tang, Y.L.; Hu, J.L.; Zhou, W.J.; Huang, Z.H.; Luo, X.F.; Li, Z.; Zhu, W.B. Update on techniques for cryopreservation of human spermatozoa. Asian. J. Androl. 2022, 24, 563–569. [Google Scholar]
- Raoul, J.; Elsen, J.M. The levels of artificial insemination and missing sire information make genomic selection not always beneficial in meat sheep. Animal 2021, 15, 100040. [Google Scholar] [CrossRef] [PubMed]
- Turgut, G.; Abban, G.; Turgut, S.; Take, G. Effect of overdose zinc on mouse testis and its relation with sperm count and motility. Biol. Trace Elem. Res. 2003, 96, 271–279. [Google Scholar] [CrossRef]
- Pérez-Llano, B.; López-Fernández, C.; García-Casado, P.; Arroyo, F.; Gosalbez, A.; Sala, R.; Gosálvez, J. Dynamics of sperm DNA fragmentation in the swine: Ejaculate and temperature effects. Anim. Reprod. Sci. 2010, 119, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Mislei, B.; Bucci, D.; Malama, E.; Bollwein, H.; Mari, G. Seasonal changes in ROS concentrations and sperm quality in unfrozen and frozen-thawed stallion semen. Theriogenology 2020, 1, 89–97. [Google Scholar] [CrossRef]
- Martínez-Páramo, S.; Diogo, P.; Dinis, M.T.; Herráez, M.P.; Sarasquete, C.; Cabrita, E. Incorporation of ascorbic acid and α-tocopherol to the extender media to enhance antioxidant system of cryopreserved sea bass sperm. Theriogenology 2012, 77, 1129–1136. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, J.; Pang, W.; Yang, G.; Chu, G. Astaxanthin improves preserved semen quality at 17 °C by enhancing sperm antioxidant and motility properties. Reprod. Domest. Anim. 2022, 57, 1187–1197. [Google Scholar] [CrossRef]
- Lagares, M.A.; da Silva, G.C.; Cortes, S.F.; Moreira, F.H.M.; Neves, F.C.D.; Alves, N.C.; Viegas, R.N.; Diniz, T.F.; Lemos, V.S.; de Rezende, A.S.C.; et al. L-carnitine added to post-thawed semen acts as an antioxidant and a stimulator of equine sperm metabolism. Andrologia 2022, 54, e14338. [Google Scholar] [CrossRef]
- Azawi, O.I.; Hussein, E.K. Effect of vitamins C or E supplementation to Tris diluent on the semen quality of Awassi rams preserved at 5 °C. Vet. Res. Forum 2013, 4, 157–160. [Google Scholar]
- Arkoun, B.; Galas, L.; Dumont, L.; Rives, A.; Saulnier, J.; Delessard, M.; Rondanino, C.; Rives, N. Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue. Int. J. Mol. Sci. 2019, 20, 5380. [Google Scholar] [CrossRef]
- Iqbal, S.; Haleem, S.; Akhtar, M.; Zia-Ul-Haq, M.; Akbar, J. Efficiency of pomegranate peel extracts in stabilization of sunflower oil under accelerated conditions. Food Res. Int. 2008, 41, 194–200. [Google Scholar] [CrossRef]
- Shin, J.W.; Chun, K.S.; Kim, D.H.; Kim, S.J.; Kim, S.H.; Cho, N.C.; Na, H.K.; Surh, Y.J. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochem. Pharmacol. 2020, 173, 113820. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wu, B.; Shen, G.; Li, X.; Wu, Q. Curcumin alleviates liver oxidative stress in type 1 diabetic rats. Mol. Med. Rep. 2018, 17, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.M.; Sheng, M.; Luo, H.Q.; Liu, G.; Meng, F.; Cao, Z.B.; Zhang, Y.H. Characterization of freezability-associated metabolites in boar semen. Theriogenology 2023, 196, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Wen, Z.; Xu, X.; Wang, J.; Li, X.; Meng, K.; Yang, P. Serum Metabolomics Analysis for Biomarkers of Lactobacillus plantarum FRT4 in High-Fat Diet-Induced Obese Mice. Foods 2022, 11, 184. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, M.A.; Yániz, J.L.; Peña, F.J.; Santolaria, P.; Castelló-Ruiz, M. Role of Antioxidants in Cooled Liquid Storage of Mammal Spermatozoa. Antioxidants 2021, 10, 1096. [Google Scholar] [CrossRef] [PubMed]
- Varisli, O.; Uguz, C.; Agca, C.; Agca, Y. Various physical stress factors on rat sperm motility, integrity of acrosome, and plasma membrane. J. Androl. 2009, 30, 75–86. [Google Scholar] [CrossRef]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R.J. Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol. Hum. Reprod. 2015, 21, 502–515. [Google Scholar] [CrossRef]
- Aitken, R.J.; Smith, T.B.; Lord, T.; Kuczera, L.; Koppers, A.J.; Naumovski, N.; Connaughton, H.; Baker, M.A.; De Iuliis, G.N. On methods for the detection of reactive oxygen species generation by human spermatozoa: Analysis of the cellular responses to catechol oestrogen, lipid aldehyde, menadione and arachidonic acid. Andrology 2013, 1, 192–205. [Google Scholar] [CrossRef]
- Zhang, G.; Jiang, F.; Chen, Q.; Yang, H.; Zhou, N.; Sun, L.; Zou, P.; Yang, W.; Cao, J.; Zhou, Z.; et al. Associations of ambient air pollutant exposure with seminal plasma MDA, sperm mtDNA copy number, and mtDNA integrity. Environ. Int. 2020, 136, 105483. [Google Scholar] [CrossRef]
- Aitken, R.J.; Whiting, S.; De Iuliis, G.N.; McClymont, S.; Mitchell, L.A.; Baker, M.A. Electrophilic aldehydes generated by sperm metabolism activate mitochondrial reactive oxygen species generation and apoptosis by targeting succinate dehydrogenase. J. Biol. Chem. 2012, 287, 33048–33060. [Google Scholar] [CrossRef]
- Netherton, J.K.; Hetherington, L.; Ogle, R.A.; Gavgani, M.M.; Velkov, T.; Villaverde, A.I.B.; Tanphaichitr, N.; Baker, M.A. Mass Spectrometry Reveals New Insights into the Production of Superoxide Anions and 4-Hydroxynonenal Adducted Proteins in Human Sperm. Proteomics 2020, 20, e1900205. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive oxygen species impact on sperm DNA and its role in male infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.M.; Wang, T.; Wen, D.G.; Hou, J.Q.; Li, H.B. Protective effect of Rhodiola rosea polysaccharides on cryopreserved boar sperm. Carbohydr. Polym. 2016, 135, 44–47. [Google Scholar] [CrossRef]
- Feng, C.; Zhu, Z.; Bai, W.; Li, R.; Zheng, Y.; Tian, X.; Wu, D.; Lu, H.; Wang, Y.; Zeng, W. Proline Protects Boar Sperm against Oxidative Stress through Proline Dehydrogenase-Mediated Metabolism and the Amine Structure of Pyrrolidine. Animals 2020, 10, 1549. [Google Scholar] [CrossRef]
- Huang, M.C.; Watson, S.R.; Liao, J.J.; Goetzl, E.J. Th17 augmentation in OTII TCR plus T cell-selective type 1 sphingosine 1-phosphate receptor double transgenic mice. J. Immunol. 2007, 178, 6806–6813. [Google Scholar] [CrossRef] [PubMed]
- Eini, F.; Bidadkosh, A.; Nazarian, H.; Piryaei, A.; Ghaffari, N.M.; Joharchi, K. Thymoquinone reduces intracytoplasmic oxidative stress and improves epigenetic modification in polycystic ovary syndrome mice oocytes, during in-vitro maturation. Mol. Reprod. Dev. 2019, 86, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Park, M.T.; Kim, M.J.; Kang, Y.H.; Choi, S.Y.; Lee, J.H.; Choi, J.A.; Kang, C.M.; Cho, C.K.; Kang, S.; Bae, S.; et al. Phytosphingosine in combination with ionizing radiation enhances apoptotic cell death in radiation-resistant cancer cells through ROS-dependent and -independent AIF release. Blood 2005, 105, 1724–1733. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time/(Day) (Day) | Control | 0/(µmol/L) | 10/(µmol/L) | 20/(µmol/L) | 40/(µmol/L) | 80/(µmol/L) |
|---|---|---|---|---|---|---|
| 1 | 97.86 ± 0.71 | 95.59 ± 0.38 | 97.92 ± 0.77 | 96.75 ± 2.31 | 97.87 ± 1.72 | 96.03 ± 1.00 |
| 2 | 92.68 ± 2.18 ab | 94.27 ± 0.71 a | 94.03 ± 2.58 a | 94.71 ± 2.72 a | 93.76 ± 2.61 ab | 90.21 ± 1.69 b |
| 3 | 92.11 ± 1.52 a | 93.93 ± 0.81 a | 94.19 ± 1.18 a | 93.38 ± 0.87 a | 92.95 ± 1.61 a | 87.88 ± 1.59 b |
| 4 | 80.09 ± 3.97 cd | 82.86 ± 0.70 bc | 85.99 ± 1.73 ab | 87.81 ± 1.28 a | 77.51 ± 3.05 de | 75.49 ± 2.73 e |
| 5 | 76.52 ± 1.72 b | 78.95 ± 0.92 b | 82.31 ± 1.44 a | 83.29 ± 1.34 a | 76.58 ± 1.47 b | 69.34 ± 1.48 c |
| 6 | 52.23 ± 1.58 d | 51.21 ± 2.16 d | 62.03 ± 1.84 b | 66.48 ± 2.07 a | 56.42 ± 1.35 c | 52.24 ± 1.59 d |
| 7 | 42.80 ± 3.26 d | 43.82 ± 3.70 d | 52.02 ± 3.21 b | 60.94 ± 2.03 a | 48.17 ± 2.61 bc | 45.15 ± 2.85 c |
| Time/(Day) (Day) | Control | 0/(µmol/L) | 10/(µmol/L) | 20/(µmol/L) | 40/(µmol/L) | 80/(µmol/L) |
|---|---|---|---|---|---|---|
| 1 | 94.00 ± 0.97 | 96.17 ± 1.06 | 94.62 ± 1.13 | 93.66 ± 2.91 | 93.69 ± 3.95 | 93.40 ± 1.17 |
| 2 | 84.54 ± 1.27 b | 88.74 ± 1.90 a | 89.31 ± 1.92 a | 89.37 ± 1.84 a | 85.75 ± 0.66 b | 83.24 ± 2.58 b |
| 3 | 80.54 ± 2.29 c | 84.83 ± 0.93 b | 84.36 ± 1.10 b | 86.71 ± 0.81 a | 81.27 ± 2.53 c | 81.48 ± 2.11 c |
| 4 | 67.71 ± 1.70 c | 72.54 ± 1.20 b | 74.87 ± 1.86 b | 79.27 ± 1.87 a | 67.05 ± 3.36 cd | 64.15 ± 1.85 d |
| 5 | 54.81 ± 2.60 c | 56.61 ± 2.66 c | 60.21 ± 1.26 b | 73.70 ± 2.22 a | 63.23 ± 2.17 b | 56.59 ± 2.06 c |
| 6 | 35.55 ± 2.09 c | 43.53 ± 1.02 c | 54.31 ± 1.46 b | 60.84 ± 2.30 a | 45.94 ± 3.95 c | 45.29 ± 2.82 c |
| 7 | 34.27 ± 2.93 c | 40.26 ± 2.79 b | 42.21 ± 2.00 b | 53.27 ± 1.71 a | 35.68 ± 3.69 c | 35.26 ± 2.64 c |
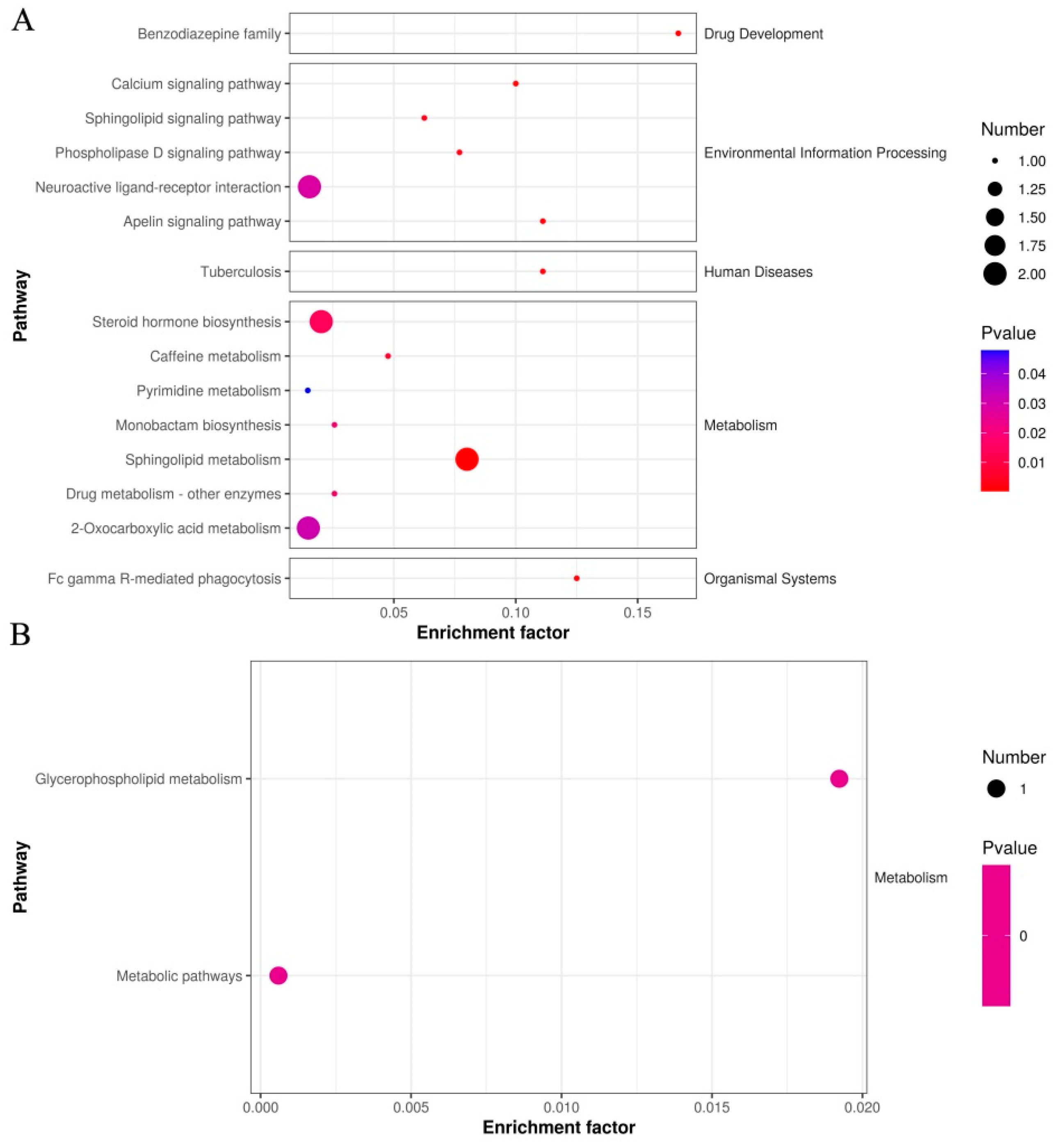
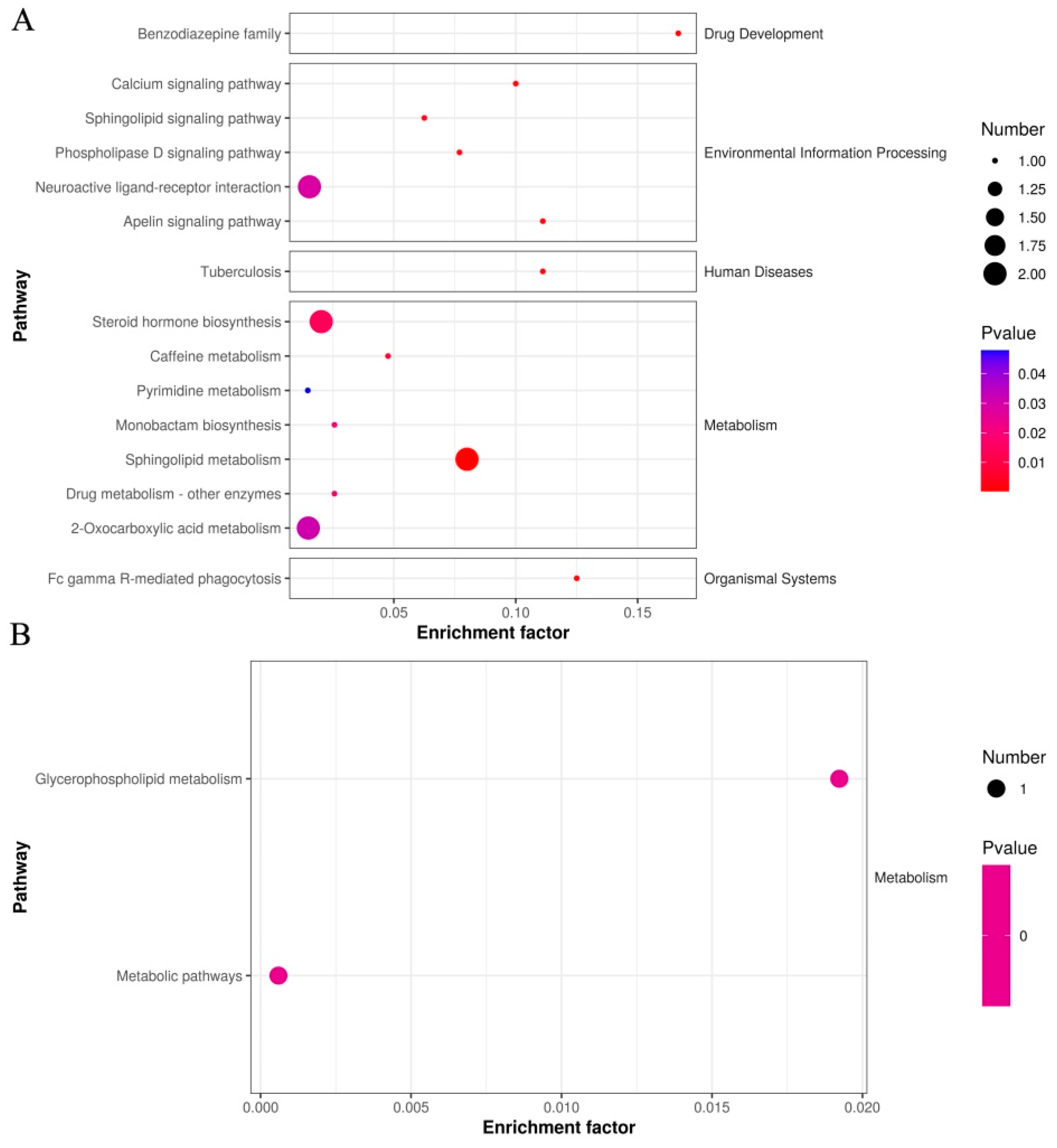
| Metabolic Pathway | p Value | FDR | Compound | Compound ID |
|---|---|---|---|---|
| Benzodiazepine family | 0.0003 | 0.0032 | L-365260 | C15026 |
| Apelin signaling pathway | 0.0009 | 0.0033 | Sphingosine 1-phosphate | C06124 |
| Calcium signaling pathway | 0.0011 | 0.0034 | Sphingosine 1-phosphate | C06124 |
| Phospholipase D signaling pathway | 0.0019 | 0.005 | Sphingosine 1-phosphate | C06124 |
| Sphingolipid signaling pathway | 0.0029 | 0.0067 | Sphingosine 1-phosphate | C06124 |
| Neuroactive ligand-receptor interaction | 0.0284 | 0.0394 | UTP; Sphingosine 1-phosphate | C00075; C06124 |
| Tuberculosis | 0.0009 | 0.0033 | Sphingosine 1-phosphate | C06124 |
| Caffeine metabolism | 0.0051 | 0.0102 | Xanthosine | C01762 |
| Monobactam biosynthesis | 0.0170 | 0.0256 | MM 42842 | C20928 |
| Metabolic pathways | 0.8469 | 0.847 | UTP; Xanthosine; Gibberellin A20; Sphingosine 1-phosphate; Cephaeline; Phytosphingosine | C00075; C01762; C02035; C06124; C09390; C12144 |
| 2-Oxocarboxylic acid metabolism | 0.0307 | 0.0395 | (R)-(Homo)2-citrate; (-)-threo-Iso(homo)2-citrate | C16583; C16597 |
| Sphingolipid metabolism | 0.0002 | 0.0033 | Sphingosine 1-phosphate; Phytosphingosine | C06124; C12144 |
| Steroid hormone biosynthesis | 0.0138 | 0.0249 | Dehydroepiandrosterone sulfate; 2-Methoxyestrone 3-glucuronide | C04555; C11132 |
| Purine metabolism | 0.0817 | 0.092 | Xanthosine | C01762 |
| Pyrimidine metabolism | 0.0479 | 0.0575 | UTP | C00075 |
| Drug metabolism–other enzymes | 0.0170 | 0.0256 | 5-Fluorouridine | C16633 |
| Bile secretion | 0.0911 | 0.0965 | Dehydroepiandrosterone sulfate | C04555 |
| Fc gamma R-mediated phagocytosis | 0.0007 | 0.0033 | Sphingosine 1-phosphate | C06124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, K.; Wei, J.; Fan, Z.; Zhu, M.; Yuan, X.; Zhang, S.; Li, S.; Xu, H.; Ling, Y. Preservative Effects of Curcumin on Semen of Hu Sheep. Animals 2024, 14, 947. https://doi.org/10.3390/ani14060947
Ji K, Wei J, Fan Z, Zhu M, Yuan X, Zhang S, Li S, Xu H, Ling Y. Preservative Effects of Curcumin on Semen of Hu Sheep. Animals. 2024; 14(6):947. https://doi.org/10.3390/ani14060947
Chicago/Turabian StyleJi, Kaiyuan, Jinbo Wei, Zhiwei Fan, Mengkang Zhu, Xin Yuan, Sihuan Zhang, Shuang Li, Han Xu, and Yinghui Ling. 2024. "Preservative Effects of Curcumin on Semen of Hu Sheep" Animals 14, no. 6: 947. https://doi.org/10.3390/ani14060947
APA StyleJi, K., Wei, J., Fan, Z., Zhu, M., Yuan, X., Zhang, S., Li, S., Xu, H., & Ling, Y. (2024). Preservative Effects of Curcumin on Semen of Hu Sheep. Animals, 14(6), 947. https://doi.org/10.3390/ani14060947