1. Introduction
Coffee is one of the most consumed beverages in the world and the second most important commodity produced by developing countries (after petroleum) [
1]. In 2019/2020, about 10 million tons of bagged coffee were consumed worldwide [
2], generating a large amount of by-product (pulp, husks, silverskin, and coffee waste) in this industry. In 2022, Italy was the third-largest importer of green coffee in the world (after the USA and Germany) and the second-largest exporter of roasted coffee in the EU (after Germany). Coffee is imported into Italy for roasting, which is the industrial process that determines the final taste and aroma of the cup of coffee. Coffee silverskin (CB), the tegument of the outer layer of the coffee bean, is the only by-product of the roasting process. Since roasting 10 tons of coffee produces about 83 kg of CB [
3], this by-product is a potential pollutant if simply discarded into the environment [
4]. Coffee silverskin is high in fiber (62%), protein (19%), fat (from 1.6 to 3.3%), and minerals (5% ash). In addition, this by-product has an antioxidant effect due to the presence of melanoidins and phenolic compounds [
3,
5] (e.g., chlorogenic acid). Various utilization alternatives have been explored to both reduce the environmental impact and increase the added value of CB, such as the recovery of functional ingredients for potential applications in the food, pharmaceutical, and cosmetics industries, direct energy production, composting, bioenergy, and the production of biomaterials [
3,
6,
7,
8,
9,
10]. The conversion of CB by larvae of the black soldier fly (BSF),
Hermetia illucens (Diptera: Stratiomydae), represents an interesting recycling alternative that complies with the principles of sustainability and the circular economy. Indeed, BSF larvae are able to convert large amounts of organic by-products into valuable nutrients (proteins and lipids) that could be used as ingredients for animal feed [
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23,
24,
25] and renewable energy [
26,
27,
28]. In addition, their frass (larval excreta mixed with substrate residues) can be used as an effective organic fertilizer to develop more sustainable agriculture [
29].
In 2017, the European Commission authorized the use of insects as an ingredient in aquafeed (Reg. (EU) 2017/893), including BSF. In addition, in 2021, the EU authorized the use of insect proteins in poultry and pig feed (Reg. (EU) 2021/1372). BSF larvae are rich in nutrients such as fat, protein, and high-quality amino acids and minerals, making them a good source of protein. Feed enriched with BSF larvae appears to improve growth performance and digestibility in pigs and poultry compared to other protein feeds. At the same time, BSFL larvae are also rich in minerals and chitin and have antioxidant and positive effects on the immune system [
30].
The composition of BSF larvae guarantees a supply of proteins and essential amino acids very similar to that of conventional aquafeed protein sources [
31,
32], but although they have a high fat content, the proportion of polyunsaturated fatty acids is very low [
17,
23,
33,
34,
35,
36,
37,
38]. However, the nutritional composition of BSF larvae, especially the lipid content and fatty acid profile, can be modulated by the feed substrate according to the nutritional requirements of the fish [
23,
39,
40,
41,
42,
43]. In addition, the quality of feed substrate also influences the performance of BSF larvae and their bioconversion efficiency [
12,
20,
24,
44,
45,
46,
47].
To the best of the authors’ knowledge, few studies have been conducted on the environmental impact of BSF larvae production [
16,
19,
48,
49,
50]. Nowadays, the most digestible and nutritious ingredients for aquafeed are fishmeal and fish oil (FAO, 2020). However, these ingredients are no longer sustainable, and alternatives are needed to promote sustainability while maintaining fish welfare [
51]. Feed is the most important production cost in aquaculture [
52]. Researchers have examined the environmental sustainability of various alternative ingredients using Life Cycle Assessment (LCA) and reported that insect-derived protein and lipid sources are the most environmentally friendly solutions compared to other alternatives such as microalgae [
50]. It should be noted that LCA analyses take into account the most important impact categories, such as global warming, resource depletion, acidification, and eutrophication, but neglect some important aspects, such as the decline of wild fish. With the help of LCA, it is possible to understand whether the proposed alternative useful for replacing fishmeal and fish oil can have a significant impact on other very important issues; otherwise, there is a risk that one problem is solved but many others are created.
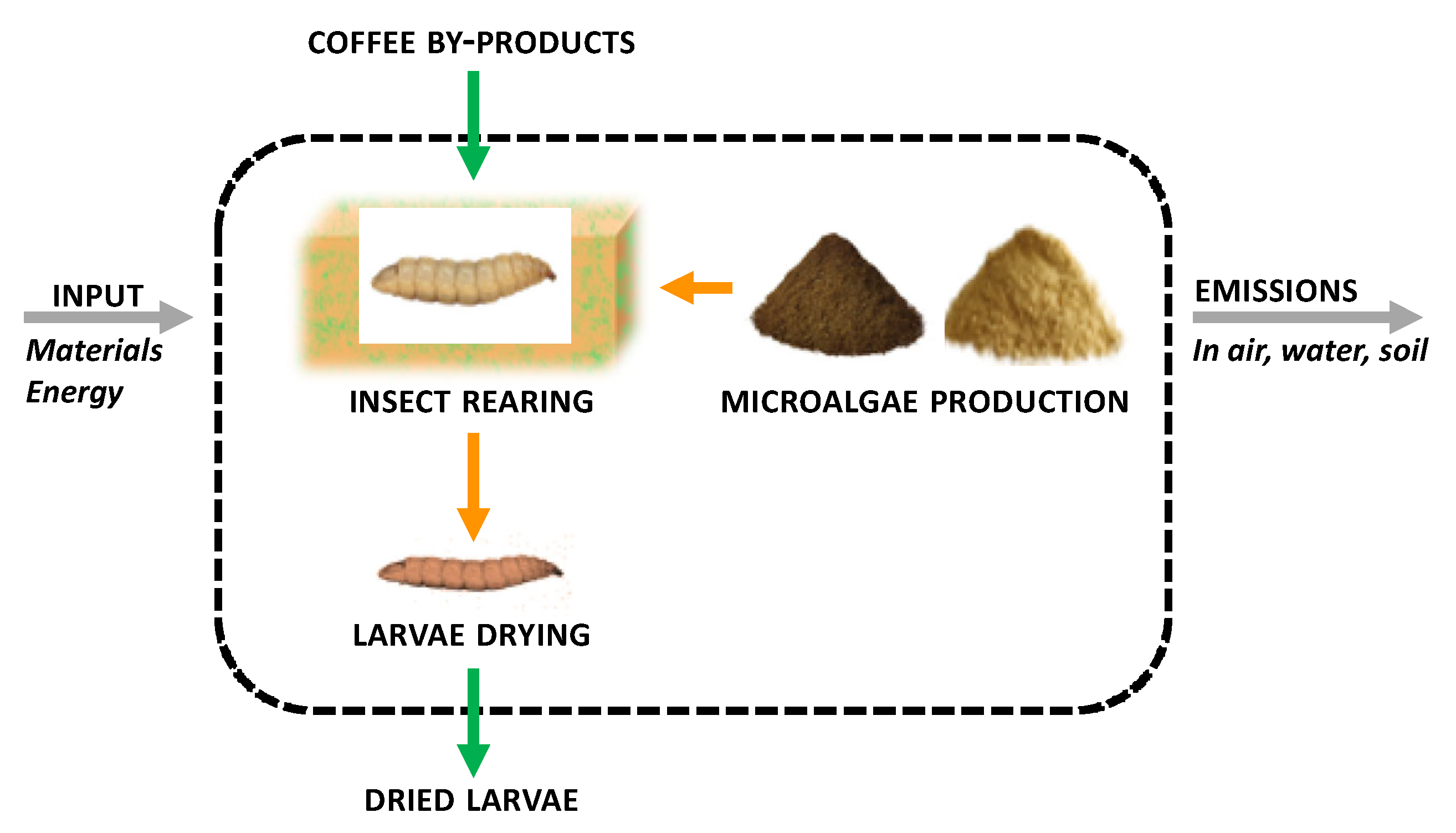
In this context, as part of the project “New nutrients for the production of valuable fish species—Nutrifish”, the locally available by-product of coffee roasting (coffee silverskin) was used as a basal rearing substrate for BSF larvae. This basic substrate for rearing was enriched with an increasing amount of biomass of
Schizochytrium limacinum or
Isochrysis galbana microalgae as a source of valuable vitamins, proteins, and polyunsaturated fatty acids [
53,
54,
55,
56,
57]. In particular, the present study was to investigate: (i) larval growth performance; (ii) final total insect biomass; (iii) macronutrient composition of mature larvae (prepupae); (iv) bioconversion efficiency; and (v) assessment of the environmental sustainability of insect biomass production by LCA.
4. Discussion
Several studies have been conducted on the nutritional requirements of BSF [
12,
14,
15,
40] and on the evaluation of by-products used as feed [
15,
17,
67,
68], but much more information is needed. For this reason, the growth performance of BSF and its impact on the environment were investigated when reared on potentially environmentally acceptable feed substrates,
S. limacinum or
I. galbana-enriched coffee silverskin. The use of a residual substrate has, in general terms, the potential to improve the sustainability of a product by substituting a dedicated input, but this sustainability has to be proven with studies and results. For this reason, we carried out an LCA supporting the insect production results. This study provided data on the effects of enriching the by-product coffee silverskin with different microalgae inclusions (
S. limacinum and
I. galbana) on the growth performance of BSF. These rearing substrates were characterized by a different chemical composition, which influenced the growth performance of the larvae. Considering that BSF stores most of their nutrients during larval development, as the adult stage is unable to feed [
69], it is clear that the quality of the rearing substrate is crucial for their fitness [
45,
70,
71]. Therefore, the nutrient composition of the rearing substrates had a major impact on critical production factors such as survival rate and larval weight, which are positively correlated with growth rate [
12,
67,
72,
73]. As nutrition is a crucial factor in insect fitness and environmental impact, studying its effects on insects, processing conditions, and final product characteristics is very important [
16]. In terms of producing more sustainable ingredients for aquafeed, it is of great interest to influence the nutritional value of insects by enriching the rearing substrate. Cb in particular consists mainly of fibers and carbohydrates, which makes this by-product unsuitable for the formulation of fish feed [
74]. Fish feed must generally have a high protein and lipid content, followed by very low amounts of fiber and carbohydrates, which are mainly indigestible for fish [
75]. Furthermore, the use of microalgae as a component of aquafeed has been extensively studied because of their beneficial properties, but they may only be used in small quantities for economic reasons and because they can lead to an impairment of nutrient absorption in the fish gut [
76]. On the contrary, various studies have shown that the use of insects grown on a substrate enriched with microalgae can positively transfer important bioactive molecules to the fish feed and thus have a positive effect on the overall health of the fish [
77,
78].
In this study, in agreement with Truzzi et al. [
57], both microalgae species showed a higher crude protein (average of 16.40 g/100 g dry matter for both microalgae) and lipid content (average of 6.15 g/100 g dry matter for
S. limacinum and an average of 4.26 g/100 g dry matter for
I. galbana) than the coffee silverskin (protein 13.47 g/100 g dry matter, lipids 3.18 g/100 g dry matter). Consequently, the best growth performance was achieved with the higher microalgae contents. Although both experimental diets significantly increased larval weight and survival,
S. limacinum provided a greater benefit than
I. galbana. As the composition of the two microalgae was different,
S. limacinum provided a higher amount of proteins and lipids compared to
I. galbana, with an emphasis on unsaturated fatty acids.
A shorter larval development time was observed in BSF larvae reared on microalgae-enriched diets compared to the control diet (19 days on average for both microalgae versus 39 days for CB). The development time was also shorter compared to the reports of other authors [
12,
45,
46,
79]. The development time of larvae reared with higher microalgal inclusions (25S and 25I) was 18 days, similar to Barragan-Fonseca et al. [
80]. The development time of larvae reared only with coffee silverskin was very long, showing that rearing with low-fat substrates tends to negatively affect larval fitness. This result confirms what is reported in the current literature, namely that BSF larvae require a high-fat diet to store the body fat needed for development [
69,
70,
81]. If the rearing substrate has a low lipid content, the larvae take longer to acquire their body fat and complete their development [
45].
Substrates enriched with microalgae proved to be more effective compared to coffee silverskin, also in terms of increasing prepupal weight (average 164.6 mg for
S. limacinum, average 123.9 mg for
I. galbana, and average 72.4 mg for CB). Indeed, the weight of larvae reared on the experimental diets (0.1363 g on average) was significantly higher than that of larvae reared on the control diet (0.0724 g), and they were comparable to the values reported in the literature by different authors [
45,
70,
81,
82,
83]. In agreement with Nguyen et al. [
45,
81], we observed that larval weight increased with both the protein and fat content of the diet.
Faster larval development indicates a higher growth rate in relation to the final biomass obtained in a shorter time, which is an advantage for rearing [
84]. In fact, the growth rate was also better for larvae reared on microalgae-enriched feed. In particular, the highest growth rate was recorded for the diet enriched with
S. limacinum, followed by the diet enriched with
I. galbana, while the larvae reared only with coffee silverskin showed the lowest growth rate. These results confirm that although BSF require proteins and lipids in their diet, an excessive amount of these macromolecules in the substrate (e.g., lipids in an
I. galbana-enriched diet) is often detrimental to the growth rate, as the larvae may have problems metabolizing too much fat during the metamorphosis process [
45,
85]. The survival rate obtained in the present study for all experimental and control diets was in line with the range reported by several authors for a variety of rearing substrates [
12,
19,
79,
81,
83].
Nowadays, by-products are becoming increasingly important feed ingredients [
86], especially as they are considered less economically valuable and less polluting than the main product [
87]. Their advantage also depends on how well these by-products can be converted into body mass by the insects [
12]. For this reason, the combination of different substances, including by-products, can be used as an efficient feed, and their composition is the most important variable to determine the FCR [
88]. Basically, FCR is the amount of feed needed to achieve a weight gain of one kilogram for the farm animal. In order to achieve better FCRs that provide optimal economic and environmental benefits when using BSF as an alternative feed, it is necessary that the tested feeds are directly and effectively utilized by the insects. In this work, FCR was more favorable for microalgae-enriched feeds, indicating better utilization of feed nutrients. The FCR values obtained in this work (from 4.5 to 9.5 for microalgae-enriched feeds; 34.2 for control feeds) were much higher than the FCR values obtained by Oonicx et al. [
12] (from 1.4 to 2.6), who studied by-products from food production. These values were lower than those obtained by Rehman et al. [
89] (from 6.3 to 10.1), who studied larvae reared on dairy fertilizer and soybean residues. The results of this work show that high-protein diets lead to lower FCRs, as also reported by Oonincx et al. [
12]. Indeed, the composition and density of proteins are important for insects, as they do not require energy to maintain a constant body temperature [
90,
91].
Finally, the waste reduction index was also analyzed, a parameter used to calculate the insect’s ability to reduce waste and the conversion efficiency of food into biomass. The higher the WRI value, the more effective the larvae are at converting by-products [
19]. In this work, the WRI value of 10S (2.6%/day) was found to be higher than the other diets in the experiment (less than 2.2%/day). This WRI value observed in this work agrees with that of Leong et al. [
92], who reared BSF larvae on fruit waste from a cafeteria (2.8%/day) and palm decanter cake (2.8%/day), but much less than Bava et al. [
19], who reared larvae on okara (4.9%/day), corn distillers (3.2%/day), chicken feed, and brewers grain feed (3.0%/day).
The addition of microalgal biomass to the rearing substrate led to a significant modulation of the nutrient composition of the BSF larvae. The proximate composition of BSF larvae reared on diets containing increasing levels of microalgae resulted in a significant increase in both protein and lipid content compared to larvae reared on coffee silverskin alone, confirming the hypothesis that modulation of substrate composition also affects the nutritional composition of insect larvae [
23,
39,
40,
41,
42]. In particular, BSF larvae reared on diets containing increasing levels of
S. limacinum had proportionally higher nutritional value, both in terms of protein and fat content.
The protein content of the larvae was significantly influenced by the increasing proportion of microalgae and reached the highest content in larvae reared on the 25S and 25I diets (18.3% CP). This trend was linear with the increase in protein content in the proximate composition of the diet. The protein content determined in the present study was lower than that reported by several authors [
19,
34,
93], but in these papers, the BSF larvae were reared on much higher protein diets, which may have influenced the nutritional composition of the larvae.
A different trend was observed in the lipid content of the larvae, which changed with increasing levels of S. limacinum, with 20I and 25I having the highest levels (7.74 and 7.93 g/100 g, respectively) and were not influenced by increasing levels of I. galbana. Although the diets containing I. galbana had a significantly higher lipid content than the diets containing S. limacinum, the larvae had similar lipid levels in their biochemical composition to the larvae reared on 5S and 10S diets.
The results of the LCA show that the calculated environmental impacts are high compared to the values given in the literature for BSF production based on various substrates [
48] for all impact categories considered. This is mainly due to the high impact of microalgae production and is in line with the results of other studies [
94,
95]. In one research study, microalgae were considered a less environmentally friendly substitute for insect meals [
49]. Therefore, it is very likely that the use of microalgae for insect production increases the impact of insect production. In order to properly evaluate the different solutions, the nutritional quality aspects should be considered at the same time, as reported in the present study. It should also be emphasized that the impact of fishmeal is in some cases higher than that of 5S and 5I86 and that the negative impact of fishmeal on the decline of marine fish stocks is not considered in the LCAs.
To summarize, the data available in the literature do not allow us to say which is the best growth substrate for BSF larvae. Therefore, the results of this work can be considered an enrichment of the database, collecting information on how the performance of BSF larvae can be influenced by the substrates. Also, in terms of LCA, it might be useful to limit the microalgae content or to choose a more sustainable microalgae production. The latter will probably be possible in the future with the improvement of microalgae production technology. It should be noted that in certain scenarios where the side streams are also recycled, such as spent substrate as compost, or where insect production is considered as an alternative waste disposal option, a significant reduction in impact could be achieved.
Although this study was conducted under laboratory conditions, it provides a good perspective on the use of BSF as a by-product degrader.
5. Conclusions
As part of the Nutrifish project, authors analyzed the influence of these microalgae-enriched substrates on the fatty acid profile of BSF prepupae [
57] and the occurrence of antibiotic resistance genes in this BSF larvae rearing chain [
96]. In addition, BSF was reared on these diets to study its use as a sustainable terrestrial ingredient for aquafeed production [
25,
43] and to investigate the physiological and behavioral responses of zebrafish to insect-based diets [
77]. Last but not least, this work evaluates the growth performance of BSF and their impact on the environment when reared on potentially environmentally benign feed substrates (microalgae-enriched coffee silverskin) in order to propose them as an alternative feed ingredient in aquaculture. This study is important because several studies on the rearing of BSF and the evaluation of by-products used as feed have been conducted to date [
15,
17,
69,
70], but there is still little information on BSF. Considering the amount of coffee produced and therefore the amount of its by-products, as well as the pollution caused by coffee mites that are simply discarded into the environment, the need for different utilization alternatives to both reduce the environmental impact and increase the added value of CB is obvious.
The choice of rearing substrate for BSF is very important for growth performance and for an environmentally friendly product. In this work, it was confirmed that the by-product coffee silverskin, which is a major environmental hazard when used as a feed additive to be upgraded, contains low amounts of proteins and lipids to ensure good growth performance of BSF. However, by enriching this substrate with S. limacinum and I. galbana, two ingredients that contain the right amount of nutrients, these problems could be solved. Of the two microalgae, S. limacinum proved to be more beneficial than I. galbana.
Considering that the rearing substrate for BSF also contributes significantly to the environmental impact of larval production, as shown by the LCA results, it is very important to study it in depth, from the sustainability of the substrate components to its effect on the insects. This is necessary to better evaluate the ecological benefits of insects as alternative feed ingredients. As the sustainability of current
S. limacinum and
I. galbana production is limited, the percentage of substrate components should be limited. Moreover, a study proposed the inclusion of spirulina in the coffee-silverskin to enrich BSF final biomass in terms of nutritional value as an alternative aquafeed ingredient in commercial trout production [
78]. Further investigations are currently being performed to use agri-food leftovers (tomato, spinach, chickpeas, peas, and wheat), focusing on their availability in a defined geographical area (Regione Marche, Italy), as rearing substrates for BSF larvae to use as non-ruminant livestock feed. Based on the present findings, further research is needed to scale up BSF larvae production and assess the environmental impacts and sustainability of large-scale farming of BSF.


 ,
, 



{kind=link}