



Mulberry Branch Extracts Enhance the Antioxidant Capacity of Broiler Breast Muscle by Activating the Nrf2 and Cytochrome P450 Signaling Pathway


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mulberry Branch Extraction Procedure
2.2. Animals, Diets, and Sample Collection
2.3. Muscle Antioxidative Status Measurements
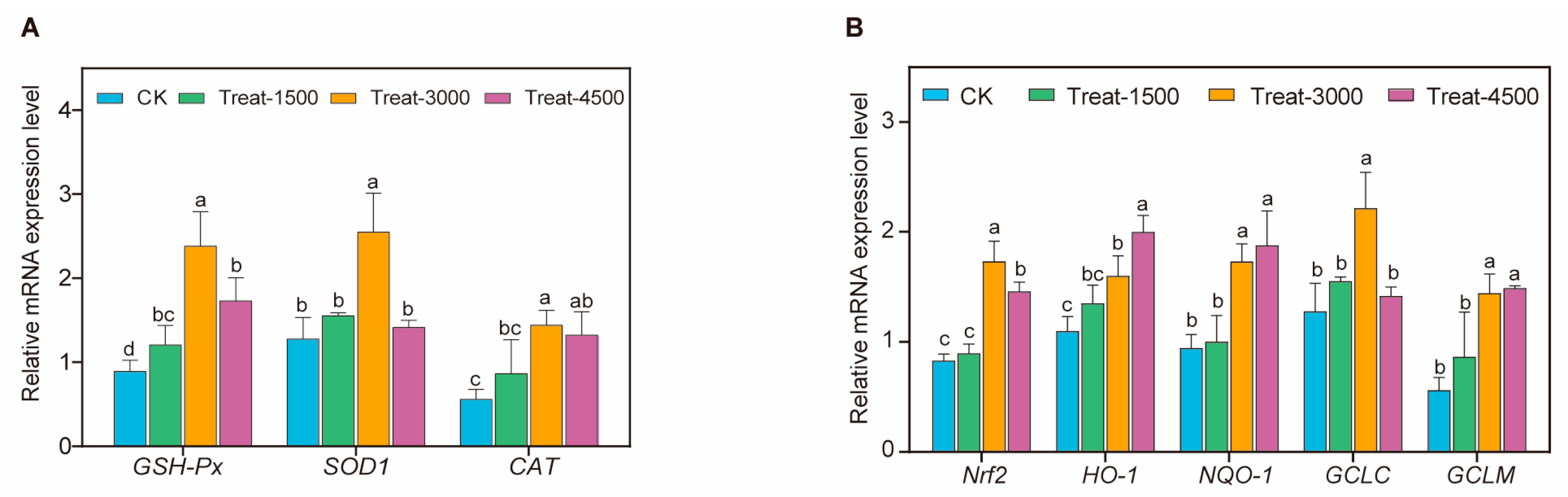
2.4. Real-Time Quantitative PCR of Key Genes’ mRNA Expression
2.5. Widely Targeted Metabolomics Analysis
2.6. Statistical Analysis
3. Results
3.1. Effects of MBEs on MDA Content and Antioxidant Enzyme Activities of Broiler Breast Muscle
3.2. Antioxidant-Related Gene Expression
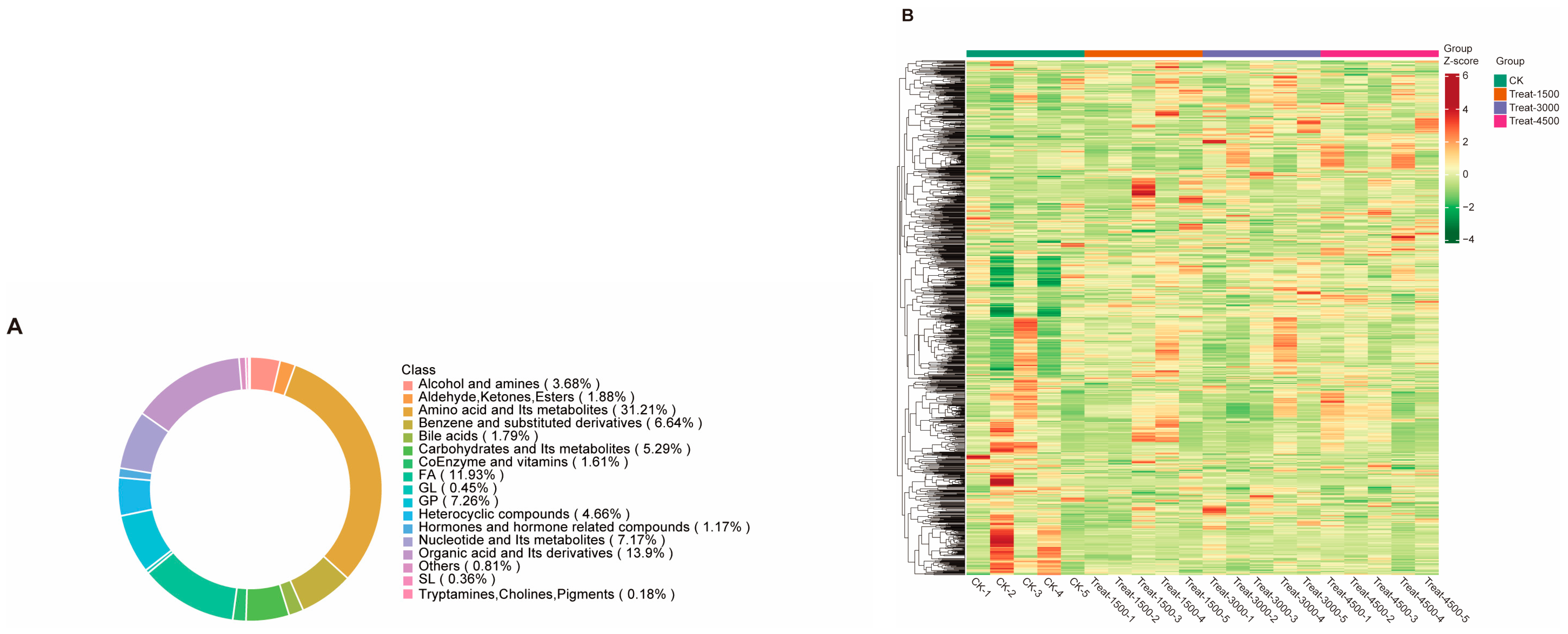
3.3. The Effects of the Addition of MBEs on the Metabolism of Broiler Breast Muscle
3.3.1. Full Mass Spectrometric and Dynamic Changes in Metabolites in Broiler Breast Muscle
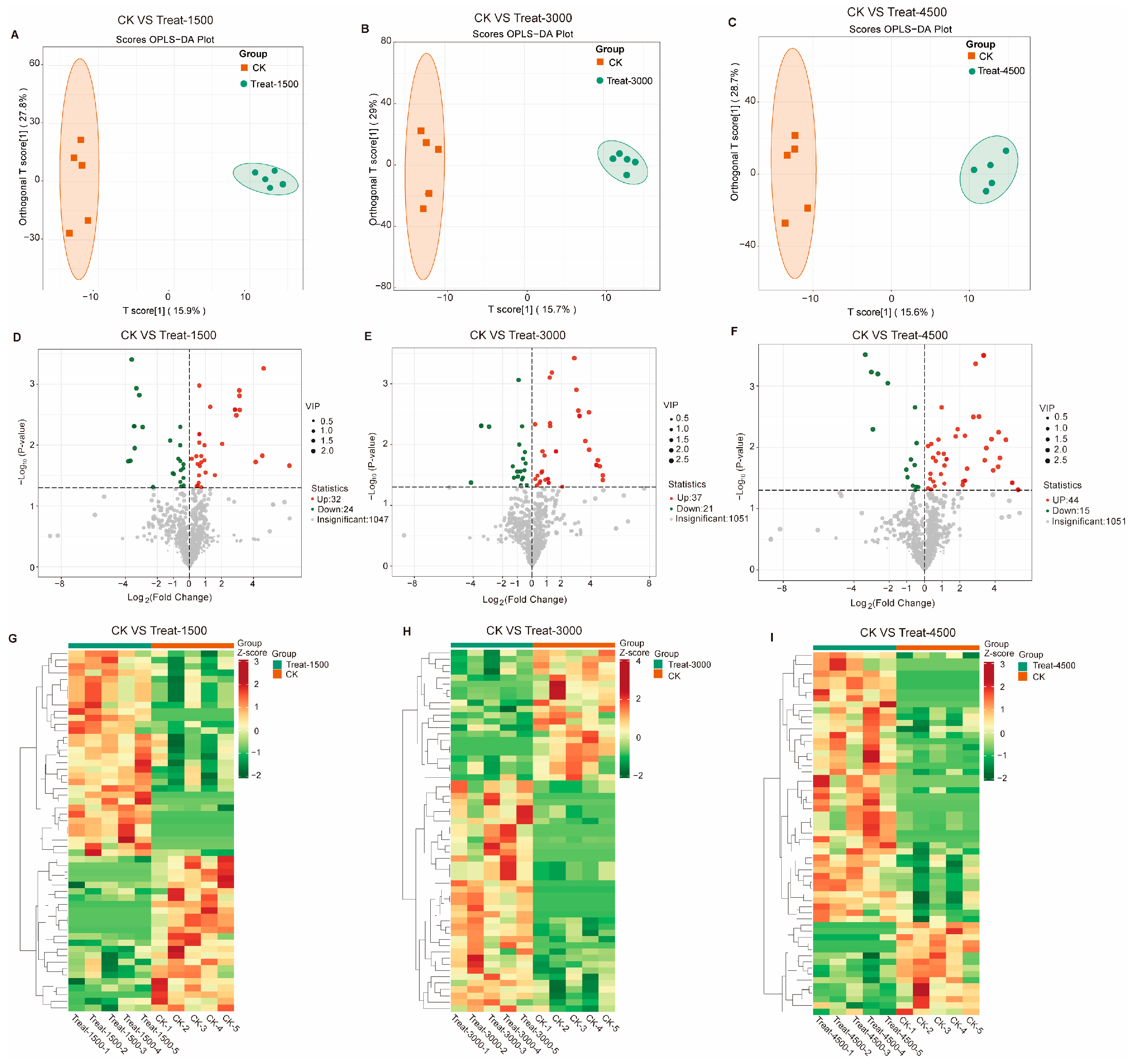
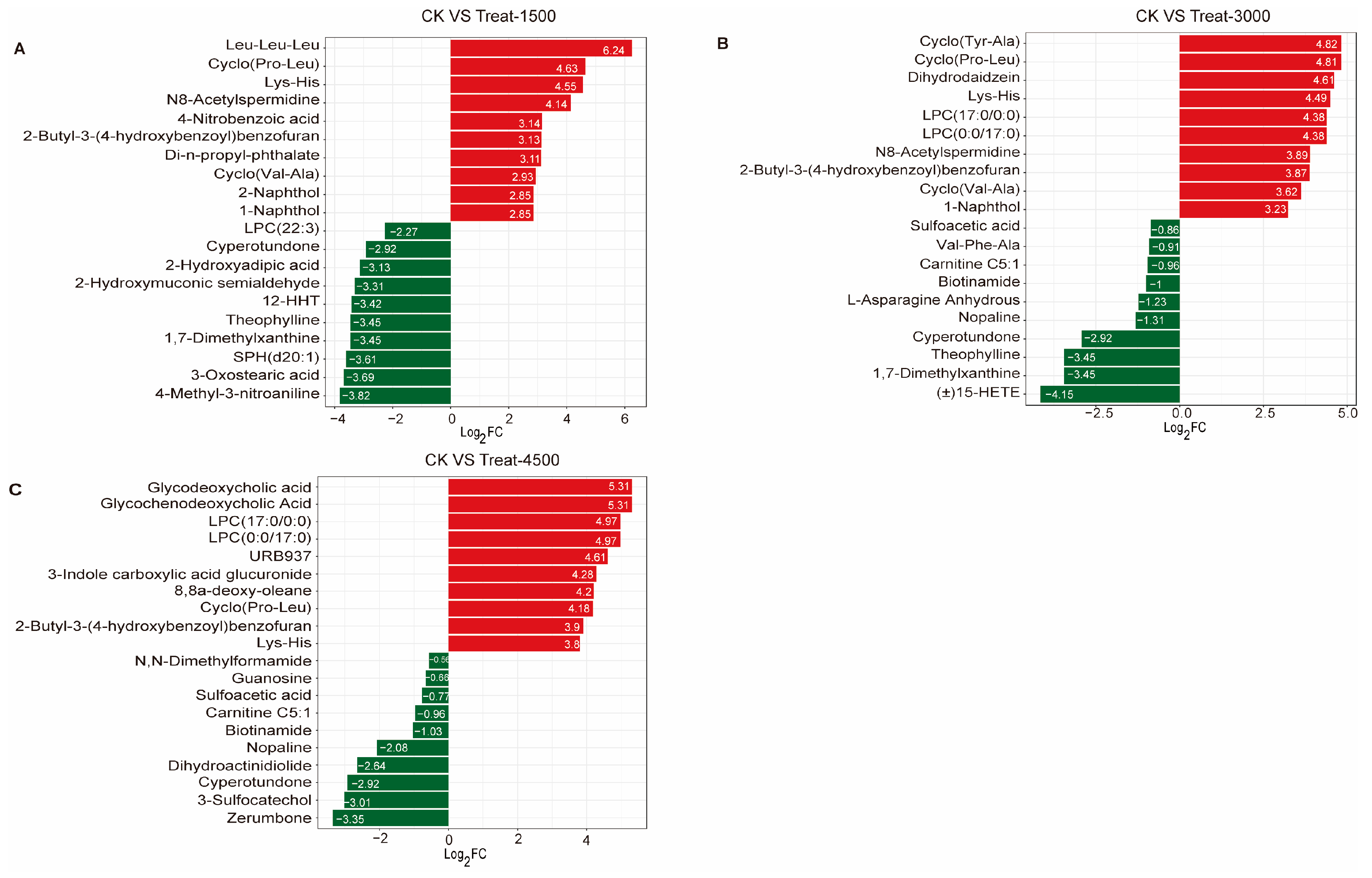
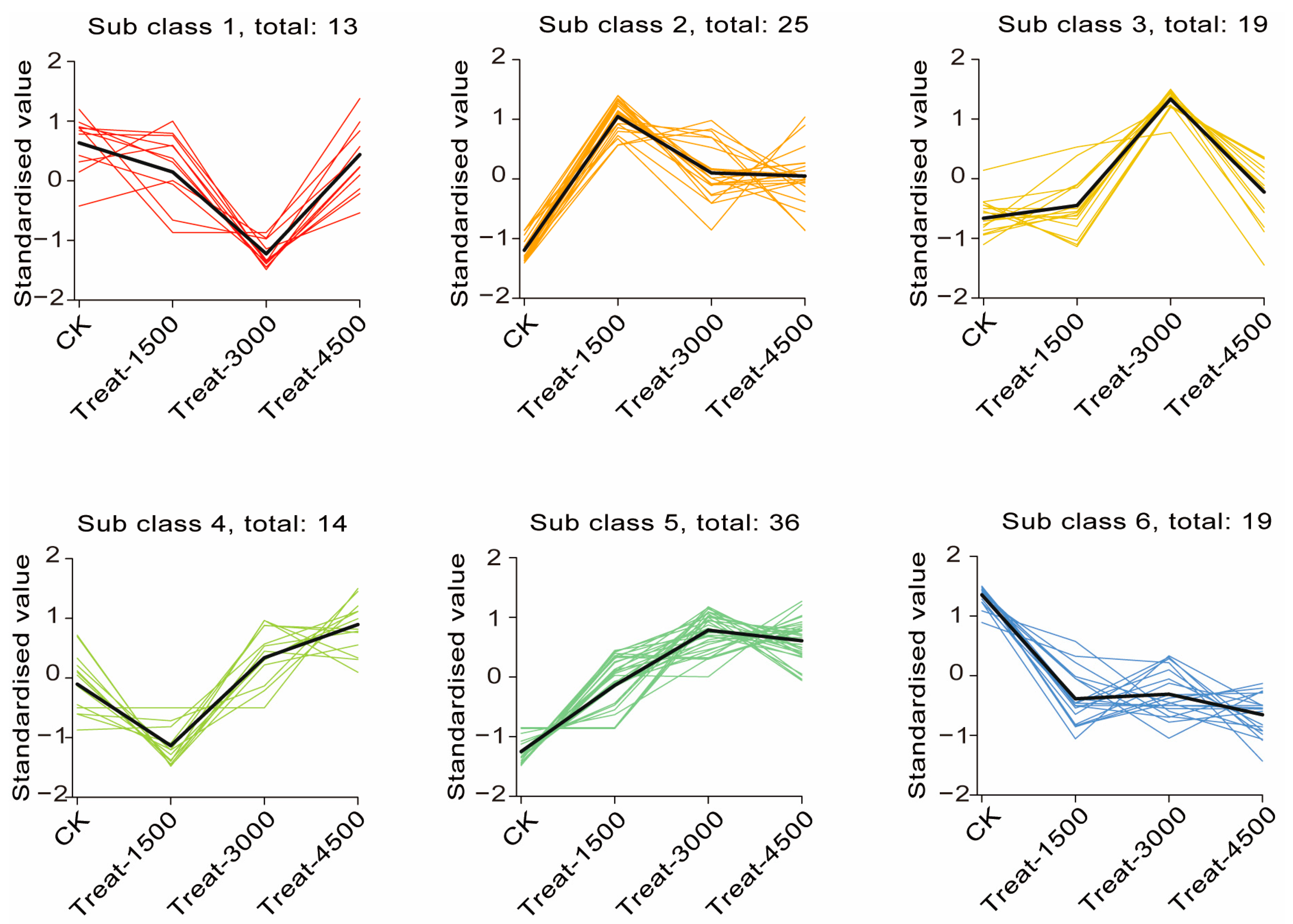
3.3.2. Multivariate Data Analysis and Identification of Characteristic Metabolites Between Groups
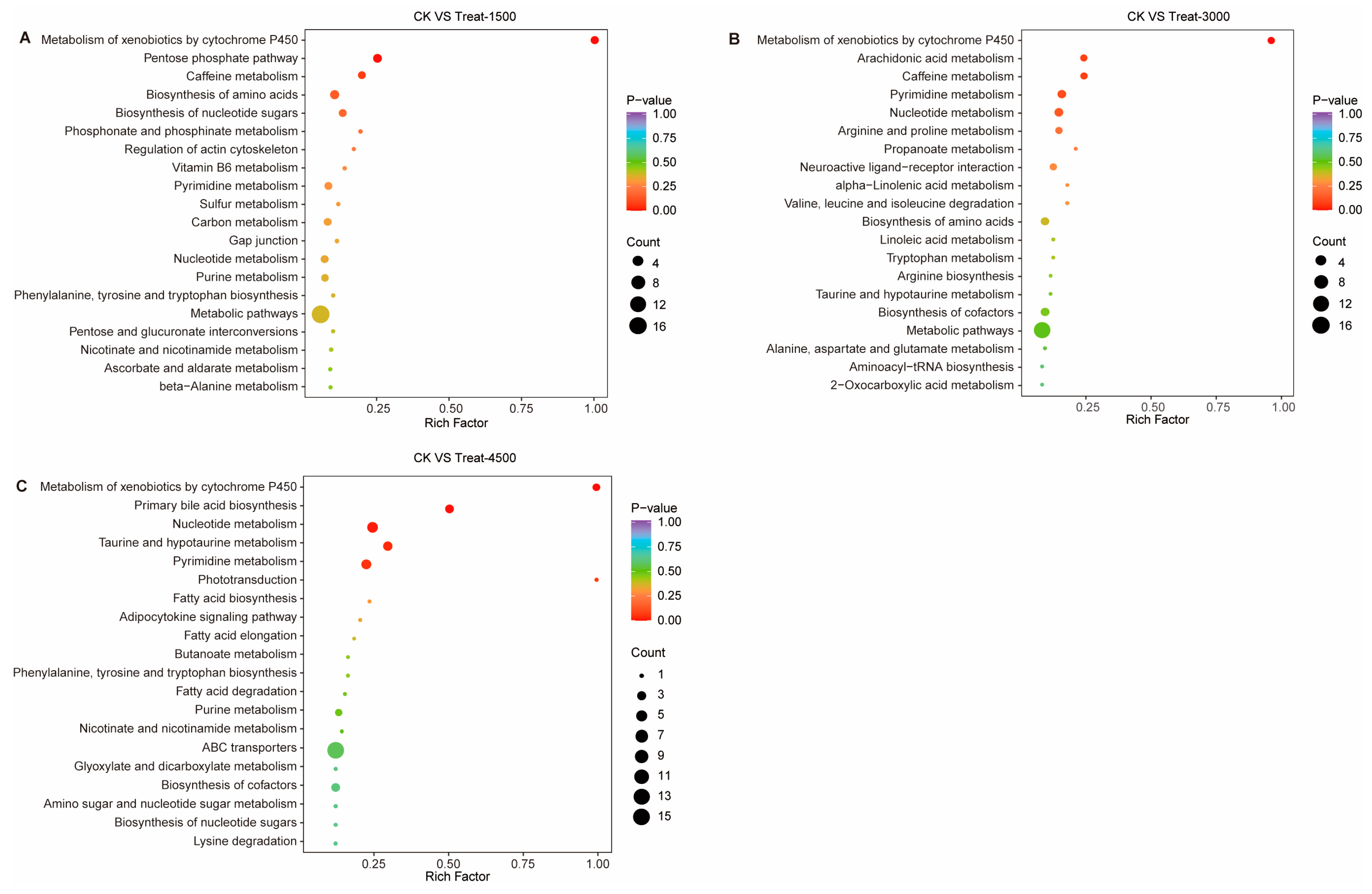
3.3.3. Effects of MBEs on Metabolic Pathways in Broiler Breast Muscle
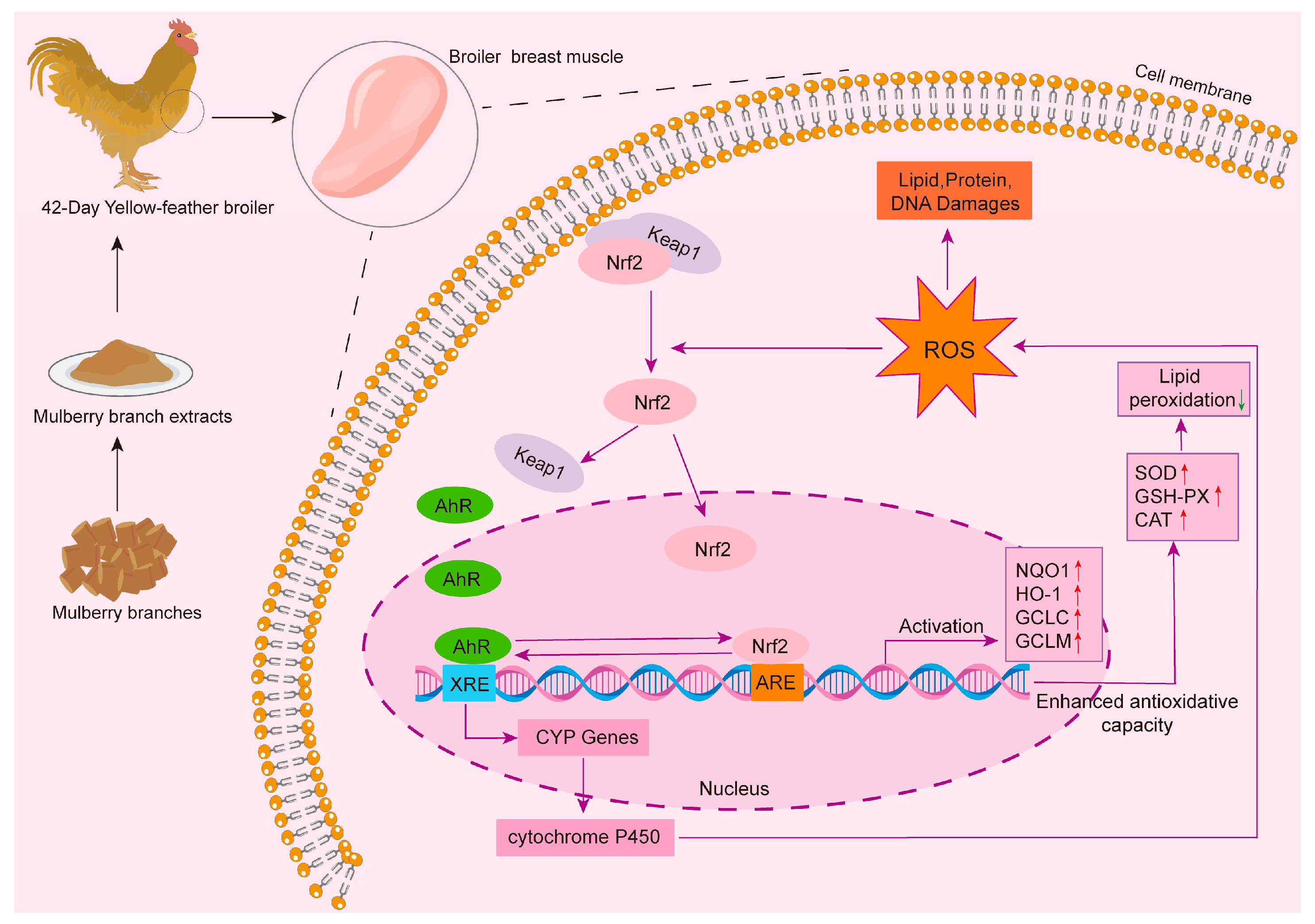
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pereira, P.M.; Vicente, A.F. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 2013, 93, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Soglia, F.; Madruga, M.; Carvalho, L.; Ida, E.; Estévez, M. Wooden-Breast, White Striping, and Spaghetti Meat: Causes, Consequences and Consumer Perception of Emerging Broiler Meat Abnormalities. Compr. Rev. Food Sci. Food Saf. 2019, 18, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Katafuchi, A.; Kamegawa, M.; Goto, S.; Kuwahara, D.; Osawa, Y.; Shimamoto, S.; Ishihara, S.; Ohtsuka, A.; Ijiri, D. Effects of Cyclic High Ambient Temperature on Muscle Imidazole Dipeptide Content in Broiler Chickens. J. Poult. Sci. 2024, 61, 2024004. [Google Scholar] [CrossRef]
- Xing, T.; Gao, F.; Tume, R.K.; Zhou, G.; Xu, X. Stress Effects on Meat Quality: A Mechanistic Perspective. Compr. Rev. Food Sci. Food Saf. 2019, 18, 380–401. [Google Scholar] [CrossRef] [PubMed]
- Hastie, M.; Ha, M.; Jacob, R.H.; Hepworth, G.; Torrico, D.D.; Warner, R.D. High consumer acceptance of mutton and the influence of ageing method on eating quality. Meat Sci. 2022, 189, 108813. [Google Scholar] [CrossRef] [PubMed]
- Gou, Z.; Abouelezz, K.F.M.; Fan, Q.; Li, L.; Lin, X.; Wang, Y.; Cui, X.; Ye, J.; Masoud, M.A.; Jiang, S.; et al. Physiological effects of transport duration on stress biomarkers and meat quality of medium-growing Yellow broiler chickens. Animal 2021, 15, 100079. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, J.; Li, Q.; Xiang, X.; Yang, Z.; Wang, T. Effects of trans-anethole supplementation on serum lipid metabolism parameters, carcass characteristics, meat quality, fatty acid, and amino acid profiles of breast muscle in broiler chickens. Poult Sci 2021, 100, 101484. [Google Scholar] [CrossRef] [PubMed]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid-protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Pateiro, M.; Munekata, P.E.S.; Zhang, W.; Garcia-Oliveira, P.; Carpena, M.; Prieto, M.A.; Bohrer, B.; Lorenzo, J.M. Protein Oxidation in Muscle Foods: A Comprehensive Review. Antioxidants 2021, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, X.; Chen, D.; Yu, B.; Yin, J.; Huang, Z. Effects of dietary apple polyphenol supplementation on carcass traits, meat quality, muscle amino acid and fatty acid composition in finishing pigs. Food Funct. 2019, 10, 7426–7434. [Google Scholar] [CrossRef] [PubMed]
- Manessis, G.; Kalogianni, A.I.; Lazou, T.; Moschovas, M.; Bossis, I.; Gelasakis, A.I. Plant-Derived Natural Antioxidants in Meat and Meat Products. Antioxidants 2020, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, X.; Liu, G.; Li, J.; Zhang, J.; Cao, Y.; Miao, J. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L.). Molecules 2021, 26, 7574. [Google Scholar] [CrossRef]
- Batiha, G.E.; Al-Snafi, A.E.; Thuwaini, M.M.; Teibo, J.O.; Shaheen, H.M.; Akomolafe, A.P.; Teibo, T.K.A.; Al-Kuraishy, H.M.; Al-Garbeeb, A.I.; Alexiou, A.; et al. Morus alba: A comprehensive phytochemical and pharmacological review. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 396, 1399–1413. [Google Scholar] [CrossRef] [PubMed]
- NY/T 3645-2020; Nutrient Requirements of Yellow Chickens. Ministry of Agriculture and Rural Affairs, PRC: Beijing, China, 2020.
- Nordberg, J.; Arnér, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Luo, X.G.; Ji, C.; Liu, B.; Yu, S.X. Effect of manganese supplementation and source on carcass traits, meat quality, and lipid oxidation in broilers. J. Anim. Sci. 2007, 85, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Weisshaar, N.; Hotz-Wagenblatt, A.; Madi, A.; Ma, S.; Mieg, A.; Hering, M.; Mohr, K.; Schlimbach, T.; Borgers, H.; et al. Skeletal muscle antagonizes antiviral CD8+ T cell exhaustion. Sci. Adv. 2020, 6, eaba3458. [Google Scholar] [CrossRef]
- Kumar, F.; Tyagi, P.K.; Mir, N.A.; Dev, K.; Begum, J.; Tyagi, P.K.; Biswas, A.; Sahu, B.; Dinani, O.P.; Sharma, D. Growth pattern, lipid composition, oxidation status, and serum biochemical profile of broiler chicken fed flaxseed meal for different durations. Lett. Anim. Biol. 2021, 1, 8–18. [Google Scholar] [CrossRef]
- Buiatte, V.; Dominguez, D.; Lesko, T.; Jenkins, M.; Chopra, S.; Lorenzoni, A.G. Inclusion of high-flavonoid corn in the diet of broiler chickens as a potential approach for the control of necrotic enteritis. Poult. Sci. 2022, 101, 101796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Li, X.; Zhao, J.; Wang, Y.; Hao, X.; Liu, K.; Liu, H. Protective effects of chlorogenic acid on the meat quality of oxidatively stressed broilers revealed by integrated metabolomics and antioxidant analysis. Food Funct. 2022, 13, 2238–2252. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Quality of chicken breast meat improved by dietary pterostilbene referring to up-regulated antioxidant capacity and enhanced protein structure. Food Chem. 2023, 405, 134848. [Google Scholar] [CrossRef]
- Zhang, S.; Pan, P.; Xie, H.; Wei, C.; Wang, Q.; Yang, B.; Sun, Y.; Li, Y.; Luo, Y.; Song, Y.; et al. Resveratrol improves meat quality traits by activating the lncRNAs-KEAP1-NRF2 axis in pigs. Meat Sci. 2024, 209, 109411. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.N.; Peng, Y.L.; Zhao, Y.P.; Cheng, X.F.; Zhou, Y.; Chai, C.L.; Zeng, L.S.; Pan, M.H.; Xu, L. Comparison of the Hepatoprotective Effects of the Three Main Stilbenes from Mulberry Twigs. J Agric Food Chem 2019, 67, 5521–5529. [Google Scholar] [CrossRef]
- Zhong, Y.; Tong, F.; Yan, J.; Tan, H.; Abudurexiti, A.; Zhang, R.; Lei, Y.; Li, D.; Ma, X. Differences in the flavonoid composition of the leaves, fruits, and branches of mulberry are distinguished based on a plant metabolomics approach. Open Life Sci 2024, 19, 20220886. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, X.; Tian, Y.; Lin, Q.; Xie, H.; Lu, W.; Chi, Y.; Chen, D. Lyophilized aqueous extracts of Mori Fructus and Mori Ramulus protect Mesenchymal stem cells from •OH-treated damage: Bioassay and antioxidant mechanism. BMC Complement. Altern. Med. 2017, 17, 242. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Ma, Y.; Zou, P.; Cheng, G.; Zhou, J.; Cai, S. Effects of Different Oligochitosans on Isoflavone Metabolites, Antioxidant Activity, and Isoflavone Biosynthetic Genes in Soybean (Glycine max) Seeds during Germination. J. Agric. Food Chem. 2019, 67, 4652–4661. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H. Dietary Curcumin Alleviated Acute Ileum Damage of Ducks (Anas platyrhynchos) Induced by AFB1 through Regulating Nrf2-ARE and NF-κB Signaling Pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef]
- Wei, X. Preparation of Mulberry Twig Extract and Its Application in Broiler Breeding. Master’s Thesis, Northwest A&F University, Xianyang, China, 2024. [Google Scholar]
- Cheng, K.; Yu, C.; Li, Z.; Li, S.; Yan, E.; Song, Z.; Zhang, H.; Zhang, L.; Wang, T. Resveratrol improves meat quality, muscular antioxidant capacity, lipid metabolism and fiber type composition of intrauterine growth retarded pigs. Meat Sci. 2020, 170, 108237. [Google Scholar] [CrossRef]
- Suman, S.P.; Joseph, P. Myoglobin chemistry and meat color. Annu. Rev. Food Sci. Technol. 2013, 4, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Wang, H.; Yu, Y.; Tao, F.; Yang, H.; Yang, S.; Qian, Y.; Li, H.; Yang, C. Dietary bamboo leaf flavonoids improve quality and microstructure of broiler meat by changing untargeted metabolome. J. Anim. Sci. Biotechnol. 2023, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Fu, B.; Zhou, D.; Li, E.; Li, Q.; Ruan, D.; Liao, S.; Zou, Y. Effects of mulberry leaf powder water extract supplementation on the growth performance, immunity, antioxidant, meat quality and intestinal microbiota of yellow feather broilers. J. Anim. Physiol. Anim. Nutr. 2024, 108, 1692–1703. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Ji, S.Y.; Lee, H.; Choi, S.H.; Kwon, C.Y.; Kim, S.Y.; Lee, E.T.; Choo, S.T.; Kim, G.Y.; Choi, Y.H.; et al. Mori Ramulus Suppresses Hydrogen Peroxide-Induced Oxidative Damage in Murine Myoblast C2C12 Cells through Activation of AMPK. Int. J. Mol. Sci. 2021, 22, 11729. [Google Scholar] [CrossRef]
- Zhu, Y.; Han, Y.; Wang, W.; Liang, G.; Qi, J. Mulberry leaves attenuate D-galactose-induced aging in vivo and in vitro. J. Ethnopharmacol. 2023, 311, 116286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, C.; Xie, K.; Wang, J.; Pan, Z. Current State of Metabolomics Research in Meat Quality Analysis and Authentication. Foods 2021, 10, 2388. [Google Scholar] [CrossRef] [PubMed]
- Lobo, R.R.; Correia, B.S.B.; Peña-Bermúdez, Y.A.; Vincenzi, R.; da Silva, C.M.; Panosso, L.L.; Ceribeli, C.; Colnago, L.A.; Cardoso, D.R.; Berndt, A.; et al. Effects of dietary inclusion of yerba mate (Ilex paraguariensis) extract on lamb muscle metabolomics and physicochemical properties in meat. J. Anim. Sci. 2021, 99, skab244. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhuang, Z.; Zhao, Y.; Song, Z.; Xiao, W.; Wang, W.; Li, Y.; Huang, N.; Jia, J.; Liu, Z.; et al. Unprocessed Red Meat and Processed Meat Consumption, Plasma Metabolome, and Risk of Ischemic Heart Disease: A Prospective Cohort Study of UK Biobank. J. Am. Heart Assoc. 2023, 12, e027934. [Google Scholar] [CrossRef]
- Ramírez-Zamudio, G.D.; Silva, L.H.P.; Vieira, N.M.; Vilela, R.S.R.; Assis, D.E.F.; Assis, G.J.F.; Estrada, M.M.; Rodrigues, R.T.S.; Duarte, M.S.; Chizzotti, M.L. Effect of short-term dietary protein restriction before slaughter on meat quality and skeletal muscle metabolomic profile in culled ewes. Livest. Sci. 2022, 261, 104956. [Google Scholar] [CrossRef]
- Qie, M.; Li, T.; Liu, C.C.; Zhao, Y. Direct analysis in real time high-resolution mass spectrometry for authenticity assessment of lamb. Food Chem. 2022, 390, 133143. [Google Scholar] [CrossRef] [PubMed]
- López, C.M.; Bru, E.; Vignolo, G.M.; Fadda, S.G. Identification of small peptides arising from hydrolysis of meat proteins in dry fermented sausages. Meat Sci. 2015, 104, 20–29. [Google Scholar] [CrossRef]
- Mora, L.; Gallego, M.; Escudero, E.; Reig, M.; Aristoy, M.C.; Toldrá, F. Small peptides hydrolysis in dry-cured meats. Int. J. Food Microbiol. 2015, 212, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Heres, A.; Mora, L.; Toldrá, F. Bioactive and Sensory Di- and Tripeptides Generated during Dry-Curing of Pork Meat. Int. J. Mol. Sci. 2023, 24, 1574. [Google Scholar] [CrossRef]
- Chen, X.; Xiang, L.; Jia, G.; Liu, G.; Zhao, H.; Huang, Z. Effects of dietary leucine on antioxidant activity and expression of antioxidant and mitochondrial-related genes in longissimus dorsi muscle and liver of piglets. Anim. Sci. J. = Nihon Chikusan Gakkaiho 2019, 90, 990–998. [Google Scholar] [CrossRef]
- Sun, B.; Sun, Y.; Han, X.; Ma, Q.; Meng, Q. Leucine Supplementation Alleviates Immune and Antioxidant Function Damage in Adult Rats Induced by Early Weaning. J. Nutr. 2023, 153, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Rong, N.; Senthil Kumar, H.V.; Swedick, S.; Samuel, R.Z.; Mehrotra, P.; Toftegaard, J.; Rajabian, N.; Thiyagarajan, R.; Podder, A.K.; et al. Proline restores mitochondrial function and reverses aging hallmarks in senescent cells. Cell Rep. 2024, 43, 113738. [Google Scholar] [CrossRef]
- Yang, P.; Deng, F.; Yuan, M.; Chen, M.; Zeng, L.; Ouyang, Y.; Chen, X.; Zhao, B.; Yang, Z.; Tian, Z. Metabolomics reveals the defense mechanism of histidine supplementation on high-salt exposure-induced hepatic oxidative stress. Life Sci. 2023, 314, 121355. [Google Scholar] [CrossRef] [PubMed]
- Loida, P.J.; Sligar, S.G. Molecular recognition in cytochrome P-450: Mechanism for the control of uncoupling reactions. Biochemistry 1993, 32, 11530–11538. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome p450 and chemical toxicology. Chem. Res. Toxicol. 2008, 21, 70–83. [Google Scholar] [CrossRef]
- Harskamp, J.; Britz-McKibbin, P.; Wilson, J.Y. Functional screening of cytochrome P450 activity and uncoupling by capillary electrophoresis. Anal. Chem. 2012, 84, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Hu, L.; Scrivens, P.J.; Batist, G. Transcriptional regulation of NF-E2 p45-related factor (NRF2) expression by the aryl hydrocarbon receptor-xenobiotic response element signaling pathway: Direct cross-talk between phase I and II drug-metabolizing enzymes. J. Biol. Chem. 2005, 280, 20340–20348. [Google Scholar] [CrossRef] [PubMed]
- Köhle, C.; Bock, K.W. Coordinate regulation of Phase I and II xenobiotic metabolisms by the Ah receptor and Nrf2. Biochem. Pharmacol. 2007, 73, 1853–1862. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | 42–65 Days of Age | 66–90 Days of Age |
|---|---|---|
| Ingredients | ||
| Corn | 58.06 | 62 |
| Soybean meal (crude protein 43%) | 26 | 19.9 |
| Fish meal | 2 | 2 |
| DDGS (distilled dried grains with solubles) | 3 | 5 |
| Wheat bran | 4 | - |
| WMAR (Wheat Middling and Red Dog) | - | 4.16 |
| Soybean oil | 2 | 2.5 |
| Limestone | 1.2 | 1 |
| CaHPO4 | 1.5 | 1.2 |
| NaCl | 0.24 | 0.24 |
| Premix 1 | 2 | 2 |
| Total | 100 | 100 |
| Nutrient levels 2 | ||
| Metabolic energy, MJ/kg | 12.1 | 12.66 |
| Crude protein | 18.51 | 16.77 |
| Calcium | 0.98 | 0.82 |
| Total phosphorus | 0.67 | 0.58 |
| Methionine + cysteine | 0.7 | 0.64 |
| Lysine | 0.96 | 0.82 |
| Methionine | 0.32 | 0.30 |
| GeneBank ID | Name | Primer Sequence (5′-3′) |
|---|---|---|
| NM_205518 | β-actin | F: GCCAACAGAGAGAAGATGACAC R: GTAACACCATCACCAGAGTCCA |
| NM_205117.1 | Nrf2 | F: AGGAAGAAGGTGCTTTTCGCA R: TCTGTTCCTCTTCACTGCCAC |
| NM_001163245.1 | GSH-Px | F: CAAAGTTGCGGTCAGTGGA R: AGAGTCCCAGGCCTTTACTACTTTC |
| NM_205064.1 | SOD1 | F: TTGTCTGATGGAGATCATGGCTTC R: TGCTTGCCTTCAGGATTAAAGTGA |
| NM_001031215.2 | CAT | F: GGTTCGGTGGGGTTGTCTTT R: CACCAGTGGTCAAGGCATCT |
| NM_001277620.2 | NQO1 | F: TCGCCGAGCAGAAGAAGATTGAA R: CGGTGGTGAGTGACAGCATGG |
| NM_205344.2 | HO-1 | F: ACGAGTTCAAGCTGGTCACG R: GGATGCTTCTTGCCAACGAC |
| XM_046915268.1 | GCLC | F: TCTGTAGATGATCGAACGC R: TCCTTTATTAGGTGCTCGTAG |
| NM_001007953.2 | GCLM | F: GCTGCTAACTCACAATGACC R: TGCATGATATAGCCTTTGGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Qian, W.; Wei, X.; Qin, X.; Han, J.; Su, C.; Bao, L. Mulberry Branch Extracts Enhance the Antioxidant Capacity of Broiler Breast Muscle by Activating the Nrf2 and Cytochrome P450 Signaling Pathway. Animals 2024, 14, 3702. https://doi.org/10.3390/ani14243702
Shi X, Qian W, Wei X, Qin X, Han J, Su C, Bao L. Mulberry Branch Extracts Enhance the Antioxidant Capacity of Broiler Breast Muscle by Activating the Nrf2 and Cytochrome P450 Signaling Pathway. Animals. 2024; 14(24):3702. https://doi.org/10.3390/ani14243702
Chicago/Turabian StyleShi, Xiang, Wei Qian, Xinlan Wei, Xiaoqing Qin, Jinyan Han, Chao Su, and Lijun Bao. 2024. "Mulberry Branch Extracts Enhance the Antioxidant Capacity of Broiler Breast Muscle by Activating the Nrf2 and Cytochrome P450 Signaling Pathway" Animals 14, no. 24: 3702. https://doi.org/10.3390/ani14243702
APA StyleShi, X., Qian, W., Wei, X., Qin, X., Han, J., Su, C., & Bao, L. (2024). Mulberry Branch Extracts Enhance the Antioxidant Capacity of Broiler Breast Muscle by Activating the Nrf2 and Cytochrome P450 Signaling Pathway. Animals, 14(24), 3702. https://doi.org/10.3390/ani14243702