
Study of Variation of ACOX1 Gene Among Different Horse Breeds Maintained in Iran
 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
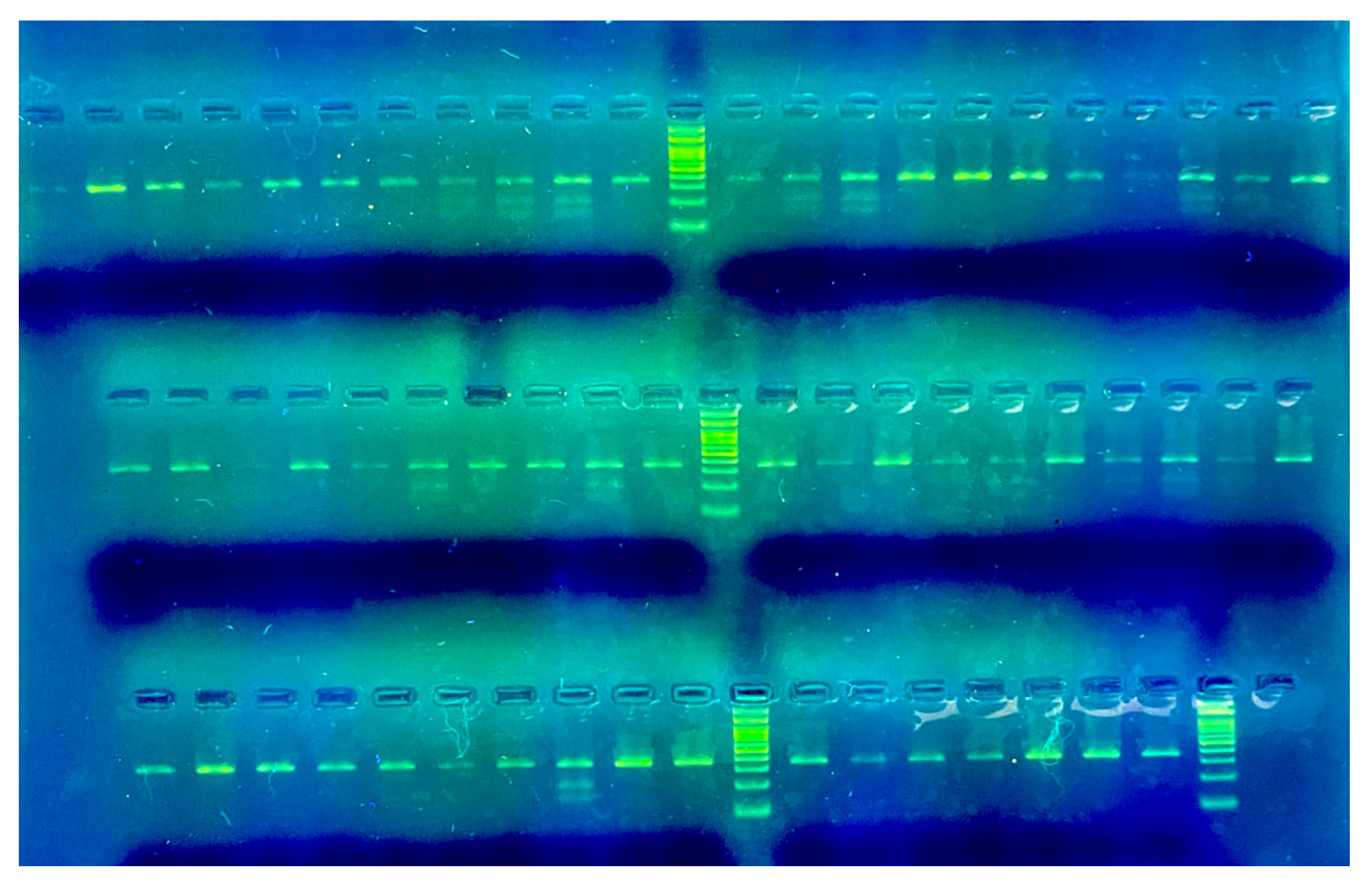
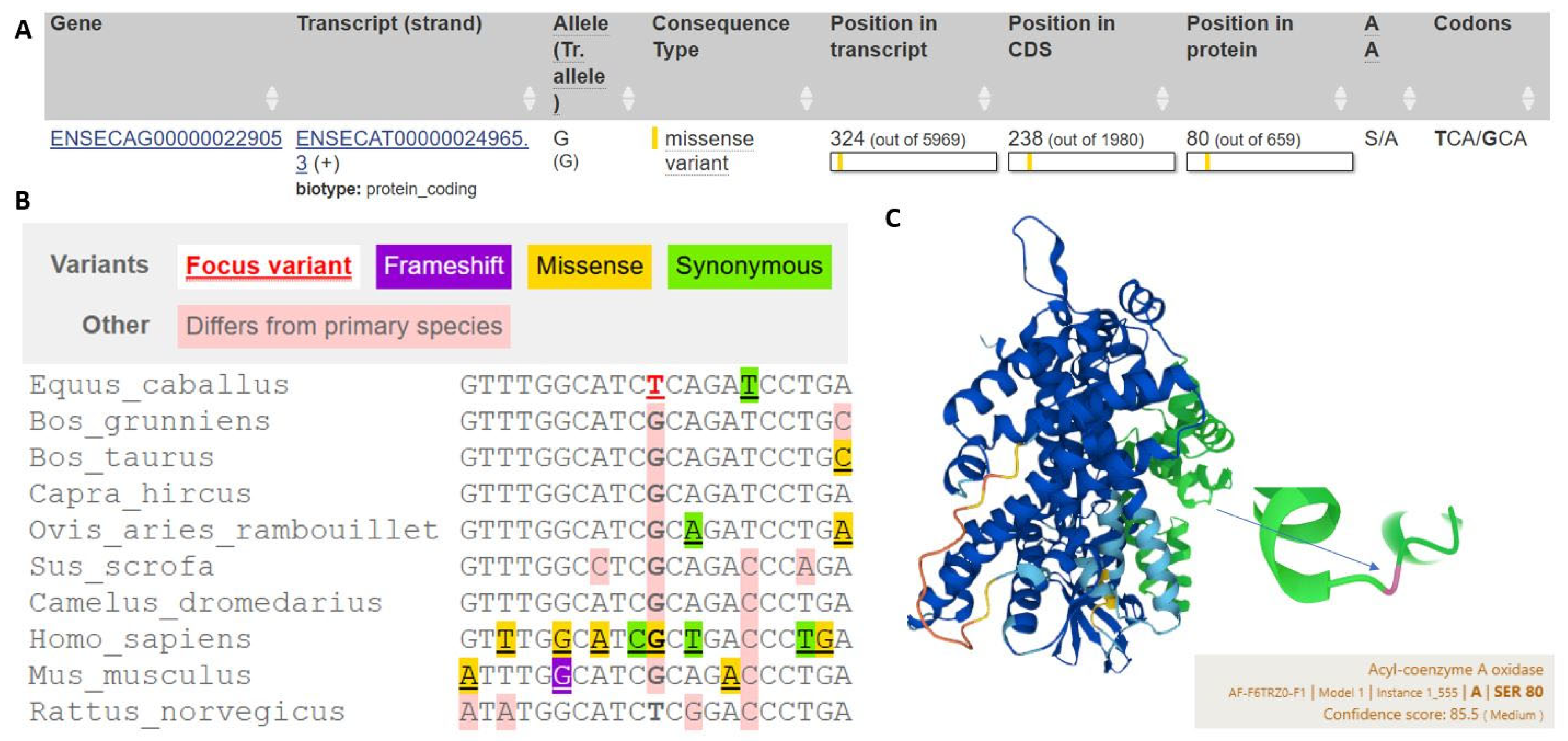
2.2. Identification of ACOX1 Polymorphism and Genotyping
3. Results
3.1. Genotype Distribution
3.2. The Chi-Square Tests for Independence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lönker, N.S.; Fechner, K.; Wahed, A.A.E. Horses as a Crucial Part of One Health. Vet. Sci. 2020, 7, 28. [Google Scholar] [CrossRef]
- Benedetti, B.; Felici, M.; Nanni Costa, L.; Padalino, B. A review of horse welfare literature from 1980 to 2023 with a text mining and topic analysis approach. Ital. J. Anim. Sci. 2023, 22, 1095–1109. [Google Scholar] [CrossRef]
- Endenburg, N. Perceptions and attitudes towards horses in European societies. Equine Vet. J. Suppl. 1999, 31, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.K.; Friend, T.H.; Evans, J.W.; Bushong, D.M. Behavioral assessment of horses in therapeutic riding programs. Appl. Anim. Behav. Sci. 1999, 63, 11–24. [Google Scholar] [CrossRef]
- Hanot, P.; Bayarsaikhan, J.; Guintard, C.; Haruda, A.; Mijiddorj, E.; Schafberg, R.; Taylor, W. Cranial shape diversification in horses: Variation and covariation patterns under the impact of artificial selection. BMC Ecol. Evol. 2021, 21, 178. [Google Scholar] [CrossRef] [PubMed]
- Warmuth, V.; Manica, A.; Eriksson, A.; Barker, G.; Bower, M. Autosomal genetic diversity in non-breed horses from eastern Eurasia provides insights into historical population movements. Anim. Genet. 2013, 44, 53–61. [Google Scholar] [CrossRef]
- Nagel, H.J.; Savier, M.M. The Arabian Horse. Nature’s Creation and the Arte of Breeding; Nawal Media: Spoleto, Italy, 2013. [Google Scholar]
- Ricard, A.; Robert, C.; Blouin, C.; Baste, F.; Torquet, G.; Morgenthaler, C.; Rivière, J.; Mach, N.; Mata, X.; Schibler, L.; et al. Endurance Exercise Ability in the Horse: A Trait with Complex Polygenic Determinism. Front. Genet. 2017, 8, 89. [Google Scholar] [CrossRef]
- Rovere, G.; Ducro, B.J.; van Arendonk, J.A.; Norberg, E.; Madsen, P. Analysis of competition performance in dressage and show jumping of Dutch Warmblood horses. J. Anim. Breed Genet. 2016, 133, 503–512. [Google Scholar] [CrossRef]
- Cassidy, R. The Sport of Kings: Kinship, Class and Thoroughbred Breeding in Newmarket; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Gaffney, B.; Cunningham, E.P. Estimation of genetic trend in racing performance of Thoroughbred horses. Nature 1988, 332, 722–724. [Google Scholar] [CrossRef]
- Katz, L.M.; Bayly, W.M.; Hines, M.T.; Sides, R.H. Differences in the ventilatory responses of horses and ponies to exercise of varying intensities. Equine Vet. J. Suppl. 1999, 31, 49–51. [Google Scholar] [CrossRef]
- Pösö, A.R.; Essén-Gustavsson, B.; Persson, S.G. Metabolic response to standardised exercise test in standardbred trotters with red cell hypervolaemia. Equine Vet. J. 1993, 25, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-D.; Park, J.; Ko, J.; Kim, B.C.; Kim, H.-S.; Ahn, K.; Do, K.-T.; Choi, H.; Kim, H.-M.; Song, S.; et al. Whole transcriptome analyses of six Thoroughbred horses before and after exercise using RNA-Seq. BMC Genom. 2012, 13, 473. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Mianji, G.; Nejati-Javaremi, A.; Farhadi, A. Genetic diversity, parentage verification and genetic bottlenecks evaluation in Iranian Turkmen horse breed. Genetika 2015, 51, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Pluta, M.; Bańka, K.; Ciesla, A.; Rogala, Ł. The state of breeding and use of Caspian horses in Europe and around the world. Acta Sci. Pol. Zootech. 2021, 19, 79–84. [Google Scholar] [CrossRef]
- Firouz, L. The Caspian Miniature Horse of Iran; International Caspian Stud Book: Lancashire, UK, 1972. [Google Scholar]
- Yousefi, N.; Mehrabani, H.; Nejati Javaremi, A.; Maloufi, F. A novel approach to establish breed type and standards for an equine breed: Persian Kurdish horse. J. Agric. Sci. Technol. 2020, 22, 1219–1233. [Google Scholar]
- Myćka, G.; Musiał, A.D.; Stefaniuk-Szmukier, M.; Piórkowska, K.; Ropka-Molik, K. Variability of ACOX1 gene polymorphisms across different horse breeds with regard to selection pressure. Animals 2020, 10, 2225. [Google Scholar] [CrossRef]
- Al Abri, M.; Holl, H.; Kalla, S.; Sutter, N.; Brooks, S. Whole genome detection of sequence and structural polymorphism in six diverse horses. PLoS ONE 2020, 15, e0230899. [Google Scholar] [CrossRef]
- Głażewska, I. Speculations on the origin of the Arabian horse breed. Livest. Sci. 2010, 129, 49–55. [Google Scholar] [CrossRef]
- Varanasi, U.; Chu, R.; Chu, S.; Espinosa, R.; LeBeau, M.M.; Reddy, J.K. Isolation of the human peroxisomal acyl-CoA oxidase gene: Organization, promoter analysis, and chromosomal localization. Proc. Natl. Acad. Sci. USA 1994, 91, 3107–3111. [Google Scholar] [CrossRef]
- Poirier, Y.; Antonenkov, V.D.; Glumoff, T.; Hiltunen, J.K. Peroxisomal β-oxidation—A metabolic pathway with multiple functions. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2006, 1763, 1413–1426. [Google Scholar] [CrossRef]
- Barish, G.D.; Narkar, V.A.; Evans, R.M. PPAR delta: A dagger in the heart of the metabolic syndrome. J. Clin. Investig. 2006, 116, 590–597. [Google Scholar] [CrossRef] [PubMed]
- El Hajj, H.I.; Vluggens, A.; Andreoletti, P.; Ragot, K.; Mandard, S.; Kersten, S.; Waterham, H.R.; Lizard, G.; Wanders, R.J.A.; Reddy, J.K.; et al. The inflammatory response in acyl-CoA oxidase 1 deficiency (pseudoneonatal adrenoleukodystrophy). Endocrinology 2012, 153, 2568–2575. [Google Scholar] [CrossRef] [PubMed]
- Boemer, F.; Detilleux, J.; Cello, C.; Amory, H.; Marcillaud-Pitel, C.; Richard, E.; Van Galen, G.; Van Loon, G.; Lefère, L.; Votion, D.-M. Acylcarnitines profile best predicts survival in horses with atypical myopathy. PLoS ONE 2017, 12, e0182761. [Google Scholar] [CrossRef] [PubMed]
- Fontanel, M.; Todd, E.; Drabbe, A.; Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Myćka, G.; Velie, B.D. Variation in the SLC16A1 and the ACOX1 genes is associated with gallop racing performance in Arabian horses. J. Equine Vet. Sci. 2020, 93, 103202. [Google Scholar] [CrossRef]
- Parker, K.L.; Gillingham, M.P.; Hanley, T.A.; Robbins, C.T. Seasonal patterns in body mass, body composition, and water transfer rates of free-ranging and captive black-tailed deer (Odocoileus hemionus sitkensis) in Alaska. Can. J. Zool. 1993, 71, 1397–1404. [Google Scholar] [CrossRef]
- Gu, J.; Orr, N.; Park, S.D.; Katz, L.M.; Sulimova, G.; MacHugh, D.E.; Hill, E.W. A genome scan for positive selection in Thoroughbred horses. PLoS ONE 2009, 4, e5767. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TT | GG | TG | Number of Horses | G * | T * | HWE | ||
|---|---|---|---|---|---|---|---|---|
| Arabian | 6 (7%) | 63 (70%) | 21 (23%) | 90 | 0.82 | 0.18 | 0.03 | |
| Caspian | 2 (10%) | 11 (52%) | 8 (38% | 21 | 0.71 | 0.29 | 0.75 | |
| KWPN | 10 (20%) | 31 (62%) | 9 (18%) | 50 | 0.71 | 0.29 | 0.000069 | |
| HORSE BREEDS | Kurdish | 1 (2%) | 63 (86%) | 9 (12%) | 73 | 0.92 | 0.08 | 0.32 |
| Thoroughbred | 8 (23%) | 16 (46%) | 11 (31%) | 35 | 0.61 | 0.39 | 0.046 | |
| Turkmen | 13 (24%) | 21 (38%) | 21 (38%) | 55 | 0.57 | 0.43 | 0.102 | |
| TOTAL | 40 (12.34%) | 205 (63.27%) | 79 (24.38%) | 324 | 0.75 | 0.25 | 0.000000 |
| Arabian | Caspian | KWPN | Kurdish | Thoroughbred | Turkmen | |
|---|---|---|---|---|---|---|
| Arabian | ---- | |||||
| Caspian | ns | ----- | ||||
| KWPN | ns | ns | ---- | |||
| Kurdish | 0.0356 | 0.0028 | 0.0007 | ---- | ||
| Thoroughbred | 0.0117 | ns | ns | <0.0001 | ---- | |
| Turkmen | 0.0003 | ns | 0.0319 | <0.0001 | ns | ---- |
| Genotypes | Sex | |||
| Mare Count | Stallion Count | |||
| GG | Arabian | 35 | 28 | |
| Caspian | 7 | 4 | ||
| KWPN | 14 | 17 | ||
| Kurdish | 21 | 42 | ||
| Thoroughbred | 4 | 12 | ||
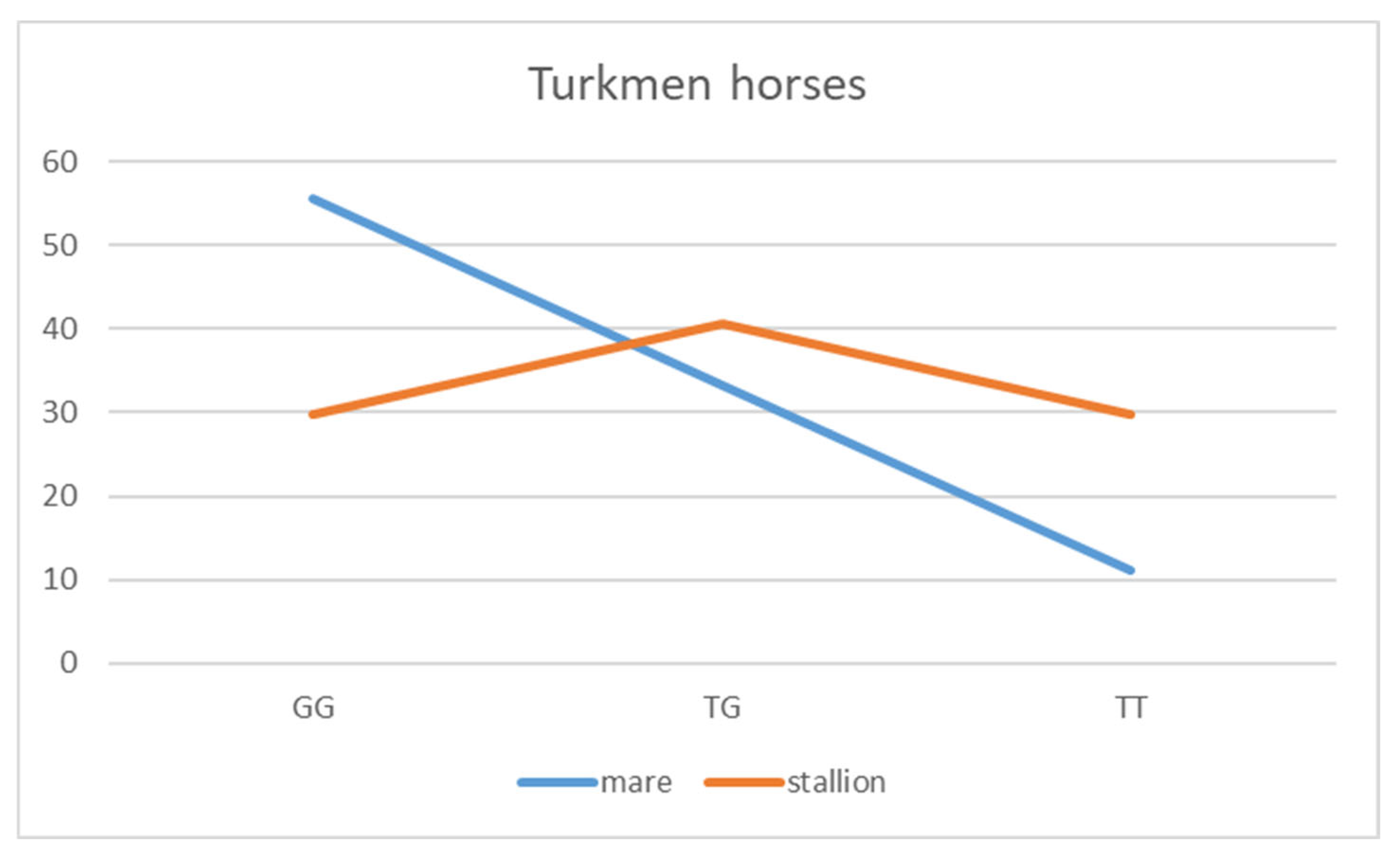
| Turkmen | 10 (55.5) | 11 (29.7) | ||
| TG | Arabian | 12 | 9 | |
| Caspian | 4 | 4 | ||
| KWPN | 6 | 3 | ||
| Kurdish | 4 | 5 | ||
| Thoroughbred | 4 | 7 | ||
| Turkmen | 6 (33.3) | 15 (40.6) | ||
| TT | Arabian | 3 | 3 | |
| Caspian | 1 | 1 | ||
| KWPN | 4 | 6 | ||
| Kurdish | 0 | 1 | ||
| Thoroughbred | 3 | 5 | ||
| Turkmen | 2 (11.1) | 11 (29.7) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boozarjomehri Amnieh, S.; Hassanpour, A.; Moghaddam, S.; Sakhaee, F.; Ropka-Molik, K. Study of Variation of ACOX1 Gene Among Different Horse Breeds Maintained in Iran. Animals 2024, 14, 3566. https://doi.org/10.3390/ani14243566
Boozarjomehri Amnieh S, Hassanpour A, Moghaddam S, Sakhaee F, Ropka-Molik K. Study of Variation of ACOX1 Gene Among Different Horse Breeds Maintained in Iran. Animals. 2024; 14(24):3566. https://doi.org/10.3390/ani14243566
Chicago/Turabian StyleBoozarjomehri Amnieh, Shayan, Ali Hassanpour, Sina Moghaddam, Fatemeh Sakhaee, and Katarzyna Ropka-Molik. 2024. "Study of Variation of ACOX1 Gene Among Different Horse Breeds Maintained in Iran" Animals 14, no. 24: 3566. https://doi.org/10.3390/ani14243566
APA StyleBoozarjomehri Amnieh, S., Hassanpour, A., Moghaddam, S., Sakhaee, F., & Ropka-Molik, K. (2024). Study of Variation of ACOX1 Gene Among Different Horse Breeds Maintained in Iran. Animals, 14(24), 3566. https://doi.org/10.3390/ani14243566