
Genome-Wide Selection Signals Reveal Candidate Genes Associated with Plateau Adaptation in Tibetan Sheep
 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Resequencing
2.2. Quality Control and Reference Genome Alignment
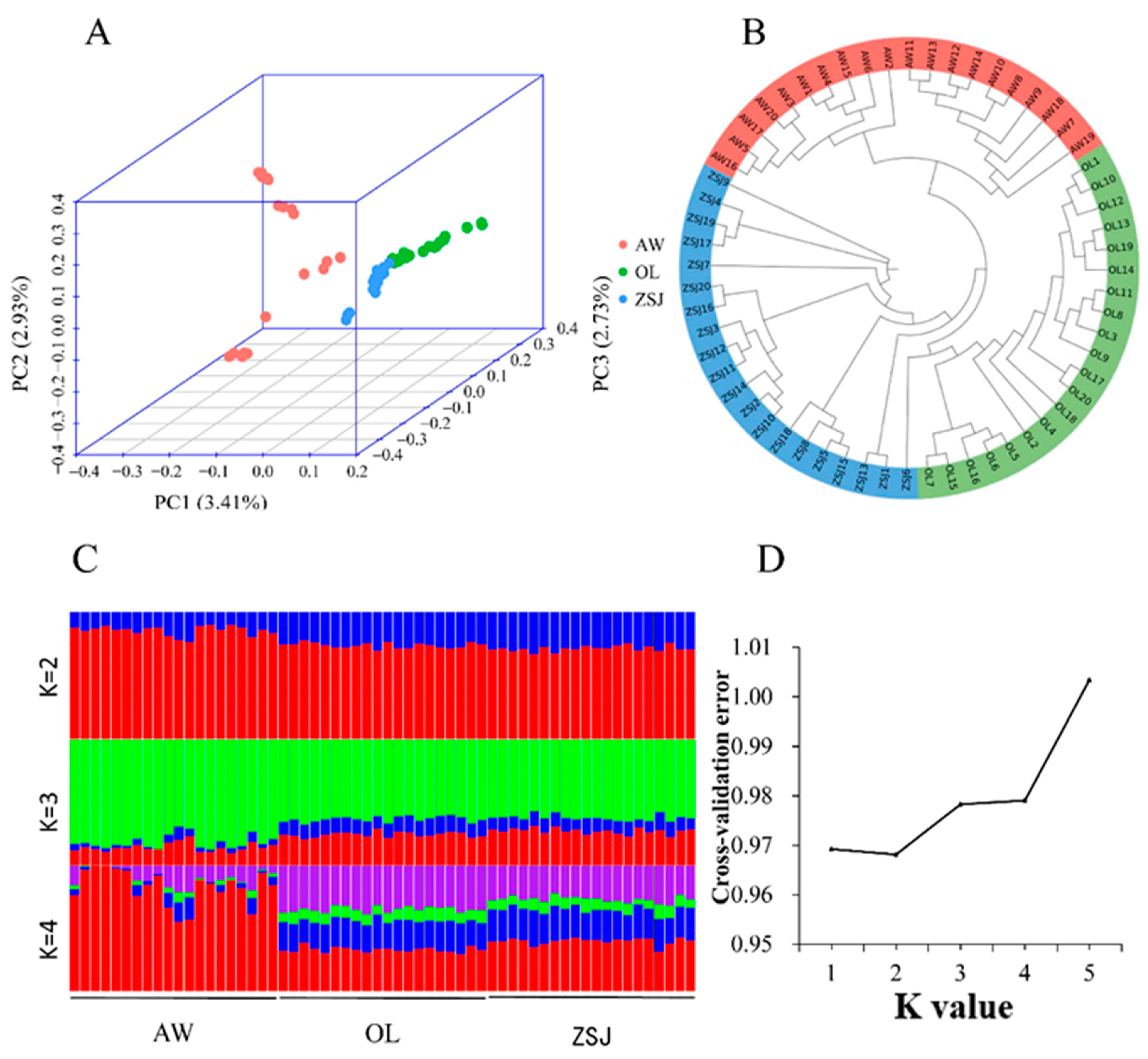
2.3. Population Structure Analysis
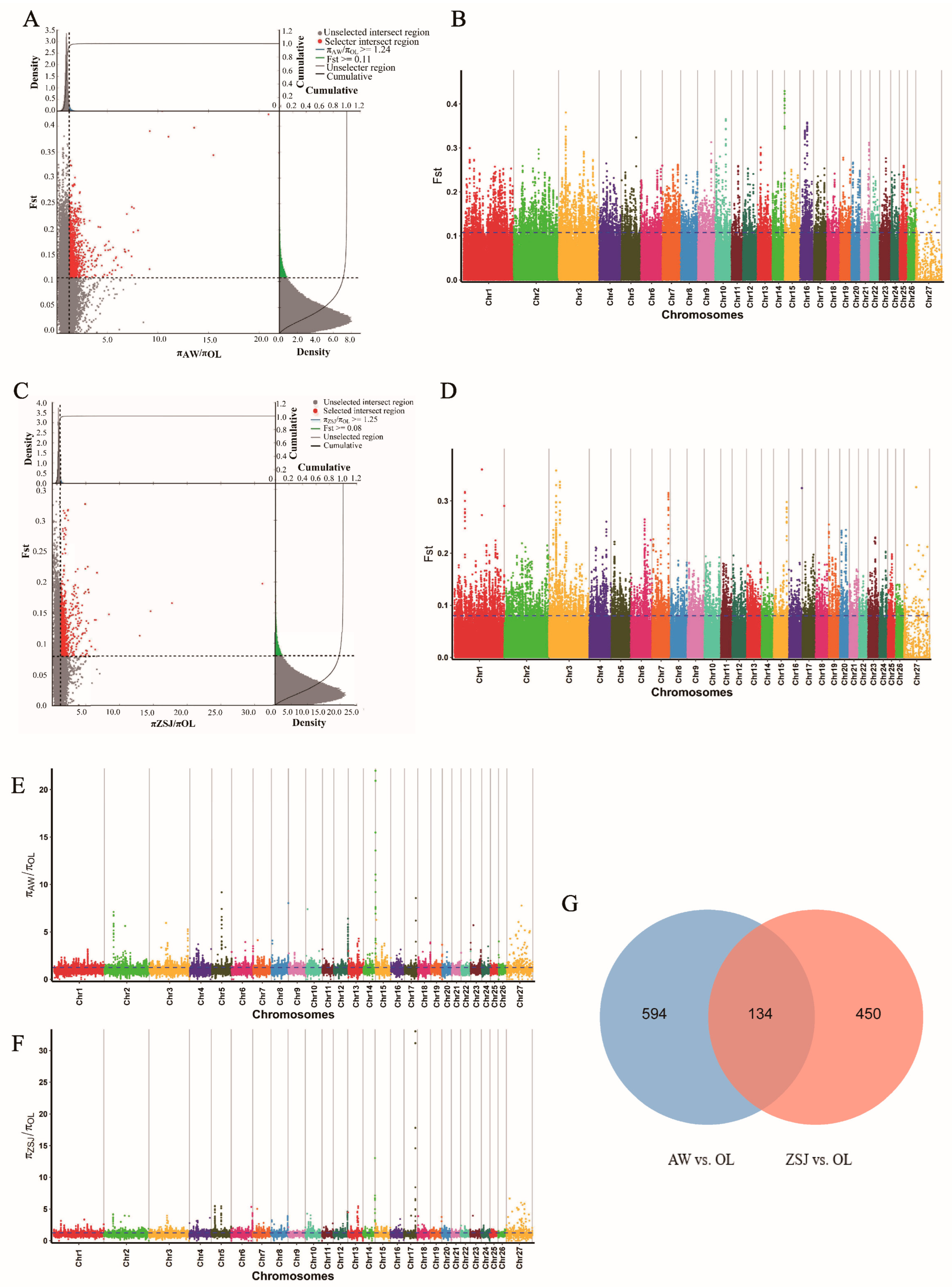
2.4. Selection Signal Analyses
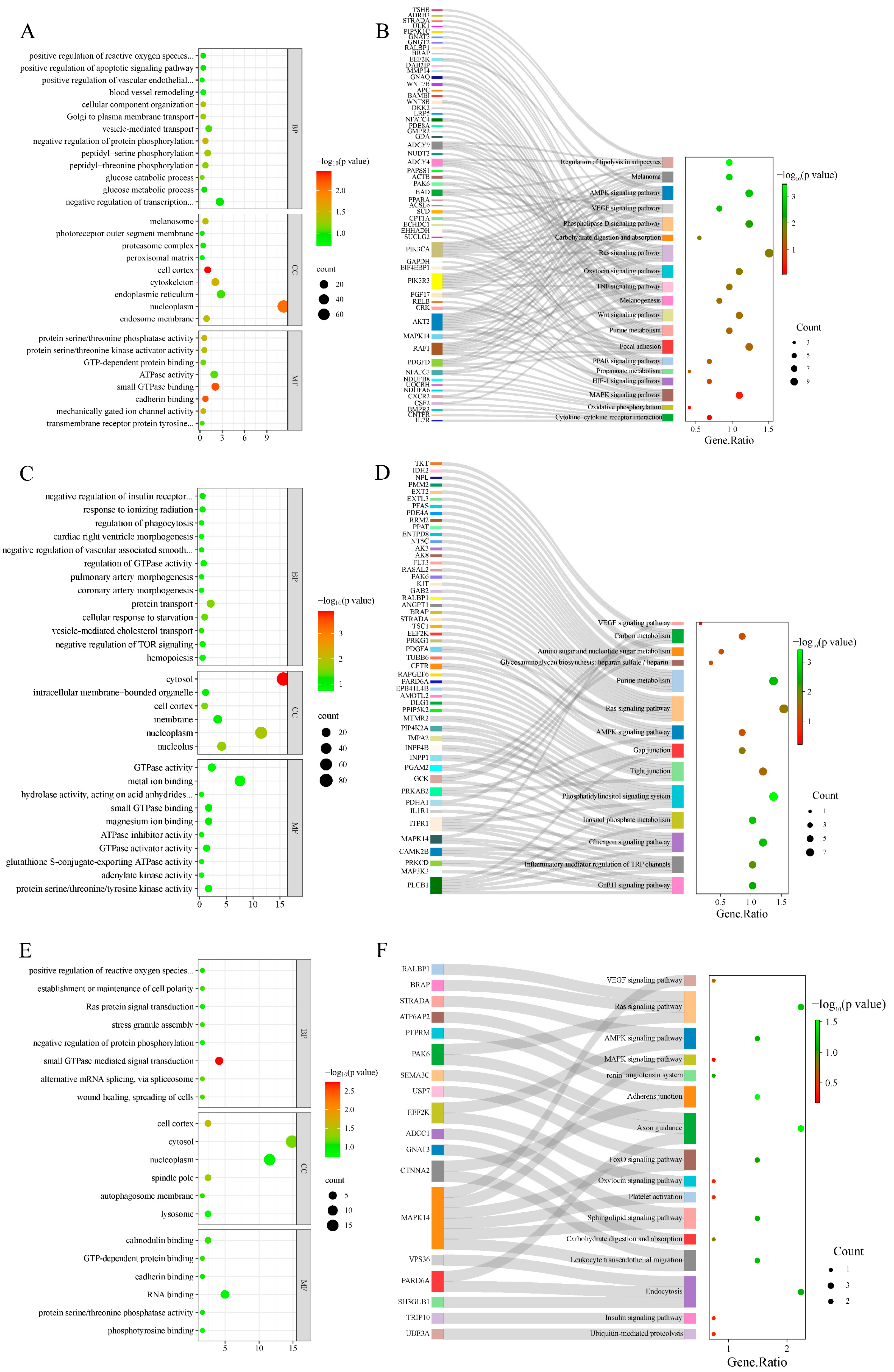
2.5. Candidate Gene Enrichment Analysis
3. Results
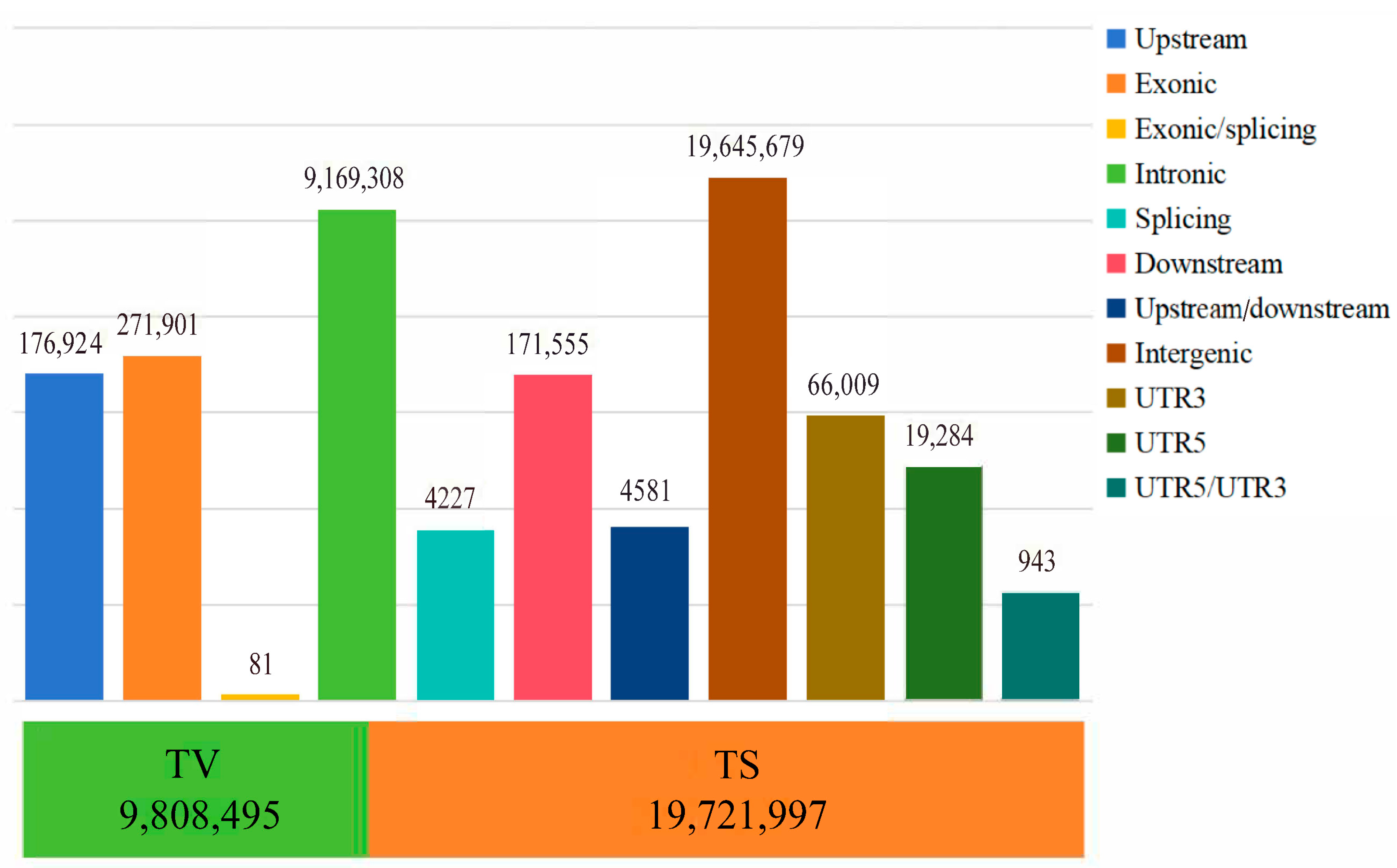
3.1. Genetic Variation and Population Genetic Analysis
3.2. Analysis of Selection Signals
3.3. Enrichment Analysis of Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, L.G.; Yao, T.; Mosley-Thompson, E.; Davis, M.E.; Henderson, K.A.; Lin, P.N. A high-resolution millennial record of the south asian monsoon from himalayan ice cores. Science 2000, 289, 1916–1919. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Feng, J.; Li, L.; Zhong, T.; Wang, L.J.; Guo, J.Z.; Ba, G.; Song, T.Z.; Zhang, H.P. Polymorphisms, differentiation, and phylogeny of 10 tibetan goat populations inferred from mitochondrial d-loop sequences. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Bakoev, S.Y.; Korobeinikova, A.V.; Mishina, A.I.; Kabieva, S.S.; Mitrofanov, S.I.; Ivashechkin, A.A.; Akinshina, A.I.; Snigir, E.A.; Yudin, S.M.; Yudin, V.S.; et al. Genomic signatures of positive selection in human populations of the oxt, oxtr, avp, avpr1a and avr1b gene variants related to the regulation of psychoemotional response. Genes 2023, 14, 2053. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z. Unveiling recent and ongoing adaptive selection in human populations. PLoS Biol. 2024, 22, e3002469. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Zhong, X.; Junjing, W.; Yu, Z.; Mu, Q.; Jiawei, Z.; Yue, F.; Zipeng, L.; Hua, S.; Ruiyi, L.; Zhongxu, S.; et al. Genome-wide detection of selection signatures in jianli pigs reveals novel cis-regulatory haplotype in ednrb associated with two-end black coat color. BMC Genom. 2024, 25, 23. [Google Scholar]
- Jing, X.P.; Wang, W.J.; Degen, A.A.; Guo, Y.M.; Kang, J.P.; Liu, P.P.; Ding, L.M.; Shang, Z.H.; Zhou, J.W.; Long, R.J. Small intestinal morphology and sugar transporters expression when consuming diets of different energy levels: Comparison between Tibetan and small-tailed Han sheep. Animal 2022, 16, 100463. [Google Scholar] [CrossRef]
- Liu, J.; Yuan, C.; Guo, T.; Wang, F.; Zeng, Y.; Ding, X.; Lu, Z.; Renqing, D.; Zhang, H.; Xu, X.; et al. Genetic signatures of high-altitude adaptation and geographic distribution in Tibetan sheep. Sci. Rep. 2020, 10, 18332. [Google Scholar] [CrossRef]
- Zhang, Q.; Que, M.; Li, W.; Gao, S.; Tan, X.; Bu, D. Gangba sheep in the Tibetan plateau: Validating their unique meat quality and grazing factor analysis. J. Environ. Sci. 2021, 101, 117–122. [Google Scholar] [CrossRef]
- Liu, G.; Liu, R.; Li, Q.; Tang, X.; Yu, M.; Li, X.; Cao, J.; Zhao, S. Identification of microRNAs in wool follicles during anagen, catagen, and telogen phases in Tibetan sheep. PLoS ONE 2013, 8, e77801. [Google Scholar] [CrossRef]
- Zhu, K.; Ge, D.; Wen, Z.; Xia, L.; Yang, Q. Evolutionary Genetics of Hypoxia and Cold Tolerance in Mammals. J. Mol. Evol. 2018, 86, 618–634. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Han, B.; Li, X.; Liu, D.; Zhou, B.; Zhao, C.; Zhang, N.; Wang, L.; Pei, Q.; Zhao, K. Genetic diversity and selection of Tibetan sheep breeds revealed by whole-genome resequencing. Anim. Biosci. 2023, 36, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lu, J.; Fei, X.; Lu, Z.; Quan, K.; Liu, Y.; Chu, M.; Di, R.; Wei, C.; Wang, H. Selection signatures analysis reveals genes associated with high-altitude adaptation in tibetan goats from nagqu, tibet. Animals 2020, 10, 1599. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Yang, J.; Xie, X.L.; Lv, F.H.; Cao, Y.H.; Li, W.R.; Liu, M.J.; Wang, Y.T.; Li, J.Q.; Liu, Y.G.; et al. The genome landscape of tibetan sheep reveals adaptive introgression from argali and the history of early human settlements on the qinghai-tibetan plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, W.R.; Lv, F.H.; He, S.G.; Tian, S.L.; Peng, W.F.; Sun, Y.W.; Zhao, Y.X.; Tu, X.L.; Zhang, M.; et al. Whole-genome sequencing of native sheep provides insights into rapid adaptations to extreme environments. Mol. Biol. Evol. 2016, 33, 2576–2592. [Google Scholar] [CrossRef]
- Nosrati, M.; Asadollahpour Nanaei, H.; Amiri Ghanatsaman, Z.; Esmailizadeh, A. Whole genome sequence analysis to detect signatures of positive selection for high fecundity in sheep. Reprod. Domest. Anim. 2018, 54, 358–364. [Google Scholar] [CrossRef]
- Shi, H.; Li, T.; Su, M.; Wang, H.; Li, Q.; Lang, X.; Ma, Y. Whole genome sequencing revealed genetic diversity, population structure, and selective signature of panou tibetan sheep. BMC Genom. 2023, 24, 50. [Google Scholar] [CrossRef]
- Guo, J.; Tao, H.; Li, P.; Li, L.; Zhong, T.; Wang, L.; Ma, J.; Chen, X.; Song, T.; Zhang, H. Whole-genome sequencing reveals selection signatures associated with important traits in six goat breeds. Sci. Rep. 2018, 8, 10405. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.; Zhou, G.; Guo, J.; Yan, H.; Niu, Y.; Li, Y.; Yuan, C.; Geng, R.; Lan, X.; et al. Whole-genome sequencing of eight goat populations for the detection of selection signatures underlying production and adaptive traits. Sci. Rep. 2016, 6, 38932. [Google Scholar] [CrossRef]
- Ge, R.L.; Cai, Q.; Shen, Y.Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.; Huang, Y.; et al. Draft genome sequence of the tibetan antelope. Nat. Commun. 2013, 4, 1858. [Google Scholar] [CrossRef]
- Wei, C.; Wang, H.; Liu, G.; Zhao, F.; Kijas, J.W.; Ma, Y.; Lu, J.; Zhang, L.; Cao, J.; Wu, M.; et al. Genome-wide analysis reveals adaptation to high altitudes in tibetan sheep. Sci. Rep. 2016, 6, 26770. [Google Scholar] [CrossRef] [PubMed]
- Heng, L.; Richard, D. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar]
- Heng, L.; Bob, H.; Alec, W.; Tim, F.; Jue, R.; Nils, H.; Gabor, M.; Goncalo, A.; Richard, D. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar]
- Quinlan, A.R.; Hall, I.M. Bedtools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Vilella, A.J.; Severin, J.; Ureta-Vidal, A.; Heng, L.; Durbin, R.; Birney, E. Ensemblcompara genetrees: Complete, duplication-aware phylogenetic trees in vertebrates. Genome. Res. 2009, 19, 327–335. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (itol) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome. Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T. The variant call format and vcftools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating f-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Wright, S. Genetical structure of populations. Nature 1950, 166, 247–249. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, B.; Wittelsbürger, U.; Ramos-Onsins, S.E.; Lercher, M.J. Popgenome: An efficient swiss army knife for population genomic analyses in R. Mol. Biol. Evol. 2014, 31, 1929–1936. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Mingyao, L.; Hakon, H. Annovar: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar]
- Ding, X.Z.; Liang, C.N.; Guo, X.; Wu, X.Y.; Wang, H.B.; Johnson, K.A.; Yan, P. Physiological insight into the high-altitude adaptations in domesticated yaks (Bos grunniens) along the qinghai-tibetan plateau altitudinal gradient. Livest. Sci. 2014, 162, 233–239. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, M.; Li, T.; Lu, Z.; Wang, H.; Yuan, Z.; Wei, C. Whole-genome resequencing reveals selection signal related to sheep wool fineness. Animals 2023, 13, 2944. [Google Scholar] [CrossRef]
- Li, X.; Yuan, L.; Wang, W.; Zhang, D.; Zhao, Y.; Chen, J.; Xu, D.; Zhao, L.; Li, F.; Zhang, X. Whole genome re-sequencing reveals artificial and natural selection for milk traits in East Friesian sheep. Front. Vet. Sci. 2022, 9, 1034211. [Google Scholar] [CrossRef]
- Liu, L.L.; Meng, J.; Ma, H.Y.; Cao, H.; Liu, W.J. Candidate genes for litter size in xinjiang sheep identified by specific locus amplified fragment (slaf) sequencing. Anim. Biotechnol. 2023, 34, 3053–3062. [Google Scholar] [CrossRef]
- Bigham, A.W.; Lee, F.S. Human high-altitude adaptation: Forward genetics meets the hif pathway. Genes. Dev. 2014, 28, 2189–2204. [Google Scholar] [CrossRef]
- Minet, E.; Arnould, T.; Michel, G.; Roland, I.; Mottet, D.; Raes, M.; Remacle, J.; Michiels, C. Erk activation upon hypoxia: Involvement in hif-1 activation. FEBS Lett. 2000, 468, 53–58. [Google Scholar] [CrossRef]
- Zhang, B.; Chamba, Y.; Shang, P.; Wang, Z.; Ma, J.; Wang, L.; Zhang, H. Comparative transcriptomic and proteomic analyses provide insights into the key genes involved in high-altitude adaptation in the tibetan pig. Sci. Rep. 2017, 7, 3654. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, W.; Zhang, B.; Ling, Y.; Kim, W.K.; Zhang, H. Comprehensive analysis of coding and non-coding rna transcriptomes related to hypoxic adaptation in tibetan chickens. J. Anim. Sci. Biotechnol. 2021, 12, 1309–1322. [Google Scholar] [CrossRef] [PubMed]
- Siyuan, F.; Jideng, M.; Keren, L.; Jinwei, Z.; Wanling, Q.; Yan, L.; Long, J.; Xun, W.; Anan, J.; Lingyan, L.; et al. Comparative microrna transcriptomes in domestic goats reveal acclimatization to high altitude. Front. Genet. 2020, 11, 809. [Google Scholar]
- Xin, J.W.; Chai, Z.X.; Zhang, C.F.; Yang, Y.M.; Zhang, Q.; Zhu, Y.; Cao, H.W.; Yang Ji, C.; Zhong, J.C. Transcriptome analysis identified long non-coding RNAs involved in the adaption of yak to high-altitude environments. R. Soc. Open Sci. 2020, 7, 200625. [Google Scholar] [CrossRef] [PubMed]
- Xin, C.; Rui, K.; Guido, K.; Daolin, T. Ferroptosis in infection, inflammation, and immunity. J. Exp. Med. 2021, 218, e20210518. [Google Scholar]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Schödel, J.; Grampp, S.; Maher, E.R.; Moch, H.; Ratcliffe, P.J.; Russo, P.; Mole, D.R. Hypoxia, hypoxia-inducible transcription factors, and renal cancer. Eur. Urol. 2016, 69, 646–657. [Google Scholar] [CrossRef]
- Hewitson, K.S.; McNeill, L.A.; Riordan, M.V.; Tian, Y.M.; Bullock, A.N.; Welford, R.W.; Elkins, J.M.; Oldham, N.J.; Bhattacharya, S.; Gleadle, J.M.; et al. Hypoxia-inducible factor (hif) asparagine hydroxylase is identical to factor inhibiting HIF (fih) and is related to the cupin structural family. J. Biol. Chem. 2002, 277, 26351–26355. [Google Scholar] [CrossRef]
- Qu, Y.; Zhao, H.; Han, N.; Zhou, G.; Song, G.; Gao, B.; Tian, S.; Zhang, J.; Zhang, R.; Meng, X.; et al. Ground tit genome reveals avian adaptation to living at high altitudes in the tibetan plateau. Nat. Commun. 2013, 4, 2071. [Google Scholar] [CrossRef]
- Lee, S.H.; Koo, K.H.; Park, J.W.; Kim, H.J.; Ye, S.K.; Park, J.B.; Park, B.K.; Kim, Y.N. Hif-1 is induced via egfr activation and mediates resistance to anoikis-like cell death under lipid rafts/caveolae-disrupting stress. Carcinogenesis 2009, 30, 1997–2004. [Google Scholar] [CrossRef]
- Malec, V.; Gottschald, O.R.; Li, S.; Rose, F.; Seeger, W.; Hänze, J. Hif-1 alpha signaling is augmented during intermittent hypoxia by induction of the nrf2 pathway in nox1-expressing adenocarcinoma A549 cells. Free Radic. Biol. Med. 2010, 48, 1626–1635. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Z.; Zhang, R.; Hafner, M.S.; Wong, H.K.; Jiao, Z.; Chopp, M. Erythropoietin up-regulates socs2 in neuronal progenitor cells derived from svz of adult rat. Neuroreport 2004, 15, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Jiang, Y.Q.; Lu, M.M.; Morrisey, E.E. Wnt7b regulates mesenchymal proliferation and vascular development in the lung. Development 2002, 129, 4831–4842. [Google Scholar] [CrossRef] [PubMed]
- Hotary, K.; Allen, E.; Punturieri, A.; Yana, I.; Weiss, S.J. Regulation of cell invasion and morphogenesis in a three-dimensional type i collagen matrix by membrane-type matrix metalloproteinases 1, 2, and 3. J. Cell Biol. 2000, 149, 1309–1323. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.D. A Study of Effects on Mmp14 Transcriptional Regulation and Angiogenesis by Hypoxia and Statins. Bachelor’s Thesis, Queen Mary University of London, London, UK, 2014. [Google Scholar]
- Leung, T.; Chen, H.; Stauffer, A.M.; Giger, K.E.; Sinha, S.; Horstick, E.J.; Humbert, J.E.; Hansen, C.A.; Robishaw, J.D. Zebrafish G protein gamma2 is required for VEGF signaling during angiogenesis. Blood 2006, 108, 160–166. [Google Scholar] [CrossRef]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef]
- Jia, C.; Kong, X.; Koltes, J.E.; Gou, X.; Yang, S.; Yan, D.; Lu, S.; Wei, Z. Gene co-expression network analysis unraveling transcriptional regulation of high-altitude adaptation of tibetan pig. PLoS ONE 2016, 11, e0168161. [Google Scholar] [CrossRef]
- Tiedke, J.; Gerlach, F.; Mitz, S.A.; Hankeln, T.; Burmester, T. Ontogeny of globin expression in zebrafish (Danio rerio). J. Comp. Physiol. B 2011, 181, 1011–1021. [Google Scholar] [CrossRef]
- Zhuang, Z.; Guo, S.; Ji, Z.P.; Zhuang, M.Y.; Yin, L.L.; Gangping, W.; Cheng, J.; Zhongliang, M.; Tian, J.; Peijian, Z.; et al. Hypoxia preconditioning induced hif-1α promotes glucose metabolism and protects mitochondria in liver I/R injury. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 610–619. [Google Scholar]
- Sharma, N.K.; Sethy, N.K.; Bhargava, K. Comparative proteome analysis reveals differential regulation of glycolytic and antioxidant enzymes in cortex and hippocampus exposed to short-term hypobaric hypoxia. J. Proteomics 2013, 79, 277–298. [Google Scholar] [CrossRef]
- González-Cinca, N.; Pérez de la Ossa, P.; Carreras, J.; Climent, F. Effects of thyroid hormone and hypoxia on 2,3-bisphosphoglycerate, bisphosphoglycerate synthase and phosphoglycerate mutase in rabbit erythroblasts and reticulocytes in vivo. Horm. Res. 2004, 62, 191–196. [Google Scholar] [CrossRef]
- Tang, W.; Arnett, D.K.; Devereux, R.B.; Panagiotou, D.; Province, M.A.; Miller, M.B.; Simone, G.d.; Gu, C.; Ferrell, R.E. Identification of a novel 5–base pair deletion in calcineurin b (ppp3r1) promoter region and its association with left ventricular hypertrophy. Am. Heart J. 2004, 150, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chai, Z.; Hu, D.; Ji, Q.; Xin, J.; Zhang, C.; Zhong, J. A global analysis of cnvs in diverse yak populations using whole-genome resequencing. BMC Genom. 2019, 20, 61. [Google Scholar] [CrossRef] [PubMed]
- Madison, S.; Hannah, D.; Jennifer, T.; Jane, B. Fat deposition and fat effects on meat quality-a review. Animals 2022, 12, 1550. [Google Scholar] [CrossRef] [PubMed]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B.; et al. Genetic evidence for high-altitude adaptation in tibet. Science 2010, 329, 72–75. [Google Scholar] [CrossRef]
- Kennedy, S.L.; Stanley, W.C.; Panchal, A.R.; Mazzeo, R.S. Alterations in enzymes involved in fat metabolism after acute and chronic altitude exposure. J. Appl. Physiol. 2001, 90, 17–22. [Google Scholar] [CrossRef]
- Robinson, E.; Grieve, D.J. Significance of peroxisome proliferator-activated receptors in the cardiovascular system in health and disease. Pharmacol. Ther. 2009, 122, 246–263. [Google Scholar] [CrossRef]
- Holden, J.E.; Stone, C.K.; Clark, C.M.; Brown, W.D.; Nickles, R.J.; Stanley, C.; Hochachka, P.W. Enhanced cardiac metabolism of plasma glucose in high-altitude natives: Adaptation against chronic hypoxia. J. Appl. Physiol. 1995, 79, 222–228. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Abbr. | Sex | Age | Size | Location | Altitude | Longitude and Latitude |
|---|---|---|---|---|---|---|---|
| Oula sheep | OL | Male | Adult | 20 | Oula Township, Maqu County, Gannan Tibetan Autonomous Prefecture, Gansu Province | 3501 m | N: 33°51′312″ E: 101°52′424″ |
| Zashijia sheep | ZSJ | Male | Adult | 20 | Yocai Township, Qumalai County, Yushu Tibetan Autonomous Prefecture, Qinghai Province | 4269 m | N: 34°14′866″ E: 95°80′422″ |
| Awang sheep | AW | Male | Adult | 20 | Awang Township, Gongjue County, Changdu, Xizang Autonomous Region | 4643 m | N: 30°12′101″ E: 98°63′98″ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Yuan, C.; An, X.; Guo, T.; Zhang, W.; Lu, Z.; Liu, J. Genome-Wide Selection Signals Reveal Candidate Genes Associated with Plateau Adaptation in Tibetan Sheep. Animals 2024, 14, 3212. https://doi.org/10.3390/ani14223212
Song Y, Yuan C, An X, Guo T, Zhang W, Lu Z, Liu J. Genome-Wide Selection Signals Reveal Candidate Genes Associated with Plateau Adaptation in Tibetan Sheep. Animals. 2024; 14(22):3212. https://doi.org/10.3390/ani14223212
Chicago/Turabian StyleSong, Yufang, Chao Yuan, Xuejiao An, Tingting Guo, Wentao Zhang, Zengkui Lu, and Jianbin Liu. 2024. "Genome-Wide Selection Signals Reveal Candidate Genes Associated with Plateau Adaptation in Tibetan Sheep" Animals 14, no. 22: 3212. https://doi.org/10.3390/ani14223212
APA StyleSong, Y., Yuan, C., An, X., Guo, T., Zhang, W., Lu, Z., & Liu, J. (2024). Genome-Wide Selection Signals Reveal Candidate Genes Associated with Plateau Adaptation in Tibetan Sheep. Animals, 14(22), 3212. https://doi.org/10.3390/ani14223212