
Dentex maroccanus Valenciennes, 1830 Otolith Morphology, Age, and Growth in the Aegean Sea (E. Mediterranean)

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
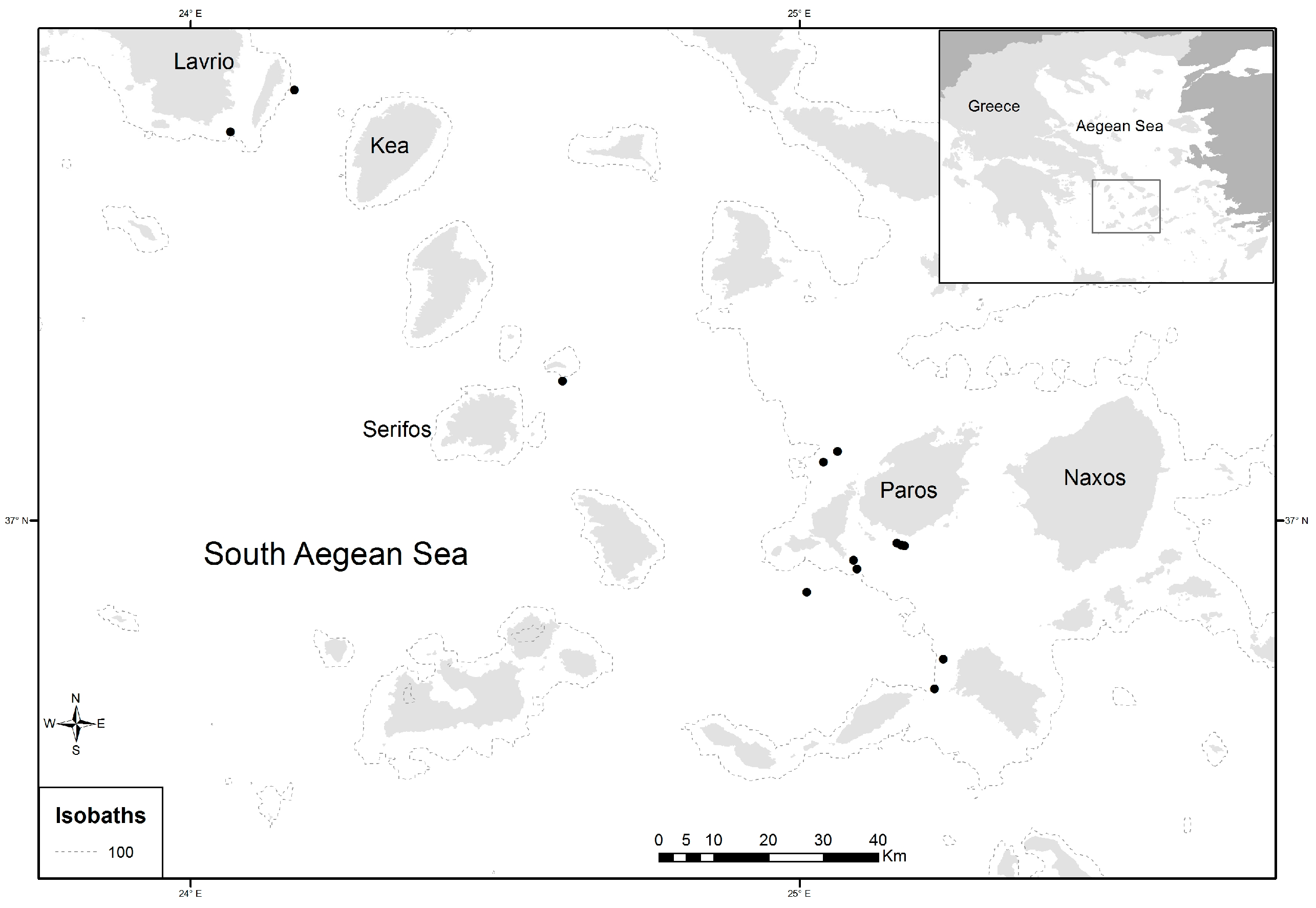
2.1. Study Area and Sampling
2.2. Data Analysis
2.2.1. Length Frequency Distribution
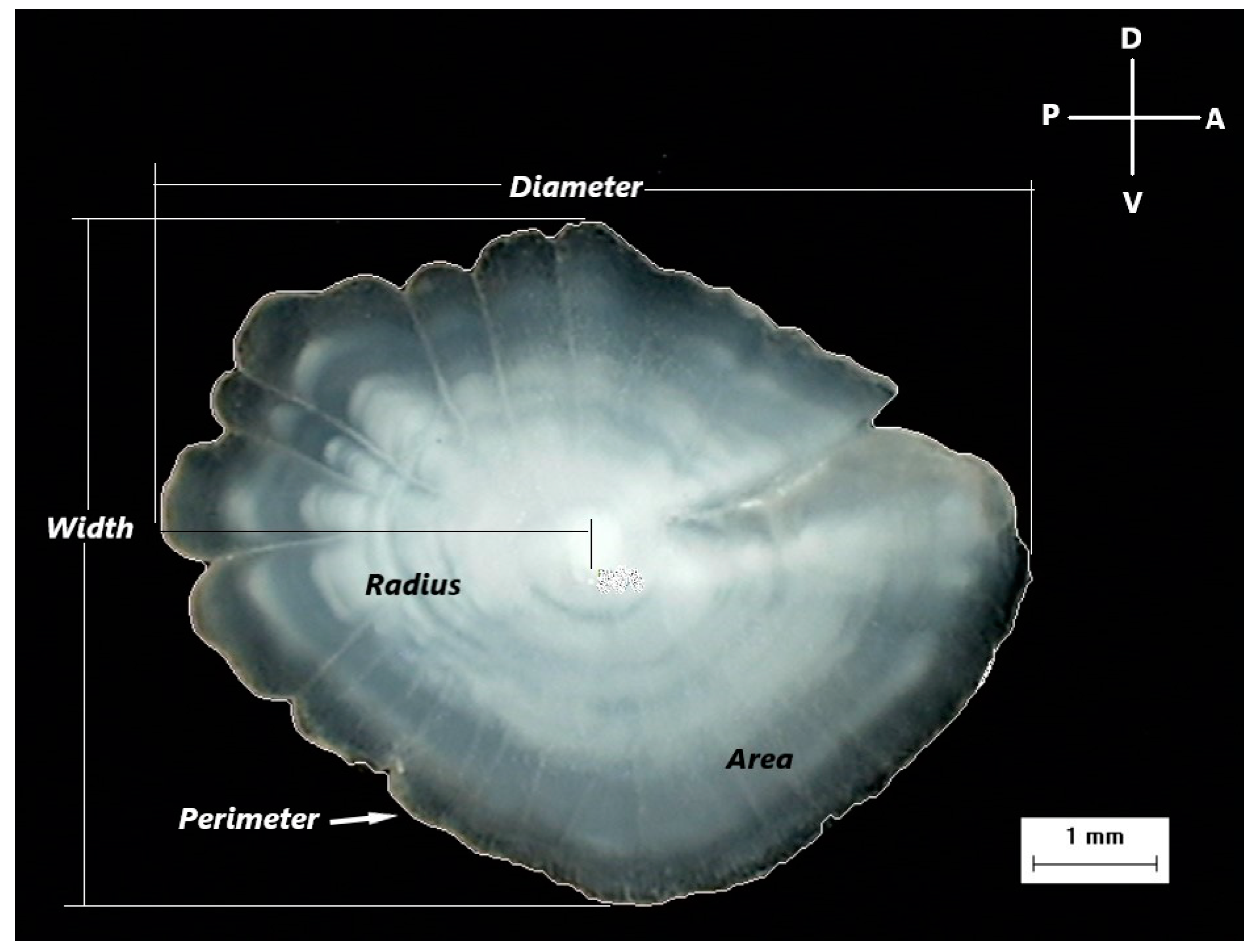
2.2.2. Otolith Morphometrics
2.2.3. Growth and Age
3. Results
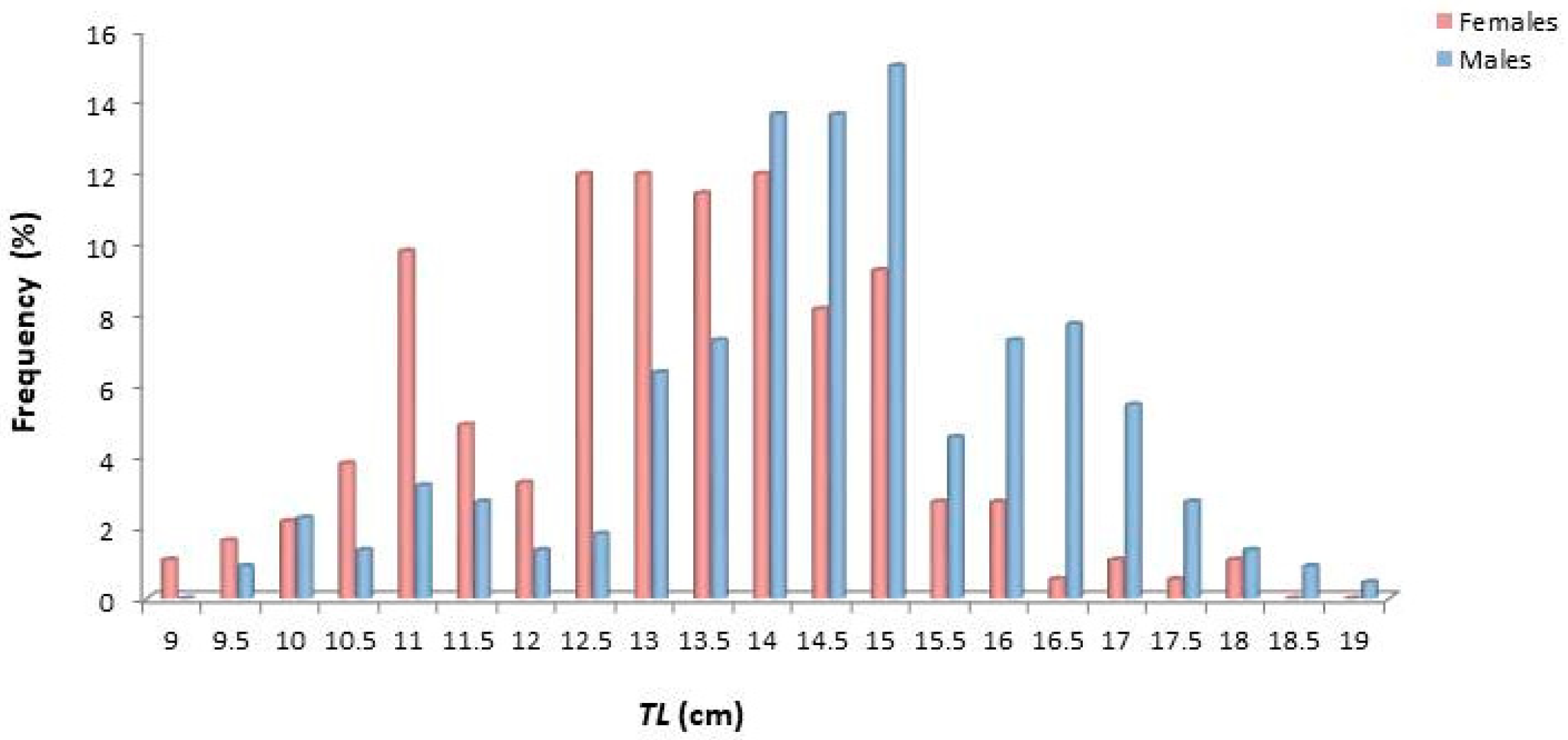
3.1. Length Distribution
3.2. Otolith Morphometrics
3.3. Growth and Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campana, S.E.; Casselman, J.M. Stock discrimination using otolith shape analysis. Can. J. Fish. Aquat. Sci. 1993, 50, 1062–1083. [Google Scholar] [CrossRef]
- Tuset, V.M.; Lombarte, A.; González, J.A.; Pertusa, J.F.; Lorente, M.J. Comparative morphology of the sagittal otolith in Serranus spp. J. Fish. Biol. 2003, 63, 1491–1504. [Google Scholar] [CrossRef]
- Gagliano, M.; McCormick, M.I. Feeding history influences otolith shape in tropical fish. Mar. Ecol. Progr. Ser. 2004, 278, 291–296. [Google Scholar] [CrossRef]
- Morales-Nin, B.; Tores, G.; Lombarte, A.; Recasens, L. Otolith growth and age estimation in the European hake. J. Fish. Biol. 2005, 53, 1155–1168. [Google Scholar] [CrossRef]
- Teimori, A.; Jawad, L.A.J.; Al-Kharusi, L.H.; Al-Mamry, J.M.; Reichenbacher, B. Late Pleistocene to Holocene diversification and historical zoogeography of the Arabian killifish (Aphanius dispar) inferred from otolith morphology. Sci. Mar. 2012, 76, 637–645. [Google Scholar] [CrossRef]
- Vignon, M. Ontogenetic trajectories of otolith shape during shift in habitat use: Interaction between otolith growth and environment. J. Exp. Mar. Biol. Ecol. 2012, 420–421, 26–32. [Google Scholar] [CrossRef]
- Giménez, J.; Manjabacas, A.; Tuset, V.M.; Lombarte, A. Relationships between otolith and fish size from Mediterranean and north-eastern Atlantic species to be used in predator–prey studies. J. Fish. Biol. 2016, 89, 2195–2202. [Google Scholar] [CrossRef]
- Lombarte, A.; Miletić, M.; Kovačić, M.; Otero-Ferrer, J.L.; Tuset, V.M. Identifying sagittal otoliths of Mediterranean Sea gobies: Variability among phylogenetic lineages. J. Fish. Biol. 2018, 92, 1768–1787. [Google Scholar] [CrossRef]
- Biolé, F.G.; Callicó Fortunato, R.; Thompson, G.A.; Volpedo, A.V. Application of otolith morphometry for the study of ontogenetic variations of Odontesthes argentinensis. Environ. Biol. Fishes 2019, 102, 1301–1310. [Google Scholar] [CrossRef]
- Carvalho, B.; Volpedo, A.; Fávaro, L. Ontogenetic and sexual variation in the sagitta otolith of Menticirrhus americanus (Teleostei; Sciaenidae) (Linnaeus, 1758) in a subtropical environment. Pap. Avulsos. Zool. 2020, 60, 1–12. [Google Scholar] [CrossRef]
- Mahé, K.; MacKenzie, K.; Ider, D.; Massaro, A.; Hamed, O.; Jurado-Ruzafa, A.; Gonçalves, P.; Anastasopoulou, A.; Jadaud, A.; Mytilineou, C.; et al. Directional Bilateral Asymmetry in Fish Otolith: A Potential Tool to Evaluate Stock Boundaries? Symmetry 2021, 13, 987. [Google Scholar] [CrossRef]
- Carvalho, B.M.; Oliveira-Freitas, M.; Lapuch, I.; Volpedo, A.V.; Vitule, J.R. Age, growth, and ontogenetic variation in the sagitta otolith of Opsanus beta (Goode & Bean, 1880), a non-native species in a wetland of international importance. Lat. Am. J. Aquat. Res. 2022, 50, 124–134. [Google Scholar]
- Morales, C.J.C.; Barnuevo, K.D.E.; Delloro, E.S., Jr.; Cabebe-Barnuevo, R.A.; Calizo, J.K.S.; Lumayno, S.D.P.; Babaran, R.P. Otolith Morphometric and Shape Distinction of Three Redfin Species under the Genus Decapterus (Teleostei: Carangidae) from Sulu Sea, Philippines. Fishes 2023, 8, 95. [Google Scholar] [CrossRef]
- Campana, S.E. Otolith science entering the 21st century. Mar. Freshw. Res. 2005, 56, 485–496. [Google Scholar] [CrossRef]
- Tuset, V.M.; Lombarte, A.; Assis, C.A. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Sci. Mar. 2008, 72, 7–198. [Google Scholar] [CrossRef]
- Jemaa, S.; Bacha, M.; Khalaf, G.; Dessailly, D.; Rabhi, K.; Amara, R. What can otolith shape analysis tell us about population structure of the European sardine, Sardina pilchardus, from Atlantic and Mediterranean waters? J. Sea Res. 2015, 96, 11–17. [Google Scholar] [CrossRef]
- Echreshavi, S.; Esmaeili, H.R.; Teimori, A.; Safaie, M. Otolith morphology: A hidden tool in the taxonomic study of goatfishes (Teleostei: Perciformes: Mullidae). Zool. Stud. 2021, 60, 36. [Google Scholar] [CrossRef]
- Ferri, J. Otoliths and Their Applications in Fishery Science. Fishes 2023, 8, 35. [Google Scholar] [CrossRef]
- Nikiforidou, V.; Mytilineou, C.; Alexandropoulos, A.; Anastasopoulou, A. Age, Growth, and Otolith Morphometrics of Trachinus draco (L., 1758) and Trachinus radiatus (Cuvier, 1829) in the Eastern Mediterranean. Fishes 2024, 9, 152. [Google Scholar] [CrossRef]
- Volpedo, A.; Echeverría, D.D. Ecomorphological patterns of the sagitta in fish on the continental shelf off Argentine. Fish. Res. 2003, 60, 551–560. [Google Scholar] [CrossRef]
- Cardinale, M.; Doering-Arjes, P.; Kastowsky, M.; Mosegaard, H. Effects of sex, stock, and environment on the shape of known-age Atlantic cod (Gadus morhua) otoliths. Can. J. Fish. Aquat. Sci. 2004, 61, 158–167. [Google Scholar] [CrossRef]
- Lombarte, A.; Palmer, M.; Matallanas, J.; Gómez-Zurita, J.; Morales-Nin, B. Ecomorphological trends and phylogenetic inertia of otolith sagittae in Nototheniidae. Environ. Biol. Fishes 2010, 89, 607–618. [Google Scholar] [CrossRef]
- Vignon, M.; Morat, F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish. Mar. Ecol. Prog. Ser. 2010, 411, 231–241. [Google Scholar] [CrossRef]
- He, T.; Chen, C.; Qin, J.; Li, Y.; Wu, R.; Gao, T. The use of otolith shape to identify stocks of redlip mullet, Liza haematocheilus. Pak. J. Zool. 2020, 52, 2265–2273. [Google Scholar] [CrossRef]
- Wang, C.; Hu, L.; Song, Y.; Xie, H.; Yang, L.; Serekbol, G.; Huo, B.; Chen, S. The Evolution of Three Schizothoracinae Species from Two Major River Systems in Northwest China Based on Otolith Morphology and Skeletal Structure. Biology 2024, 13, 517. [Google Scholar] [CrossRef]
- Bolles, K.L.; Begg, G.A. Distinctions between Silver Hake (Merluccius bilinearis) Stocks in U.S. Waters of the Northwest Atlantic Based on Whole Otolith Morphometric. Fish. Bull. 2000, 98, 451–462. [Google Scholar]
- Mahe, K.; Gourtay, C.; Defruit, G.B.; Chantre, C.; de Pontual, H.; Amara, R.; Claireaux, G.; Audet, C.; Zarnbonino-Infante, J.L.; Ernande, B. Do environmental conditions (temperature and food composition) affect otolith shape during fish early juvenile phase? An experimental approach applied to European Seabass (Dicentrarchus labrax). J. Exp. Mar. Biol. Ecol. 2019, 521, 151239. [Google Scholar] [CrossRef]
- Tuset, V.M.; Imondi, R.; Aguado, G.; Otero-Ferrer, J.L.; Santschi, L.; Lombarte, A.; Love, M. Otolith patterns of rockfishes from the northeastern Pacific. J. Morphol. 2015, 276, 458–469. [Google Scholar] [CrossRef]
- Tuset, V.M.; Otero-Ferrer, J.L.; Stransky, C.; Imondi, R.; Orlov, A.; Zhenjiang, Y.; Venerus, L.; Santschi, L.; Afanasiev, P.; Zhuang, L.O.; et al. Otolith shape lends support to the sensory drive hypothesis in rockfishes. J. Evol. Biol. 2016, 29, 2083–2097. [Google Scholar] [CrossRef]
- Parmentier, E.; Boistel, R.; Bahri, M.A.; Plenevaux, A.; Schwarzhans, W. Sexual dimorphism in the sonic system and otolith morphology of Neobythites gilli (Ophidiiformes). J. Zool. 2018, 305, 274–280. [Google Scholar] [CrossRef]
- Vaux, F.; Rasmuson, L.K.; Kautzi, L.A.; Rankin, P.S.; Blume, M.T.O.; Lawrence, K.A.; Bohn, S.; O’Malley, K.G. Sex matters: Otolith shape and genomic variation in deacon rockfish (Sebastes diaconus). Ecol. Evol. 2019, 9, 13153–13173. [Google Scholar] [CrossRef] [PubMed]
- Başusta, N.; Dürrani, Ö. Sexual dimorphism in the otolith shape of shi drum, Umbrina cirrosa (L.), in the eastern Mediterranean Sea: Fish size–otolith size relationships. J. Fish. Biol. 2021, 99, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Bauchot, M.L.; Hureau, J.C. Sparidae. In Fishes of the North-Eastern Atlantic and the Mediterranean (FNAM); Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volumr II, pp. 883–907. [Google Scholar]
- Golani, D.; Öztürk, B.; Başusta, N. Fishes of the Eastern Mediterranean; Publication no. 24; Turkish Marine Research Foundation: Istanbul, Turkey, 2006; 259p. [Google Scholar]
- Grech, D.; Asciutto, E.; Bakiu, R.; Battaglia, P.; Ben-Grira, C.; Öznur, Y.Ç.; Cappuccinelli, R.; Carmona, L.; Chebaane, S.; Crocetta, F.; et al. New records of rarely reported species in the Mediterranean Sea (July 2023). Mediterr. Mar. Sci. 2023, 24, 392–418. [Google Scholar] [CrossRef]
- Hemida, F.; Ghazli, R. Contribution à l’ecologie de Dentex maroccanus (Valenciennes, 1830) des cotes Algeriennes: Repartition geographique, bathymetrique et en fonction du substrat. Rapp. Comm. Int. L’exploration Mer Mediterr. (CIESM) Marseille Fr. 1998, 35, 448–449. [Google Scholar]
- Mina, A.; Mytilineou, C.; Kaminas, A.; Rekleiti, A.; Siapatis, A.; Anastasopoulou, A. Feeding Habits of Dentex maroccanus and the Effect of Body Size. Animals 2023, 13, 939. [Google Scholar] [CrossRef]
- Maravelias, C.D.; Tsitsika, E.V.; Papaconstantinou, C. Evidence of Morocco dentex (Dentex maroccanus) distribution in the NE Mediterranean and relationships with environmental factors determined by Generalized Additive Modelling. Fish. Oceanogr. 2007, 16, 294–302. [Google Scholar] [CrossRef]
- Asimakopoulos, C.; Mytilineou, C. Distribution pattern of Dentex maroccanus in relation to environmental factors in the Aegean Sea. In Proceedings of the 4th International Congress on Applied Ichthyology, Oceanography & Aquatic Environment, Virtual, 4–6 November 2021. [Google Scholar]
- Chemmam-Abdelkader, B.; Kraiem, M.M.; El Abed, A. Période de ponte, sex-ratio et maturité sexuelle de Dentex maroccanus (Teleostei, Sparidae) des côtes tunisiennes. Bull. Inst. Natn. Scien. Tech. Mer. Salammbô 2002, 29, 5–10. [Google Scholar]
- Bayhan, B.; Sever, T.M.; Heral, O. Diet composition of the Morocco dentex: Dentex maroccanus Valenciennes, 1830 (Teleostei: Sparidae) in the central Turkish Aegean Sea. Oceanol. Hydrobiol. Stud. 2017, 46, 133–139. [Google Scholar] [CrossRef]
- Mohdeb, R.; Derbal, F.; Kara, M.H. Diet composition and variations of Morocco dentex Dentex maroccanus (Sparidae) from eastern Algeria. Cybium 2017, 41, 317–324. [Google Scholar]
- Taylan, B.; Bayhan, B.; Heral, O. Fecundity of Morocco Dentex Dentex maroccanus Valenciennes, 1830 distributed in Izmir Bay (Central Aegean Sea of Turkey). TURJAF 2018, 6, 624–627. [Google Scholar] [CrossRef]
- Stromplou, D.; Mina, A.; Pappou, G.; Anastasopoulou, A.; Mytilineou, C. Morphometric characteristics of Dentex maroccanus in the Aegean Sea. In Proceedings of the 4th International Congress on Applied Ichthyology, Oceanography & Aquatic Environment, Virtual, 4–6 November 2021. [Google Scholar]
- Kontaş Yalçınkaya, S. Some Population Parameters of Morocco dentex, Dentex maroccanus Valenciennes, 1830 in the Northeastern Mediterranean Sea. Cumhur. Sci. J. 2023, 44, 276–282. [Google Scholar] [CrossRef]
- Chemmam-Abdelkader, B.; Kraiem, M.M.; El Abed, A. Etude de l’age et de la croissance de deux especes de dentes (Dentex dentex et Dentex maroccanus) des cotes Tunisiennes. Bull. Inst. Natn. Scien. Tech. Mer Salammbô 2004, 31, 43–51. [Google Scholar]
- Mohdeb, R.; Kara, M.H. Age, growth and reproduction of Morocco dentex Dentex maroccanus (Valenciennes, 1830) in Eastern coasts of Algeria. J. Mar. Biol. Ass. UK 2014, 95, 1261–1270. [Google Scholar] [CrossRef]
- Gül, G.; İşmen, A.; Arslan, M. Age, Growth, and Reproduction of Dentex maroccanus (Actinopterygii: Perciformes: Sparidae) in the Saros Bay (North Aegean Sea). Acta Ichthyo. Piscat. 2014, 44, 295–300. [Google Scholar] [CrossRef]
- Heral, O.; Bayhan, B. Age and Growth of Morocco Dentex Dentex maroccanus Valenciennes, 1830 (Actinopterygii: Sparidae) in Izmir Bay, Central Aegean Sea, Turkey. Acta Zool. Bulg. 2020, 72, 149–154. [Google Scholar]
- Ponton, D. Is geometric morphometrics efficient for comparing otolith shape of different fish species? J. Morphol. 2006, 267, 750–757. [Google Scholar] [CrossRef]
- Aguera, A.; Brophy, D. Use of saggital otolith shape analysis to discriminate Northeast Atlantic and Western Mediterranean stocks of Atlantic saury, Scomberesox saurus saurus (Walbaum). Fish. Res. 2011, 110, 465–471. [Google Scholar] [CrossRef]
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish. Biol. 1995, 46, 202–220. [Google Scholar] [CrossRef]
- Pauly, D.; Munro, J.L. Once More on the Comparison of Growth in Fish and Invertebrates. ICLARM Fishbyte 1984, 2, 21. [Google Scholar]
- Bhattacharya, C.G. A simple method of resolution of a distribution into Gaussian components. Biometrics 1967, 23, 115–135. [Google Scholar] [CrossRef]
- Gayanilo, F.; Sparre, P.; Pauly, D. FAO-ICLARM Stock Assessment Tool II (FiSAT II) Revised version—User’s Guide. FAO Comput. Inf. Ser. 2005, 8, 1–168. [Google Scholar]
- Loc, N.X.; Wojciechowski, J. Comparative biology of fish from genus Dentex (Sparidae) of north-west African coast. Acta Ichthyol. Piscat. 1972, 2, 77–79. [Google Scholar]
- Lamrini, A.; Bouymajjane, A. Biologie de Dentex maroccanus (Valenciennes, 1830) dans la region de Safi. Actes L’institut Agron. Vet. Hassan II Rabat (Maroc) 2002, 22, 11–18. [Google Scholar]
- Hussy, K. Otolith shape in juvenile cod (Gadus morhua): Ontogenetic and environmental effects. J. Exp. Mar. Biol. Ecol. 2008, 364, 35–41. [Google Scholar] [CrossRef]
- Volpedo, A.; Thompson, G. Diferencias en el crecimiento de las sagittae de Prionotus nudigula Ginsburg, 1950 (Piscis: Triglidae) en relación al sexo. Boletín Inst. Espańol Oceanogr. 1996, 12, 3–16. [Google Scholar]
- Bose, A.P.H.; Adragna, J.B.; Balshine, S. Otolith morphology varies between populations, sexes and male alternative reproductive tactics in a vocal toadfish Porichthys notatus. J. Fish. Biol. 2017, 90, 311–325. [Google Scholar] [CrossRef]
- Mahé, K.; Evano, H.; Mille, T.; Muths, D.; Bourjea, J. Otolith shape as a valuable tool to evaluate the stock structure of swordfish Xiphias gladius in the Indian Ocean. Afr. J. Mar. Sci. 2016, 38, 457–464. [Google Scholar] [CrossRef]
- Kéver, L.; Colleye, O.; Herrel, A.; Romans, P.; Parmentier, E. Hearing capacities and otolith size in two ophidiiform species (Ophidion rochei and Carapus acus). J. Exp. Biol. 2014, 217, 2517–2525. [Google Scholar]
- Lombarte, A.; Cruz, A. Otolith size trends in marine fish communities from different depth strata. J. Fish. Biol. 2007, 71, 53–76. [Google Scholar] [CrossRef]
- Popper, A.N.; Ramcharitar, J.; Campana, S.E. Why otoliths? Insights from inner ear physiology and fisheries biology. Mar. Freshw. Res. 2005, 56, 497–504. [Google Scholar] [CrossRef]
- Mille, T.; Mahé, K.; Cachera, M.; Villanueva, M.C.; De Pontual, H.; Ernande, B. Diet is correlated with otolith shape in marine fish. Mar. Ecol. Prog. Ser. 2016, 555, 167–184. [Google Scholar] [CrossRef]
- Zengin, M.; Saygin, S.; Polat, N. Relationships between otolith size and total length of bluefish, Pomatomus saltatrix (Linnaeus, 1766) in Black Sea (Turkey). North-West J. Zool. 2017, 13, 169–171. [Google Scholar]
- Yedier, S. Otolith shape analysis and relationships between total length and otolith dimensions of European barracuda, Sphyraena sphyraena in the Mediterranean Sea. Iran. J. Fish. Sci. 2021, 20, 1080–1096. [Google Scholar] [CrossRef]
- Tuset, V.M.; Otero-Ferrer, J.L.; Siliprandi, C.; Manjabacas, A.; Marti-Puig, P.; Lombarte, A. Paradox of otolith shape indices: Routine but overestimated use. Can. J. Fish. Aquat. Sci. 2021, 78, 681–692. [Google Scholar] [CrossRef]
- López-Pérez, C.; Olivar, M.P.; Hulley, P.A.; Tuset, V.M. Length–weight relationships of mesopelagic fishes from the equatorial and tropical Atlantic waters: Influence of environment and body shape. J. Fish. Biol. 2020, 96, 1388–1398. [Google Scholar] [CrossRef]
- Evagelopoulos, A.; Batjakas, I.; Koutsoubas, D. Length–weight relationships of 9 commercial fish species from the North Aegean Sea. Acta Adriat. 2017, 58, 187–192. [Google Scholar] [CrossRef]
- Froese. Cube law, condition factor and weight–length relationships: History, metaanalysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Females | Males | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Relation | a | b | r | R2 | p-Value | a | b | r | R2 | p-Value | ANCOVA p-Value for b |
| D-TL | 0.08 | 0.89 | 0.94 | 0.89 | <0.05 * | 0.07 | 0.92 | 0.95 | 0.91 | <0.05 * | 0.34 |
| W-TL | 0.05 | 0.93 | 0.96 | 0.92 | <0.05 * | 0.03 | 1.01 | 0.95 | 0.90 | <0.05 * | 0.02 |
| R-TL | 0.04 | 0.89 | 0.94 | 0.89 | <0.05 * | 0.04 | 0.93 | 0.95 | 0.91 | <0.05 * | 0.22 |
| A-TL | 0.003 | 1.83 | 0.96 | 0.92 | <0.05 * | 0.002 | 1.95 | 0.95 | 0.91 | <0.05 * | 0.08 |
| P-TL | 0.30 | 0.84 | 0.94 | 0.89 | <0.05 * | 0.21 | 0.91 | 0.94 | 0.88 | <0.05 * | 0.06 |
| Rn-TL | 2.27 | −0.13 | −0.43 | 0.19 | <0.05 * | 2.16 | −0.12 | −0.36 | 0.14 | <0.05 * | 0.75 |
| C-TL | 28.57 | −0.13 | −0.43 | 0.19 | <0.05 * | 27.18 | −0.12 | −0.36 | 0.13 | <0.05 * | 0.75 |
| Rc-TL | 0.71 | 0.02 | 0.13 | 0.02 | >0.05 | 0.76 | 0.007 | 0.06 | 0.003 | >0.05 | - |
| FF-TL | 0.44 | 0.13 | 0.43 | 0.19 | <0.05 * | 0.47 | 0.12 | 0.36 | 0.13 | <0.05 * | 0.75 |
| E-TL | 0.19 | −0.03 | −0.04 | 0.002 | >0.05 | 0.62 | −0.27 | −0.36 | 0.13 | <0.05 * | - |
| Otolith Parameter | Mean ± SE | Common b of Exponential Regression | |
|---|---|---|---|
| Females | Males | ||
| D | 6.551 ± 0.069 (6.667 ± 0.024) | 6.690 ± 0.070 (6.587 ± 0.022) | 0.8995 |
| W | 4.661 ± 0.051 | 4.797 ± 0.055 | - |
| R | 3.394 ± 0.036 (3.455 ± 0.013) | 3.480 ± 0.037 (3.426 ± 0.012) | 0.9054 |
| A | 24.347 ± 0.503 (24.834 ± 0.163) | 25.557 ± 0.535 (24.347 ± 0.170) | 1.8798 |
| P | 18.834 ± 0.191 (19.167 ± 0.067) | 19.296 ± 0.205 (19.006 ± 0.075) | 0.8721 |
| Rn | 1.180 ± 0.004 (1.175 ± 0.004) | 1.182 ± 0.005 (1.183 ± 0.004) | −0.1254 |
| C | 14.825 ± 0.054 (14.771 ± 0.049) | 14.852 ± 0.057 (14.867 ± 0.053) | −0.1253 |
| Rc | 0.785 ± 0.001 | 0.783 ± 0.001 | - |
| FF | 0.849 ± 0.003 (0.852 ± 0.003) | 0.847 ± 0.003 (0.847 ± 0.003) | 0.1253 |
| E | 0.169 ± 0.001 | 0.168 ± 0.001 | - |
| Otolith Parameter | Type III SS | df | MS | F | p-Value |
|---|---|---|---|---|---|
| D | 0.40 | 1 | 0.40 | 5.56 | 0.019 * |
| LOG R | 0.006 | 1 | 0.006 | 3.92 | 0.049 * |
| LOG A | 0.05 | 1 | 0.05 | 8.06 | 0.005 * |
| LOG P | 0.008 | 1 | 0.008 | 4.62 | 0.032 * |
| 1/Rn | 0.001 | 1 | 0.0009 | 0.75 | 0.387 |
| 1/C | 0.00001 | 1 | 0.00001 | 0.74 | 0.391 |
| Rc | 0.0005 | 1 | 0.0005 | 2.03 | 0.156 |
| SQRT FF | 0.0003 | 1 | 0.0003 | 0.75 | 0.386 |
| E | 0.0001 | 1 | 0.0001 | 0.49 | 0.487 |
| Sex | TL (cm) | TW (gr) | a | b | r | R2 | p-Value |
| Females | 9.2–18.1 | 13–95 | 0.009 | 3.20 | 0.97 | 0.95 | <0.05 |
| Males | 9.6–19.2 | 13–99.9 | 0.013 | 3.07 | 0.97 | 0.95 | <0.05 |
| Sex | TL (cm) | EW (gr) | a | b | r | R2 | p-Value |
| Females | 9.2–18.1 | 12.3–87.4 | 0.011 | 3.11 | 0.97 | 0.94 | <0.05 |
| Males | 9.6–19.2 | 12.1–96.5 | 0.012 | 3.08 | 0.98 | 0.95 | <0.05 |
| Age (years) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Females | Males | ||||||||
| Length Interval (cm) | 0+ | I | II | III | 0+ | I | II | III | IV |
| 9.0–9.9 | 5 | 2 | |||||||
| 10.0–10.9 | 11 | 9 | |||||||
| 11.0–11.9 | 6 | 21 | 5 | 9 | |||||
| 12.0–12.9 | 29 | 7 | |||||||
| 13.0–13.9 | 40 | 3 | 30 | ||||||
| 14.0–14.9 | 18 | 19 | 27 | 33 | |||||
| 15.0–15.9 | 22 | 41 | |||||||
| 16.0–16.9 | 2 | 4 | 22 | 11 | |||||
| 17.0–17.9 | 3 | 18 | |||||||
| 18.0–18.9 | 2 | 3 | 2 | ||||||
| 19.0–19.9 | 1 | ||||||||
| N | 22 | 108 | 46 | 9 | 16 | 73 | 96 | 32 | 3 |
| % | 11.89 | 58.37 | 24.87 | 4.87 | 7.27 | 33.18 | 43.64 | 14.55 | 1.36 |
| MTL | 10.41 | 12.94 | 14.92 | 17.06 | 10.53 | 13.43 | 15.31 | 17.21 | 18.73 |
| SE | 0.12 | 0.09 | 0.08 | 0.27 | 0.14 | 0.1 | 0.07 | 0.08 | 0.24 |
| Age Groups | Computed Mean Length (cm) | S.D. | S.I. |
|---|---|---|---|
| Females | |||
| 0+ | 11.21 | 0.63 | n.a. |
| I | 13.32 | 0.46 | 2.17 |
| II | 14.92 | 0.78 | 2.05 |
| III | 17.0 | 0.64 | 2.08 |
| Males | |||
| 0+ | 11.14 | 0.93 | n.a. |
| I | 13.55 | 0.40 | 2.18 |
| II | 14.77 | 0.60 | 2.03 |
| III | 16.61 | 0.91 | 2.04 |
| IV | 18.50 | 1.0 | 2.0 |
| Source | Area | Method | Size (cm) | Age (yrs) | Sex | L∞ (cm) | k (yrs−1) | t0 (yrs) | Φ′ |
|---|---|---|---|---|---|---|---|---|---|
| Atlantic | |||||||||
| [56] | White Cape (NE Atlantic) | Otoliths | C | 34.3 | 0.180 | −0.490 | 2.32 | ||
| Cape Verde (NE Atlantic) | C | 32.5 | 0.180 | −0.620 | 2.27 | ||||
| [57] | Southern Morocco (NE Atlantic) | Scales | C | 30.34 | 0.191 | −1.638 | 2.25 | ||
| M | 30.24 | 0.164 | −1.974 | 2.18 | |||||
| F | 31.55 | 0.181 | −1.853 | 2.26 | |||||
| Mediterranean Sea | |||||||||
| [46] | Tunisian coasts (SW Mediterranean) | Scales | 1–7 | C | 33.54 | 0.191 | −1.434 | 2.33 | |
| 1–7 | M | 33.89 | 0.184 | −1.592 | 2.32 | ||||
| 1–7 | F | 35.93 | 0.156 | −1.848 | 2.29 | ||||
| [47] | Eastern Algeria (SW Mediterranean) | Scales | 1–8 | C | 37.26 | 0.2 | 0.5 | 2.45 | |
| 11.5–26.1 | 1–8 | M | 37.26 | 0.2 | 0.5 | 2.45 | |||
| 10.5–26.7 | 1–7 | F | 36.64 | 0.2 | 0.5 | 2.36 | |||
| [48] | Saros Bay (NE Mediterranean) | Otoliths | 9.0– 22.0 | 1–3 | C | 25.31 | 0.49 | −0.30 | 2.16 |
| [49] | Izmir Bay (NE Mediterranean) | Otoliths | 7.0–22.1 | 1–5 | C | 27.43 | 0.21 | −0.68 | 2.20 |
| Present study | South Aegean Sea (NE Mediterranean) | Otoliths | 9.2–19.2 | 1–4 | C | 23.14 | 0.275 | −1.94 | 2.17 |
| 9.6–19.2 | 1–4 | M | 24.07 | 0.239 | −2.26 | 2.14 | |||
| 9.2–18.1 | 1–3 | F | 23.08 | 0.272 | −1.93 | 2.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legaki, A.; Leonhard, I.; Mytilineou, C.; Anastasopoulou, A. Dentex maroccanus Valenciennes, 1830 Otolith Morphology, Age, and Growth in the Aegean Sea (E. Mediterranean). Animals 2024, 14, 3151. https://doi.org/10.3390/ani14213151
Legaki A, Leonhard I, Mytilineou C, Anastasopoulou A. Dentex maroccanus Valenciennes, 1830 Otolith Morphology, Age, and Growth in the Aegean Sea (E. Mediterranean). Animals. 2024; 14(21):3151. https://doi.org/10.3390/ani14213151
Chicago/Turabian StyleLegaki, Aglaia, Isabella Leonhard, Chryssi Mytilineou, and Aikaterini Anastasopoulou. 2024. "Dentex maroccanus Valenciennes, 1830 Otolith Morphology, Age, and Growth in the Aegean Sea (E. Mediterranean)" Animals 14, no. 21: 3151. https://doi.org/10.3390/ani14213151
APA StyleLegaki, A., Leonhard, I., Mytilineou, C., & Anastasopoulou, A. (2024). Dentex maroccanus Valenciennes, 1830 Otolith Morphology, Age, and Growth in the Aegean Sea (E. Mediterranean). Animals, 14(21), 3151. https://doi.org/10.3390/ani14213151