
Effect of Supplementation with Black Soldier Fly Extract on Intestinal Function in Piglets Infected with Porcine Epidemic Diarrhea Virus

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Trial
2.2. Sample Collection
2.3. Blood Biochemical Measurements in Plasma
2.4. Determination of D-Xylose and Diamine Oxidase (DAO) Activity in Plasma
2.5. Antioxidant Capacity and Oxidation-Relevant Products in Plasma and Intestinal Tissue
2.6. Intestinal Histomorphology
2.7. Real-Time PCR
2.8. Statistical Analyses
3. Results
3.1. Average Daily Weight Gain of Piglets
3.2. Plasma Biochemical Parameters
3.3. Blood DAO Activity and D-Xylose Concentrations
3.4. Levels of Antioxidant Enzymes and Oxidation-Related Products in Plasma and Intestinal Tissues
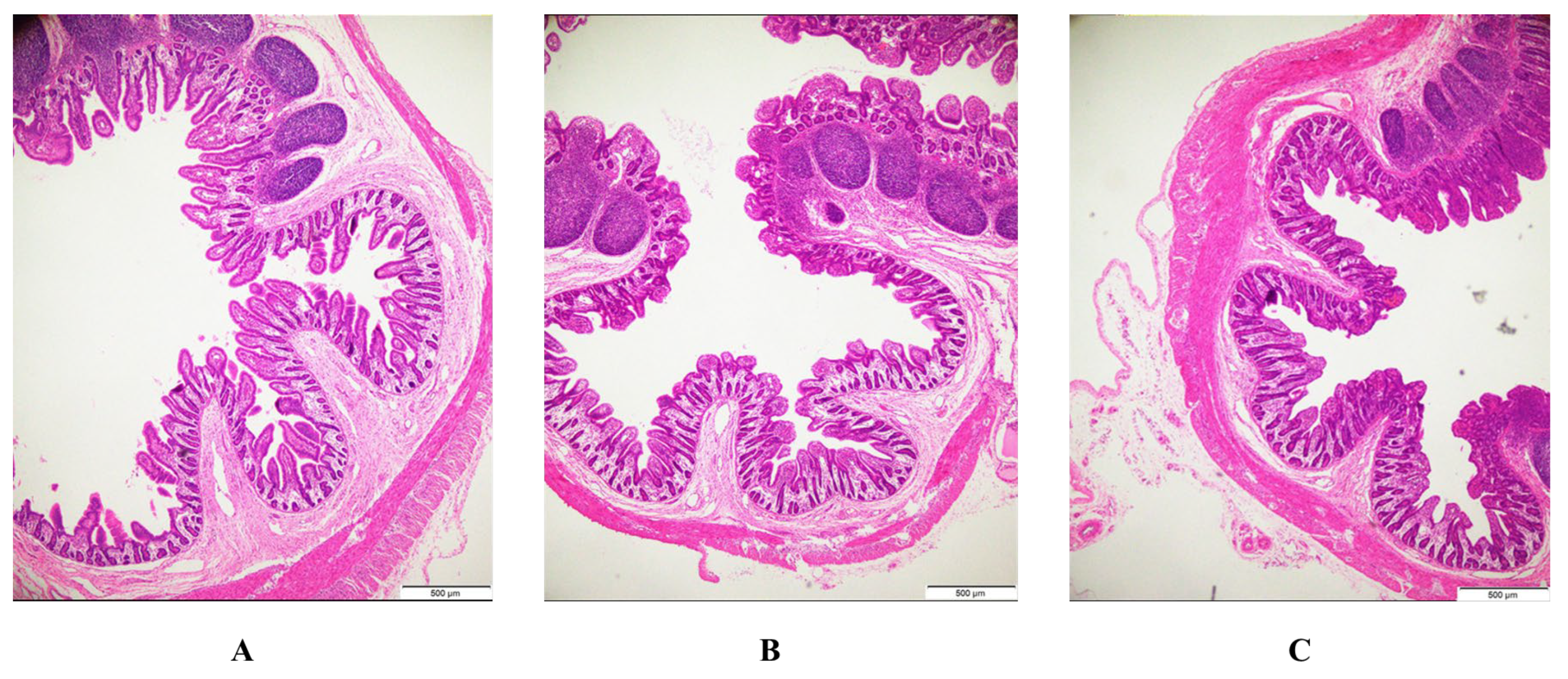
3.5. Intestinal Morphology
3.6. IFN-β, MX1, ISG15 and OASL, and Villin mRNA Levels in the Jejunum, Ileum, and Colon Tissue Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wood, E.N. An apparently new syndrome of porcine epidemic diarrhoea. Vet. Rec. 1977, 100, 243–244. [Google Scholar] [CrossRef]
- Debouck, P.; Pensaert, M. Experimental infection of pigs with a new porcine enteric coronavirus, CV 777. Am. J. Vet. Res. 1980, 41, 219–223. [Google Scholar] [PubMed]
- Puranaveja, S.; Poolperm, P.; Lertwatcharasarakul, P.; Kesdaengsakonwut, S.; Boonsoongnern, A.; Urairong, K.; Kitikoon, P.; Choojai, P.; Kedkovid, R.; Teankum, K.; et al. Chinese-like strain of porcine epidemic diarrhea virus, Thailand. Emerg. Infect. Dis. 2009, 15, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J. Porcine epidemic diarrhea virus infection: Etiology, epidemiology, pathogenesis and immunoprophylaxis. Vet. J. 2015, 204, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Fang, L.; Xiao, S. Porcine epidemic diarrhea in China. Virus Res. 2016, 226, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Liu, T.; Awasthi, S.K.; Duan, Y.; Pandey, A.; Zhang, Z. Manure pretreatments with black soldier fly Hermetia illucens L. (Diptera: Stratiomyidae): A study to reduce pathogen content. Sci. Total Environ. 2020, 737, 139842. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Han, C.; Wang, Z.; Li, Z.; Ruan, L.; Lin, H.; Zhou, C. Black soldier fly pulp in the diet of golden pompano: Effect on growth performance, liver antioxidant and intestinal health. Fish. Shellfish. Immunol. 2023, 142, 109156. [Google Scholar] [CrossRef] [PubMed]
- Waśko, A.; Bulak, P.; Polak-Berecka, M.; Nowak, K.; Polakowski, C.; Bieganowski, A. The first report of the physicochemical structure of chitin isolated from Hermetia illucens. Int. J. Biol. Macromol. 2016, 92, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, C.Y.; Li, C.; Xu, Z.; Peng, P.; Xue, C.; Tomberlin, J.K.; Hu, W.; Cao, Y. Black soldier fly (Hermetia illucens L.) larval diet improves CD8+ lymphocytes proliferation to eliminate chicken coronavirus at an early infection stage. Vet. Microbiol. 2021, 260, 109151. [Google Scholar] [CrossRef] [PubMed]
- Ipema, A.F.; Bokkers, E.A.M.; Gerrits, W.J.J.; Kemp, B.; Bolhuis, J.E. Providing live black soldier fly larvae (Hermetia illucens) improves welfare while maintaining performance of piglets post-weaning. Sci. Rep. 2021, 11, 7371. [Google Scholar] [CrossRef]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Ma, X. Hermetia illucens larvae as a potential dietary protein source altered the microbiota and modulated mucosal immune status in the colon of finishing pigs. J. Anim. Sci. Biotechnol. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Yuan, B.; Liu, M.; Zhu, M.; Zhang, X.; Xie, G.; Wu, W.; Wang, Z.; Xu, H.; Lv, Y.; et al. Dietary Hermetia illucens Larvae Replacement Alleviates Diarrhea and Improves Intestinal Barrier Function in Weaned Piglets Challenged With Enterotoxigenic Escherichia coli K88. Front. Vet. Sci. 2021, 8, 746224. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, J.; Hou, Y.; Yi, D.; Ding, B.; Xie, J.; Zhang, Y.; Chen, H.; Wu, T.; Zhao, D.; et al. N-Acetylcysteine supplementation alleviates intestinal injury in piglets infected by porcine epidemic diarrhea virus. Amino Acids 2017, 49, 1931–1943. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Zheng, L.; Hou, Y.; Guo, S.; Wang, L.; Zhu, L.; Deng, C.; Wu, T.; Yi, D.; et al. Tannic acid-chelated zinc supplementation alleviates intestinal injury in piglets challenged by porcine epidemic diarrhea virus. Front. Vet. Sci. 2022, 9, 1033022. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, L.; Ding, B.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Wu, X.; Yin, Y.; Wu, G. Dietary alpha-ketoglutarate supplementation ameliorates intestinal injury in lipopolysaccharide-challenged piglets. Amino Acids 2010, 39, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Li, D.; Yin, Y.; Wang, X.; Li, P.; Dangott, L.J.; Hu, W.; Wu, G. Intrauterine growth restriction affects the proteomes of the small intestine, liver, and skeletal muscle in newborn pigs. J. Nutr. 2008, 138, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yi, D.; Hou, Y.; Ding, B.; Li, K.; Li, B.; Zhu, H.; Liu, Y.; Wu, G. Dietary Supplementation with α-Ketoglutarate Activates mTOR Signaling and Enhances Energy Status in Skeletal Muscle of Lipopolysaccharide-Challenged Piglets. J. Nutr. 2016, 146, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Abdullah, N.; Idrus, Z.; Ebrahimi, M.; Goh, Y.M.; Shakeri, M.; Oskoueian, A. Palm kernel cake extract exerts hepatoprotective activity in heat-induced oxidative stress in chicken hepatocytes. BMC Complement. Altern. Med. 2014, 14, 368. [Google Scholar] [CrossRef] [PubMed]
- Yi, D.; Li, B.; Hou, Y.; Wang, L.; Zhao, D.; Chen, H.; Wu, T.; Zhou, Y.; Ding, B.; Wu, G. Dietary supplementation with an amino acid blend enhances intestinal function in piglets. Amino Acids 2018, 50, 1089–1100. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Yi, D.; Ding, B.; Chen, X.; Wang, Q.; Zhu, H.; Liu, Y.; Yin, Y.; Gong, J.; et al. Dietary supplementation with tributyrin alleviates intestinal injury in piglets challenged with intrarectal administration of acetic acid. Br. J. Nutr. 2014, 111, 1748–1758. [Google Scholar] [CrossRef]
- Shen, J.; Chen, Y.; Wang, Z.; Zhou, A.; He, M.; Mao, L.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; et al. Coated zinc oxide improves intestinal immunity function and regulates microbiota composition in weaned piglets. Br. J. Nutr. 2014, 111, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lei, Y.; Ye, G.; Sun, L.; Fang, L.; Xiao, S.; Fu, Z.F.; Yin, P.; Song, Y.; Peng, G. Identification of two antiviral inhibitors targeting 3C-like serine/3C-like protease of porcine reproductive and respiratory syndrome virus and porcine epidemic diarrhea virus. Vet. Microbiol. 2018, 213, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.; Dicke, M.; Loon, V.J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed. 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Kim, Y.B.; Kim, D.H.; Jeong, S.B.; Lee, J.W.; Kim, T.H.; Lee, H.G.; Lee, K.W. Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 2020, 99, 3133–3143. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, Q.; Yi, D.; Wu, T.; Chen, H.; Guo, S.; Li, S.; Ji, C.; Wang, L.; Zhao, D.; et al. Quantitative Proteomic Analysis Reveals Antiviral and Anti-inflammatory Effects of Puerarin in Piglets Infected With Porcine Epidemic Diarrhea Virus. Front. Immunol. 2020, 11, 169. [Google Scholar] [CrossRef]
- Curry, S.M.; Gibson, K.A.; Burrough, E.R.; Schwartz, K.J.; Yoon, K.J.; Gabler, N.K. Nursery pig growth performance and tissue accretion modulation due to porcine epidemic diarrhea virus or porcine deltacoronavirus challenge. J. Anim. Sci. 2017, 95, 173–181. [Google Scholar] [CrossRef]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol J. 2015, 12, 193. [Google Scholar] [CrossRef]
- BMansoori, H.; Nodeh, M.; Modirsanei, S.; Rahbari, P. Aparnak, d-Xylose absorption test: A tool for the assessment of the effect of anticoccidials on the intestinal absorptive capacity of broilers during experimental coccidiosis. Anim. Feed. Sci. Technol. 2009, 148, 301–308. [Google Scholar] [CrossRef]
- Luk, G.D.; Bayless, T.M.; Baylin, S.B. Plasma postheparin diamine oxidase. Sensitive provocative test for quantitating length of acute intestinal mucosal injury in the rat. J. Clin. Investig. 1983, 71, 1308–1315. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, T.; Li, S.; Meng, Y.; Tan, Z.; Wu, M.; Yi, D.; Wang, L.; Zhao, D.; Hou, Y. Protective Effect of Zinc Oxide and Its Association with Neutrophil Degranulation in Piglets Infected with Porcine Epidemic Diarrhea Virus. Oxid. Med. Cell Longev. 2021, 2021, 3055810. [Google Scholar] [CrossRef]
- Yu, B.P. Cellular defenses against damage from reactive oxygen species. Physiol. Rev. 1994, 74, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, Y.; Che, Z.; Zhu, H.; Meng, G.; Hou, Y.; Ding, B.; Yin, Y.; Chen, F. Dietary L-arginine supplementation alleviates liver injury caused by Escherichia coli LPS in weaned pigs. Innate Immun. 2012, 18, 804–814. [Google Scholar] [CrossRef]
- Rajarapu, S.P.; Mamidala, P.; Herms, D.A.; Bonello, P.; Mittapalli, O. Antioxidant genes of the emerald ash borer (Agrilus planipennis): Gene characterization and expression profiles. J. Insect Physiol. 2011, 57, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Huang, X.; Tu, F.; Wang, C.; Yang, F. Preparation, antioxidant activity evaluation, and identification of antioxidant peptide from black soldier fly (Hermetia illucens L.) larvae. J. Food Biochem. 2020, 44, e13186. [Google Scholar] [CrossRef] [PubMed]
- Phaengphairee, P.; Boontiam, W.; Wealleans, A.; Hong, J.; Kim, Y.Y. Dietary supplementation with full-fat Hermetia illucens larvae and multi-probiotics, as a substitute for antibiotics, improves the growth performance, gut health, and antioxidative capacity of weaned pigs. BMC Vet. Res. 2023, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Giannenas, I.; Bonos, E.; Anestis, V.; Filioussis, G.; Papanastasiou, D.K.; Bartzanas, T.; Papaioannou, N.; Tzora, A.; Skoufos, I. Effects of Protease Addition and Replacement of Soybean Meal by Corn Gluten Meal on the Growth of Broilers and on the Environmental Performances of a Broiler Production System in Greece. PLoS ONE 2017, 12, e0169511. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Ferrocino, I.; Colombino, E.; Gai, F.; Schiavone, A.; Cocolin, L.; Vincenti, V.; Capucchio, M.T.; Gasco, L. Effects of dietary Hermetia illucens meal inclusion on cecal microbiota and small intestinal mucin dynamics and infiltration with immune cells of weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, J.; Li, L.; Duan, Y.; Zhang, X.; Wang, T.; Zang, J.; Piao, X.; Ma, Y.; Li, D. Endogenous chitinase might lead to differences in growth performance and intestinal health of piglets fed different levels of black soldier fly larva meal. Anim. Nutr. 2023, 14, 411–424. [Google Scholar] [CrossRef]
- Godino, R.F.; Fernández, A.I. New polymorphism of the influenza virus resistance Mx1 gene in Iberian domestic pigs. Postdoc. J. 2016, 4, 15–19. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Q.; Ji, C.; Hu, Y.; Yi, D.; Wu, T.; Wang, L.; Zhao, D.; Hou, Y. Effects of monolaurin on intestinal barrier, blood biochemical profile, immunity and antioxidant function in porcine epidemic diarrhoea virus-infected piglets. Br. J. Nutr. 2024, 131, 185–192. [Google Scholar] [CrossRef]
- Wang, S.; Wu, J.; Wang, F.; Wang, H.; Wu, Z.; Wu, S.; Bao, W. Expression Pattern Analysis of Antiviral Genes and Inflammatory Cytokines in PEDV-Infected Porcine Intestinal Epithelial Cells. Front. Vet. Sci. 2020, 7, 75. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Gene Name | Primer | Amplicon Size (bp) | |
|---|---|---|---|
| RPL4 | Forward | 5′-GAGAAACCGTCGCCGAAT-3′ | 146 |
| Reverse | 5′-GCCCACCAGGAGCAAGTT-3′ | ||
| IFN-β | Forward | 5′-AGCAGATCTTCGGCATTCTC-3′ | 101 |
| Reverse | 5′-GTCATCCATCTGCCCATCAA-3′ | ||
| MX1 | Forward | 5′-AGTGCGGCTGTTTACCAAG-3′ | 150 |
| Reverse | 5′-TTCACAAACCCTGGCAACTC-3′ | ||
| ISG15 | Forward | 5′-AGCATGGTCCTGTTGATGGTG-3′ | 164 |
| Reverse | 5′-CAGAAATGGTCAGCTTGCACG-3′ | ||
| OASL | Forward | 5′-GGCACCCCTGTTTTCCTCT-3′ | 139 |
| Reverse | 5′-AGCACCGCTTTTGGATGG-3′ |
| Item | Control | PEDV | BFE + PEDV | p-Value |
|---|---|---|---|---|
| Days 1–8 (pre-challenge) | ||||
| ADG (kg) | 0.124 ± 0.029 | 0.123 ± 0.039 | 0.129 ± 0.019 | 0.934 |
| Days 9–11 (post-challenge) | ||||
| ADG (kg) | 0.109 ± 0.025 a | 0.050 ± 0.058 b | 0.068 ± 0.060 ab | 0.034 |
| Item | Control | PEDV | BFE + PEDV | p-Value |
|---|---|---|---|---|
| TB (mg/dL) | 0.239 ± 0.063 a | 0.194 ± 0.072 a | 0.107 ± 0.040 b | 0.002 |
| ALT (U/L) | 38.300 ± 7.364 | 42.700 ± 8.394 | 37.500 ± 6.091 | 0.315 |
| ALP (U/L) | 720.000 ± 141.966 a | 561.500 ± 133.213 b | 584.500 ± 98.806 b | 0.029 |
| TC (mg/dL) | 134.322 ± 23.446 a | 99.646 ± 17.311 b | 94.947 ± 10.892 b | <0.001 |
| TG (mg/dL) | 31.880 ± 6.526 b | 45.407 ± 12.005 a | 46.341 ± 5.467 a | 0.001 |
| GLU (mg/dL) | 96.110 ± 8.478 b | 95.400 ± 7.415 b | 107.017 ± 11.594 a | 0.040 |
| CA (mg/dL) | 12.717 ± 0.886 a | 11.450 ± 0.991 b | 11.512 ± 0.843 b | 0.010 |
| CREA (mg/dL) | 0.678 ± 0.076 | 0.687 ± 0.050 | 0.753 ± 0.096 | 0.127 |
| HDL (mg/dL) | 19.062 ± 4.446 | 17.145 ± 2.689 | 18.082 ± 2.748 | 0.790 |
| BUN (mg/dL) | 2.200 ± 0.607 b | 6.700 ± 1.372 a | 5.800 ± 1.694 a | <0.001 |
| GGT (U/L) | 29.000 ± 4.397 | 24.180 ± 6.983 | 29.361 ± 6.612 | 0.146 |
| CK (mg/dL) | 281.500 ± 65.129 a | 178.480 ± 33.810 b | 174.000 ± 38.601 b | <0.001 |
| LDH (mg/dL) | 752.764 ± 112.468 | 686.747 ± 93.216 | 641.017 ± 63.573 | 0.086 |
| Item | Control | PEDV | BFE + PEDV | p-Value |
|---|---|---|---|---|
| D-xylose (mmol/L) | 1.735 ± 0.393 a | 0.999 ± 0.183 b | 0.941 ± 0.461 b | <0.001 |
| DAO (U/L) | 12.350 ± 8.000 b | 30.300 ± 9.450 a | 28.090 ± 15.190 a | 0.002 |
| Item | Control | PEDV | BFE + PEDV | p-Value |
|---|---|---|---|---|
| T-SOD | ||||
| Plasma (U/mL) | 83.49 ± 1.77 a | 77.34 ± 3.05 b | 80.75 ± 4.38 a | 0.001 |
| Jejunum (U/mg prot) | 312.77 ± 60.84 ab | 344.43 ± 48.61 a | 272.53 ± 25.30 b | 0.035 |
| Ileum (U/mg prot) | 216.73 ± 27.68 b | 209.73 ± 17.52 b | 262.32 ± 28.00 a | 0.001 |
| Colon (U/mg prot) | 27.94 ± 2.73 ab | 24.93 ± 3.59 b | 30.07 ± 5.51 a | 0.042 |
| CAT | ||||
| Plasma (U/mL) | 6.31 ± 1.45 a | 5.13 ± 0.85 b | 7.22 ± 0.84 a | 0.005 |
| Jejunum (U/mg prot) | 2.18 ± 0.25 a | 1.70 ± 0.36 b | 1.77 ± 0.25 b | 0.004 |
| Ileum (U/mg prot) | 0.97 ± 0.29 | 1.03 ± 0.30 | 1.25 ± 0.11 | 0.128 |
| Colon (U/mg prot) | 0.65 ± 0.16 a | 0.38 ± 0.10 b | 0.61 ± 0.19 a | 0.001 |
| MDA | ||||
| Plasma (nmol/mL) | 2.38 ± 0.37 a | 2.33 ± 0.32 a | 1.78 ± 0.27 b | 0.004 |
| Jejunum (nmol/mg prot) | 0.30 ± 0.08 b | 0.38 ± 0.05 a | 0.30 ± 0.05 b | 0.018 |
| Ileum (nmol/mg prot) | 0.95 ± 0.21 b | 1.03 ± 0.19 b | 1.96 ± 0.41 a | <0.001 |
| Colon (nmol/mg prot) | 1.07 ± 0.25 a | 0.95 ± 0.26 a | 0.65 ± 0.16 b | 0.008 |
| H2O2 | ||||
| Plasma (mmol/L) | 37.38 ± 7.03 | 41.55 ± 11.01 | 31.38 ± 7.14 | 0.104 |
| Jejunum (mmol/mg prot) | 3.66 ± 0.56 | 3.22 ± 0.56 | 3.50 ± 0.49 | 0.216 |
| Ileum (mmol/mg prot) | 7.77 ± 1.40 b | 9.13 ± 1.51 ab | 9.82 ± 2.06 a | 0.049 |
| Colon (mmol/mg prot) | 5.51 ± 1.17 b | 7.34 ± 2.12 a | 4.57 ± 1.43 b | 0.008 |
| Item | Control | PEDV | BFE + PEDV | p-Value |
|---|---|---|---|---|
| Villus height (μm) | ||||
| Jejunum | 291.05 ± 46.88 a | 170.79 ± 11.51 b | 191.05 ± 20.30 b | <0.001 |
| Ileum | 384.06 ± 47.37 a | 154.55 ± 30.12 c | 197.17 ± 17.47 b | <0.001 |
| Crypt depth (μm) | ||||
| Jejunum | 102.25 ± 15.88 c | 150.72 ± 15.47 a | 119.14 ± 12.33 b | <0.001 |
| Ileum | 126.85 ± 16.45 b | 164.00 ± 23.96 a | 152.78 ± 13.64 a | 0.001 |
| Villus height/crypt depth | ||||
| Jejunum | 3.01 ± 0.61 a | 1.20 ± 0.17 c | 1.81 ± 0.13 b | <0.001 |
| Ileum | 3.16 ± 0.46 a | 1.01 ± 0.20 b | 1.27 ± 0.11 b | <0.001 |
| Villous surface area (μm2) | ||||
| Jejunum | 19616.55 ± 3759.46 a | 13014.98 ± 1822.92 b | 13324.72 ± 1403.05 b | <0.001 |
| Ileum | 41321.35 ± 6524.20 a | 11069.21 ± 2375.02 c | 15647.26 ± 1505.07 b | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.; Wu, M.; Sun, L.; Li, H.; Xu, Z.; Zhang, Q.; Yi, D.; Wang, L.; Zhao, D.; Hou, Y.; et al. Effect of Supplementation with Black Soldier Fly Extract on Intestinal Function in Piglets Infected with Porcine Epidemic Diarrhea Virus. Animals 2024, 14, 1512. https://doi.org/10.3390/ani14101512
Yu C, Wu M, Sun L, Li H, Xu Z, Zhang Q, Yi D, Wang L, Zhao D, Hou Y, et al. Effect of Supplementation with Black Soldier Fly Extract on Intestinal Function in Piglets Infected with Porcine Epidemic Diarrhea Virus. Animals. 2024; 14(10):1512. https://doi.org/10.3390/ani14101512
Chicago/Turabian StyleYu, Chenmin, Mengjun Wu, Lanyuan Sun, Hanxiao Li, Zhaoyang Xu, Qian Zhang, Dan Yi, Lei Wang, Di Zhao, Yongqing Hou, and et al. 2024. "Effect of Supplementation with Black Soldier Fly Extract on Intestinal Function in Piglets Infected with Porcine Epidemic Diarrhea Virus" Animals 14, no. 10: 1512. https://doi.org/10.3390/ani14101512
APA StyleYu, C., Wu, M., Sun, L., Li, H., Xu, Z., Zhang, Q., Yi, D., Wang, L., Zhao, D., Hou, Y., & Wu, T. (2024). Effect of Supplementation with Black Soldier Fly Extract on Intestinal Function in Piglets Infected with Porcine Epidemic Diarrhea Virus. Animals, 14(10), 1512. https://doi.org/10.3390/ani14101512