
Evaluating the Use of Chemical Weapons for Capturing Prey by a Venomous Mammal, the Greater Slow Loris (Nycticebus coucang)


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Slow Loris Subjects and Housing
2.2. Prey Items
2.3. Behavioral Experiments
2.4. Data Analysis
3. Results
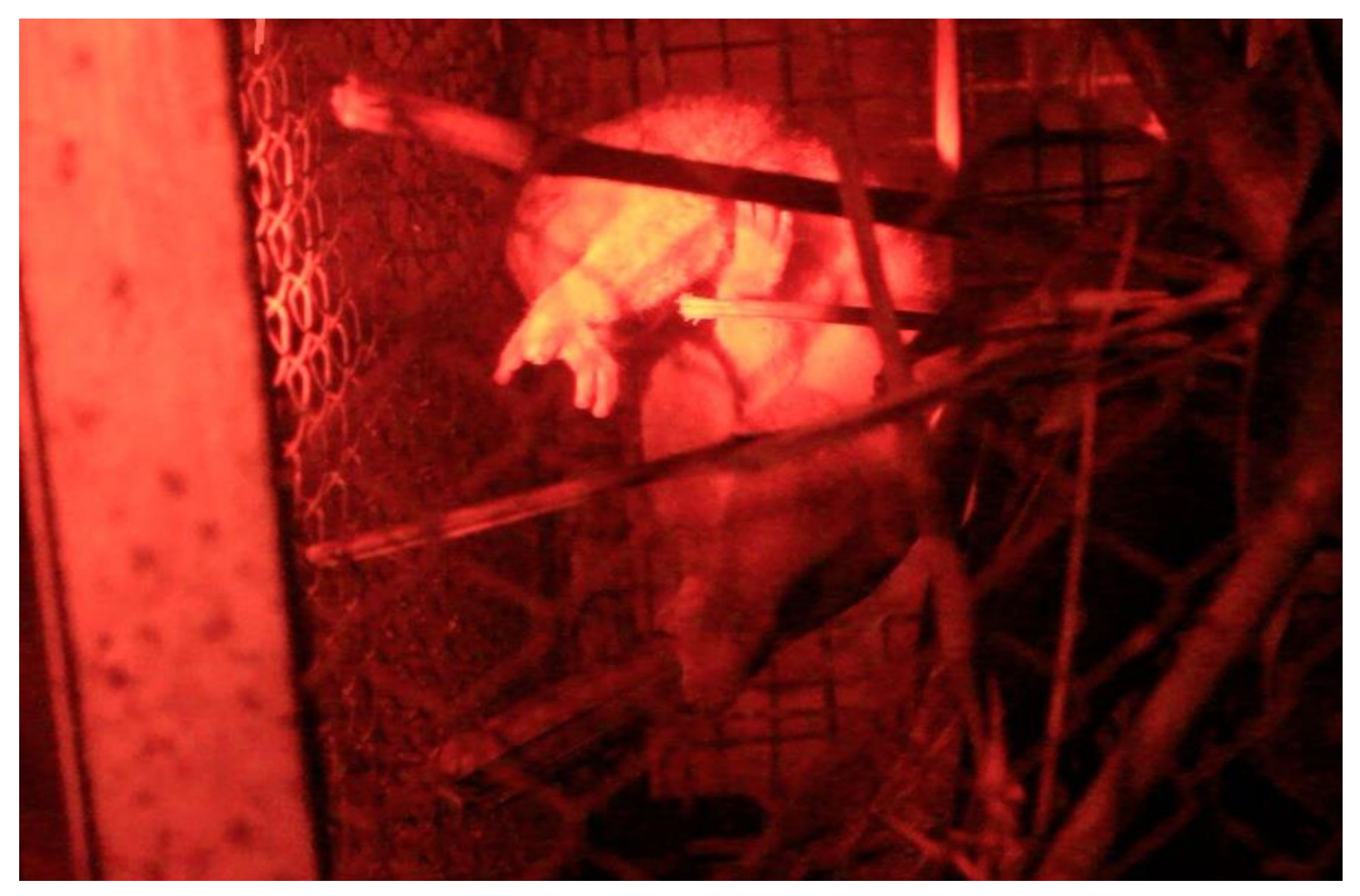
3.1. The Prey Capture Sequence
3.2. Venom-Related Behaviors
3.3. Effects of Prey Attributes
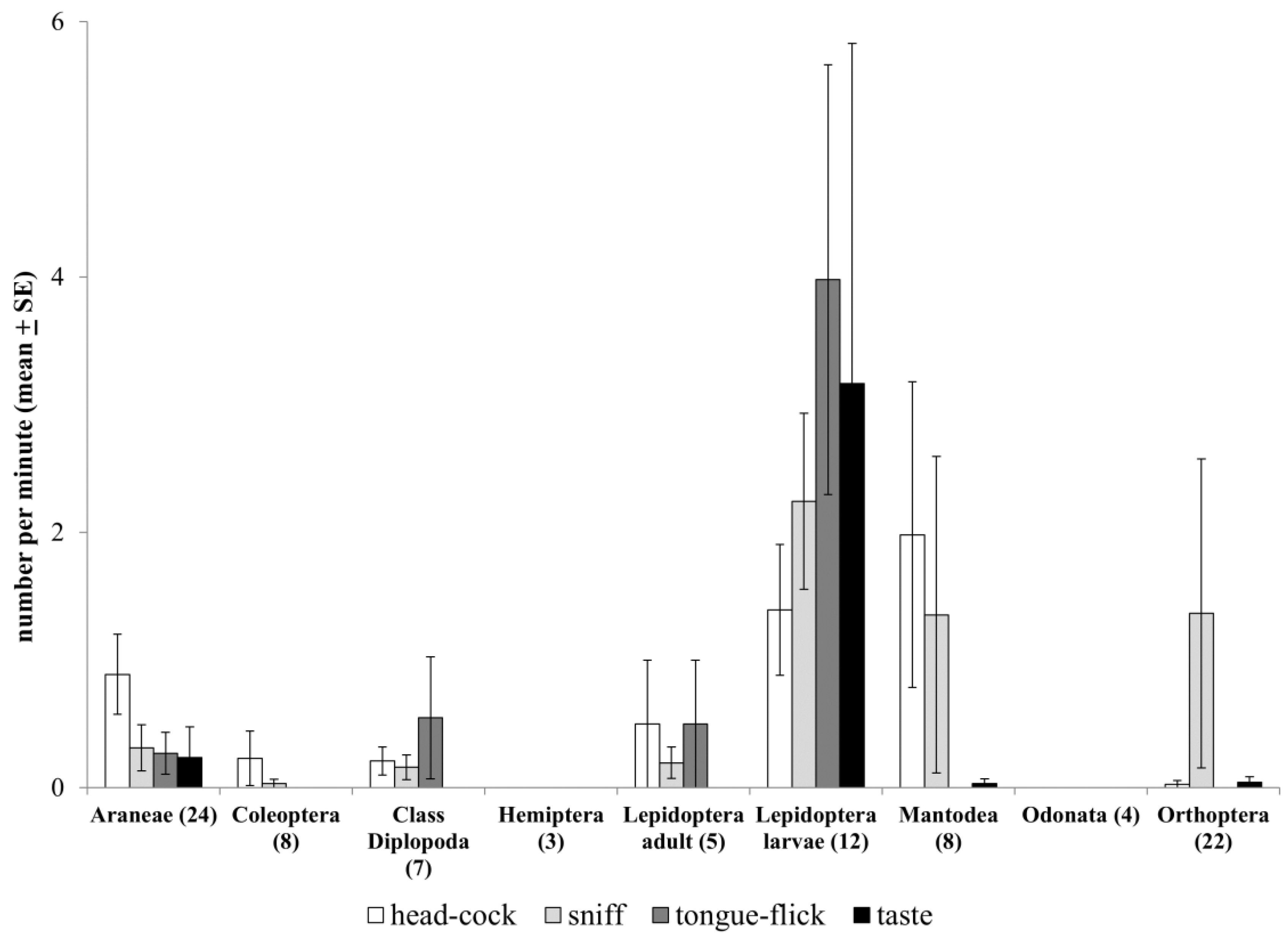
3.4. Effects of Prey Type
4. Discussion
4.1. Evidence for Envenomation of Prey
4.2. Effects of Prey Attributes on Loris Predatory Behavior
4.3. Sensory Modalities Used for Prey Assessment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fitzpatrick, L.L.; Ligabue-Braun, R.; Nekaris, K.A. Slowly Making Sense: A Review of the Two-Step Venom System within Slow (Nycticebus spp.) and Pygmy Lorises (Xanthonycticebus spp.). Toxins 2023, 15, 514. [Google Scholar] [CrossRef] [PubMed]
- Ligabue-Braun, R.; Verli, H.; Carlini, C.R. Venomous mammals: A review. Toxicon 2012, 59, 680–695. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Moore, R.S.; Rode, J.; Fry, B.G. Mad, bad and dangerous to know: The biochemistry, ecology and evolution of slow loris venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 21. [Google Scholar] [CrossRef] [PubMed]
- Alterman, L. Toxins and toothcombs: Potential allospecific chemical defenses in Nycticebus and Perodicticus. In Creatures of the Dark; Alterman, L., Doyle, G., Izard, M.K., Eds.; Springer: New York, NY, USA, 1995; pp. 413–424. [Google Scholar]
- Wilde, H. Anaphylactic shock following bite by a slow loris, Nycticebus coucang. Am. J. Trop. Med. Hyg. 1972, 21, 592–594. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Rode-Margono, E.J.; Rademaker, M.; Wirdateti; Strijkstra, A.; Nekaris, K.A.I. Noxious arthropods as potential prey of the venomous Javan slow loris (Nycticebus javanicus) in a West Javan volcanic agricultural system. J. Nat. Hist. 2015, 49, 1949–1959. [Google Scholar] [CrossRef]
- Cabana, F.; Dierenfeld, E.; Wirdateti, W.; Donati, G.; Nekaris, K.A.I. The seasonal feeding ecology of the Javan slow loris (Nycticebus javanicus). Am. J. Phys. Anthropol. 2017, 162, 768–781. [Google Scholar] [CrossRef] [PubMed]
- Starr, C.; Nekaris, K.A.I. Obligate exudativory characterizes the diet of the pygmy slow loris Nycticebus pygmaeus. Am. J. Primatol. 2013, 75, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Streicher, U.; Wilson, A.; Collins, R.L.; Nekaris, K.A.-I. Exudates and Animal Prey Characterize Slow Loris (Nycticebus pygmaeus, N. coucang and N. javanicus) Diet in Captivity and After Release into the Wild. In Leaping Ahead: Advances in Prosimian Biology; Masters, J., Gamba, M., Génin, F., Eds.; Springer: New York, NY, USA, 2013; pp. 165–172. [Google Scholar]
- Wiens, F.; Zitzmann, A.; Hussein, N.A. Fast food for slow lorises: Is low metabolism related to secondary compounds in high-energy plant diet? J. Mammal. 2006, 87, 790–798. [Google Scholar] [CrossRef]
- Nekaris, K.; Campera, M.; Nijman, V.; Birot, H.; Rode-Margono, E.J.; Fry, B.G.; Weldon, A.; Wirdateti, W.; Imron, M.A. Slow lorises use venom as a weapon in intraspecific competition. Curr. Biol. 2020, 30, R1252–R1253. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Rode-Margono, J.E.; Nekaris, K.A.-I. Cabinet of Curiosities: Venom Systems and Their Ecological Function in Mammals, with a Focus on Primates. Toxins 2015, 7, 2639–2658. [Google Scholar] [CrossRef] [PubMed]
- Pearson, O.P. On the cause and nature of a poisonous action produced by the bite of a shrew (Blarina brevicauda). J. Mammal. 1942, 23, 159–166. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The venom optimization hypothesis revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Hagey, L.R.; Fry, B.G.; Fitch-Snyder, H. Talking defensively: A dual use for the brachial gland exudate of slow and pygmy lorises. In Primate Anti-Predator Strategies; Gursky, S., Nekaris, K.A.I., Eds.; Springer: New York, NY, USA, 2007; pp. 251–270. [Google Scholar]
- Wigger, E.; Kuhn-Nentwig, L.; Nentwig, W. The venom optimisation hypothesis: A spider injects large venom quantities only into difficult prey types. Toxicon 2002, 40, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Nisani, Z.; Hayes, W.K. Defensive stinging by Parabuthus transvaalicus scorpions: Risk assessment and venom metering. Anim. Behav. 2011, 81, 627–633. [Google Scholar] [CrossRef]
- Nekaris, K.A.I. Foraging behaviour of the slender loris (Loris lydekkerianus lydekkerianus): Implications for theories of primate origins. J. Hum. Evol. 2005, 49, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Schiel, N.; Souto, A.; Huber, L.; Bezerra, B.M. Hunting strategies in wild common marmosets are prey and age dependent. Am. J. Primatol. 2010, 72, 1039–1046. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 48, 227–265. [Google Scholar] [CrossRef]
- Chatpongcharoen, P.; Campera, M.; Laithong, P.; Gibson, N.L.; Nekaris, K. Naturalising diet to reduce stereotypic behaviours in slow lorises rescued from wildlife trade. Appl. Anim. Behav. Sci. 2021, 242, 105413. [Google Scholar] [CrossRef]
- Cooper, W.E. Foraging mode and evolution of strike-induced chemosensory searching in lizards. J. Chem. Ecol. 2003, 29, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Maynard, K.Q.; Birot, H.; Campera, M.; Imron, M.A.; Jasso del Toro, C.; Poindexter, S.A.; Nekaris, K.A.I. Slow learning of feeding skills in a nocturnal extractive forager. Anim. Behav. 2021, 173, 1–7. [Google Scholar] [CrossRef]
- Sherbrooke, W.C.; Schwenk, K. Horned lizards (Phrynosoma) incapacitate dangerous ant prey with mucus. J. Exp. Zool. Part A-Ecol. Genet. Physiol. 2008, 309A, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Grow, N.B.; Wirdateti; Nekaris, K.A.I. Does toxic defence in Nycticebus spp. relate to ectoparasites? The lethal effects of slow loris venom on arthropods. Toxicon 2015, 95, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tandler, B.; Pinkstaff, C.A.; Nagato, T.; Phillips, C.J. Giant secretory granules in the ducts of the parotid and submandibular glands of the slow loris. Tissue Cell 1996, 28, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Lappin, A.K.; German, M. Feeding behavior modulation in the leopard lizard (Gambelia wislizenii): Effects of noxious versus innocuous prey. Zoology 2005, 108, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Kornilev, Y.V.; Natchev, N.D.; Lillywhite, H.B. Perils of ingesting harmful prey by advanced snakes. Biol. Rev. 2023, 98, 263–283. [Google Scholar] [CrossRef] [PubMed]
- Nijman, V.; Spaan, D.; Rode-Margono, E.J.; Wirdateti; Nekaris, K.A.I. Changes in the primate trade in Indonesian wildlife markets over a 25-year period: Fewer apes and langurs, more macaques, and slow lorises. Am. J. Primatol. 2017, 79, e22517. [Google Scholar] [CrossRef] [PubMed]
- Fuller, G.; Eggen, W.F.; Wirdateti, W.; Nekaris, K.A.I. Welfare impacts of the illegal wildlife trade in a cohort of confiscated greater slow lorises, Nycticebus coucang. J. Appl. Anim. Welf. Sci. 2018, 21, 224–238. [Google Scholar] [CrossRef]
- Glendinning, J.I. How Do Predators Cope with Chemically Defended Foods? Biol. Bull. 2007, 213, 252–266. [Google Scholar] [CrossRef]
- Nekaris, K.A.-I.; Weldon, A.; Imron, M.A.; Maynard, K.Q.; Nijman, V.; Poindexter, S.A.; Morcatty, T.Q. Venom in furs: Facial masks as aposematic signals in a venomous mammal. Toxins 2019, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Campera, M.; Watkins, A.R.; Weldon, A.V.; Hedger, K.; Morcatty, T.Q. Aposematic signaling and seasonal variation in dorsal pelage in a venomous mammal. Ecol. Evol. 2021, 11, 11387–11397. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Pimley, E.R.; Ablard, K.M. Predator defense by slender lorises and pottos. In Primate Anti-Predator Strategies; Gursky, S., Nekaris, K.A.I., Eds.; Springer: New York, NY, USA, 2007; pp. 220–238. [Google Scholar]
- Evans, C.S. Accessory chemosignaling mechanisms in primates. Am. J. Primatol. 2006, 68, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.K.; Kanagasu, R. Vomeronasal organ in tree shrew and slow loris. J. Anat. 1972, 112, 165. [Google Scholar] [PubMed]
- Kendrick, K.M.; Dixson, A.F. A quantitative description of copulatory and associated behaviors of captive marmosets (Callithrix jacchus). Int. J. Primatol. 1984, 5, 199–212. [Google Scholar] [CrossRef]
- Fisher, H.S.; Swaisgood, R.R.; Fitch-Snyder, H. Odor familiarity and female preferences for males in a threatened primate, the pygmy loris Nycticebus pygmaeus: Applications for genetic management of small populations. Naturwissenschaften 2003, 90, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Fisher, H.S.; Swaisgood, R.R.; Fitch-Snyder, H. Countermarking by male pygmy lorises (Nycticebus pygmaeus): Do females use odor cues to select mates with high competitive ability? Behav. Ecol. Sociobiol. 2003, 53, 123–130. [Google Scholar] [CrossRef]
- Martinez-Marcos, A.; Ubeda-Banon, I.; Halpern, M. Neural substrates for tongue-flicking behavior in snakes. J. Comp. Neurol. 2001, 432, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, K. Why snakes have forked tongues. Science 1994, 263, 1573–1577. [Google Scholar] [CrossRef]
- Nadler, T.; Schwierz, E.; Streicher, U. Pygmy lorises (Nycticebus pygmaeus) without sublingua. Vietnam. J. Primatol. 2013, 2, 83–86. [Google Scholar]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A modest proposal: Displacement activities as an indicator of emotions in primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Chertoff, S.; Wandia, I.N.; Leca, J.-B. Tongue-flicking: An idiosyncratic displacement behaviour in a free-ranging and urban-dwelling population of Balinese long-tailed macaques. Behaviour 2023, 160, 577–591. [Google Scholar] [CrossRef]
- Pariente, G. The role of vision in prosimian behavior. In The Study of Prosimian Behavior; Doyle, G.A., Martin, R.D., Eds.; Academic Press: New York, NY, USA, 1979; pp. 411–459. [Google Scholar]
- Cartmill, M. New views on primate origins. Evol. Anthropol. 1992, 1, 105–111. [Google Scholar] [CrossRef]
- Page, R.A.; Schnelle, T.; Kalko, E.K.V.; Bunge, T.; Bernal, X.E. Sequential assessment of prey through the use of multiple sensory cues by an eavesdropping bat. Naturwissenschaften 2012, 99, 505–509. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Order: Family | Species | N |
|---|---|---|
| Araneae | Total | 19 |
| Araneidae | Argiope sp. | 3 |
| Araneidae | unknown sp. | 7 |
| Nephilidae | Nephila pilipes | 6 |
| Sparassidae | unknown sp. | 3 |
| Coleoptera | Total | 7 |
| Cerambycidae | Xystrocera festiva | 4 |
| Scarabaeidae | Phyllophaga sp. | 1 |
| Scarabaeidae | Xylotrupes gideon | 1 |
| Scarabaeidae | Xylotrupes sp. | 1 |
| Class Diplopoda (order unknown) | Total unknown sp. | 5 |
| Hemiptera | Total | 2 |
| Cicadidae | Dundubia vaginata | 2 |
| Lepidoptera (adults) | Total | 5 |
| Noctuidae | unknown sp. | 1 |
| Nymphalidae | Charaxes nobilis | 2 |
| Nymphalidae | Junonia sp. | 1 |
| Pyralidae | unknown sp. | 1 |
| Lepidoptera (larvae) | Total | 10 |
| Arctiidae | unknown sp. | 2 |
| Lasiocampidae | unknown sp. | 3 |
| Notodontidae | unknown sp. | 2 |
| Nymphalidae | unknown sp. | 3 |
| Mantodea | Total | 6 |
| Mantidae | Amantis sp. | 1 |
| Mantidae | Hierodula sp. | 1 |
| Mantidae | Hierodula vitrea | 1 |
| Mantidae | Tenodera aridifolia | 3 |
| Odonata | Total | 3 |
| Libellulidae | Orthetrum sabina | 2 |
| Libellulidae | Pantala flavescens | 1 |
| Orthoptera | Total | 18 |
| Acrididae | Valanga nigricornis | 3 |
| Acrididae | Oxya chinensis | 6 |
| Gryllacrididae | Gryllacris signifera | 1 |
| Gryllidae | Acheta domesticus | 1 |
| Gryllidae | Gryllus sp. | 1 |
| Tettigonidae | Conocephalus sp. | 1 |
| Tettigonidae | Holochlora sp. | 1 |
| Tettigonidae | Mecapoda sp. | 1 |
| Tettigonidae | Xiphidion sp. | 1 |
| Tettigonidae | unknown sp. | 2 |
| Arthropods | Total | 75 |
| Behavior | Operational Definition |
|---|---|
| Investigatory and Preparatory Behaviors | |
| scent-marking | Rubbing the face, chest, or perineum along substrates; or urine marking. |
| circling | Radial movement above the arena without descending to the cage floor, while fixing attention on prey. |
| brachial lick | Making contact between the tongue and the brachial gland. |
| lick hands | Using the tongue to apply saliva to the fingers or palms. |
| groom other | Licking or cleaning anywhere on the body except the brachial gland or hands using the tongue, toothcomb, or grooming claw. |
| approach and retreat | Moving away from proximity to the prey item after investigating it with no attempted capture. |
| grab and release | Letting go of the prey item after capture without attempting to consume the item. |
| Sensory Behaviors | |
| visually target | Fixing the gaze on the prey item and maintaining eye contact with it while moving toward capture. |
| head-cocking | Slowly turning the head on a 180-degree axis, while focusing visual attention on the prey item. |
| ear twitch | Visibly moving the ears or moving the head to orient them toward the prey. |
| sniff | Repeatedly moving the nose to rapidly inhale air or moving the nose along a substrate while inhaling. |
| tongue-flicking | Rapidly moving the tongue in and out of the mouth without contacting the prey item or another substrate. |
| taste (lick) | Moving the tongue in contact with the prey without attempting to consume it or using the teeth or lips. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuller, G.; Wirdateti; Nekaris, K.A.I. Evaluating the Use of Chemical Weapons for Capturing Prey by a Venomous Mammal, the Greater Slow Loris (Nycticebus coucang). Animals 2024, 14, 1438. https://doi.org/10.3390/ani14101438
Fuller G, Wirdateti, Nekaris KAI. Evaluating the Use of Chemical Weapons for Capturing Prey by a Venomous Mammal, the Greater Slow Loris (Nycticebus coucang). Animals. 2024; 14(10):1438. https://doi.org/10.3390/ani14101438
Chicago/Turabian StyleFuller, Grace, Wirdateti, and K. A. I. Nekaris. 2024. "Evaluating the Use of Chemical Weapons for Capturing Prey by a Venomous Mammal, the Greater Slow Loris (Nycticebus coucang)" Animals 14, no. 10: 1438. https://doi.org/10.3390/ani14101438
APA StyleFuller, G., Wirdateti, & Nekaris, K. A. I. (2024). Evaluating the Use of Chemical Weapons for Capturing Prey by a Venomous Mammal, the Greater Slow Loris (Nycticebus coucang). Animals, 14(10), 1438. https://doi.org/10.3390/ani14101438