



First Account of the Breeding Biology of Indian Blue Robin (Larvivora brunnea) in Southwest China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Procedures
2.3. Data Analysis
3. Results
3.1. Nest and Eggs
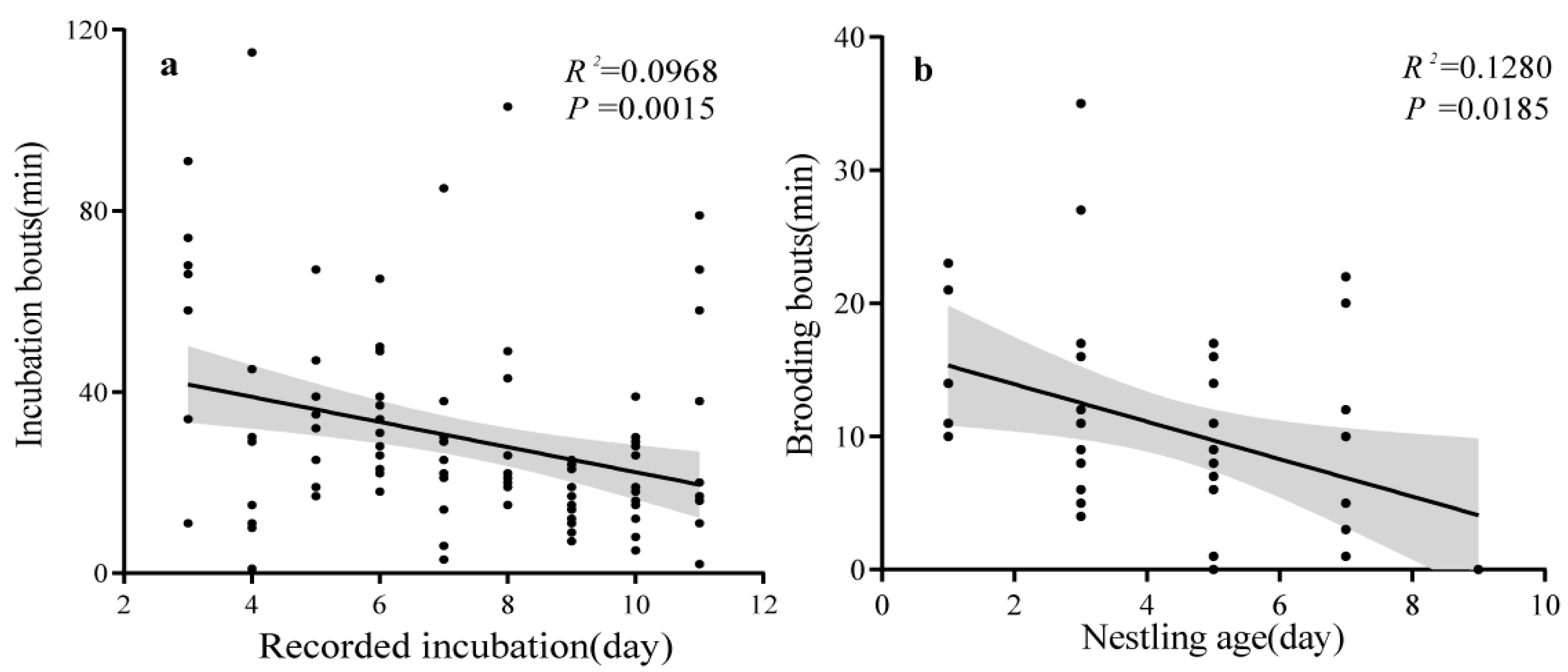
3.2. Incubation
3.3. Parental Care, Growth of Nestlings, and Breeding Success
3.4. Development of Nestlings

{kind=link}
{kind=link}
| Body Measurements | Body Mass/g | Total Length/mm | Culmen /mm | Wing /mm | Tail/mm | Tarsus/mm |
|---|---|---|---|---|---|---|
| 1-day-old nestling a | 2.349 | 32.88 | 3.91 | 11.14 | 0 | 5.87 |
| 3-day-old nestling a | 5.173 | 42.62 | 4.66 | 14.98 | 0 | 10.53 |
| 5-day-old nestling a | 8.780 | 55.82 | 6.13 | 25.28 | 0 | 14.78 |
| 7-day-old nestling a | 13.462 | 59.65 | 6.75 | 31.74 | 2.45 | 22.74 |
| 9-day-old nestling a | 15.417 | 62.96 | 8.71 | 38.24 | 4.46 | 25.57 |
| 11-day-old nestling a | 16.125 | 66.33 | 9.06 | 54.22 | 8.98 | 28.83 |
| Adult male b | 12–18 | 126–152 | 11–14 | 72–83 | 44–66 | 25–30 |
| Adult female b | 14–14 | 125–125 | 12–12 | 73–73 | 51–51 | 25–25 |
4. Discussion
| Species | Nesting Site | Nest | Egg | Incubation Period (d) | Nestling Period (d) | Breeding Month | References | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elevation (m) | Habitat | Clutch Size | Nest Material | Fresh Mass (g) | Size (mm) | Color | Pattern | |||||
| Japanese Robin (Larvivora akahige) | 1000–2500 | Broadleaf evergreen forest, mixed conifer–broadleaf forest | 4–5 | Bryophyte and dead leaves | - | 22.05 × 16.25 | Pale greenish–blue | No speckles | 12–15 | 12 | 5–7 | [7] |
| Ryukyu Robin (Larvivora komadori) | 1100–1600 | Broadleaf evergreen forest, undergrowth | 3–5 | Bryophyte and bamboo leaves | - | 22.5 × 17 | Pinkish | No speckles | - | - | 4–8 | [13] |
| Okinawa Robin (Larvivora namiyei) | 1100–1600 | Broadleaf evergreen forest | - | - | - | - | - | - | - | - | - | [35] |
| Izu Robin (Larvivora tanensis) | - | Occurs in dense undergrowth of mature or old-growth damp, montane, deciduous, and evergreen temperate forest | - | - | - | - | - | - | - | - | - | [36] |
| Rufous-tailed Robin (Larvivora sibilans) | 1200 | Damp broadleaf evergreen and semi-eve rgreen bottomland forest with dense undergrowth | 4–6 | Bryophyte, grass stems, and pine needles | - | 19–20 × 14–16 | Pale blue, grayish blue | brown blotches | - | - | 6–7 | [19,26] |
| Rufous-headed Robin (Larvivora ruficeps) | 2400–2800 | Temperate mixed coniferous and decoduous forest | 4 | - | - | 17.9–19.27 × 15.64–16.49 | - | - | - | - | 5–6 | [19,27] |
| Indian Blue Robin (Larvivora brunnea) | 1600–3300 | Broadleaf evergreen forest, bamboo forest | 4 | Bryophyte, dead leaves, and wool | 2.11–2.36 | 19.7–20.4 × 14.5–15.2 | Plain pale blue | No speckles | ≥13 | 12 | 5–6 | [12]; this study |
| Siberian Blue Robin (Larvivora cyane) | 1100–1500 | Damp broadleaf evergreen and semi-evc rgreen bottomland forest with dense undergrowth, thickets | 5–6 | Bryophyte, dead leaves, and animal hair | 1.8–2.2 | 15.0 × 19.5 | Sky-blue to bright blue or greenish–blue | No speckles | 12–13 | 13 | 5–6 | [5,6,8] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, H.T.; Hu, Y.G.; Lang, Z.D.; Fang, B.H.; Guo, W.B.; Zhang, Q.; Pan, X.; Lu, X. How much do we know about the breeding biology of bird species in the world? J. Avian. Biol. 2017, 48, 513–518. [Google Scholar] [CrossRef]
- Del Hoyo, J.; Elliott, A.; Sargatal, J. Handbook of the Birds of the World; Lynx Edicions: Barcelona, Spain, 1992; Volume 1–17. [Google Scholar]
- Fan, S.X.; Zhang, J.S.; Duan, Y.B.; Luo, X. First Description of the Breeding Biology of the Spectacled Fulvetta (Fulvetta ruficapilla sordidior) in Southwest China. Animals 2023, 13, 2157. [Google Scholar] [CrossRef] [PubMed]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Old World Flycatchers (Muscicapidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Song, Y.J.; Gao, W.; He, J.J. Breeding habits of Siberian Blue Robin (Larvivora cyane). Chi. J. Wildl. 1983, 10–11. [Google Scholar]
- Tamura, M.; Ueda, K. Breeding ecology of Siberian Blue Robin. Strix 2001, 19, 11–20. [Google Scholar]
- Haneda, K. Breeding biology of Japanese Robin, Erithacus akahige. Bull. Inst. Nat. Educ. 1976, 15, 9–19. [Google Scholar]
- Del Hoyo, J.; Elliott, A.; Christie, D.A. Handbook of the Birds of the World. In Cuckoo-Shrikes to Thrushes to Thrushes; Lynx Edicions: Barcelona, Spain, 2005; Volume 10, pp. 747–753. [Google Scholar]
- Mashru, A. Records of Indian Blue Robin Larvivora brunnea from Gujarat, India. Indian Birds 2014, 9, 160–161. [Google Scholar]
- Zhang, D.C.; He, T.L.; Feng, Y.F.; Hu, J.C.; Yang, B.; Li, S.Q. Preliminary camera-trapping survey on wild mammals and birds in the Wujiao Nature Reserve, Sichuan. Sichuan J. Zool. 2020, 39, 221–228. [Google Scholar]
- Chen, T.Y.; Chen, P.; Xu, P.; Zhang, M.Y.; Lu, C.H. Black-naped Monarch (Hypothymis azurea) and Indian Blue Robin (Larvivora brunnea) Observed in Yancheng, Jiangsu Province. Chi. J. Zool. 2020, 55, 683. [Google Scholar]
- Collar, N. Indian Blue Robin (Larvivora brunnea), Version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Kirwan, G.M.; Collar, N.; del Hoyo, J.; Christie, D.A.; Boesman, P.F.D. Ryukyu Robin (Larvivora komadori), Version 1.0. In Birds of the World; Keeney, B.K., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Zhang, Y.; Zhou, D.Q.; Zhao, Q.; Zhou, T.X.; Hyde, K.D. Diversity and ecological distribution of macrofungi in the Laojun Mountain region, southwestern China. Biodivers. Conserv. 2010, 19, 3545–3563. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L.J.; Zhou, T.X. Nidulariaceae distributed in Laojunshan mountainous area, Lijiang, northwest of Yunnan Province. J. Southwest For. Coll. 2006, 26, 62–66. [Google Scholar]
- Cao, R.J.; Yin, D.C.; Tian, K.; Xiao, D.R.; Li, Z.J.; Zhang, X.G.; Li, Z.H.; Zhang, Y. Response of radial growth of Abies georgei and Tsuga dumosa to climate change at upper distributional limits on Laojun Mountain, Lijiang, Yunnan, China. Acta Ecol. Sin. 2020, 40, 6067–6076. [Google Scholar]
- Li, Y.F.; He, J.; Li, C.H.; Xu, B.; Xie, X.J. A study of the relationship between the pollen in the surface sediments and the vegetation from the Laojun mountain of Lijiang, Yunnan Province, Southwest China. Acta Micropalaeontologica Sin. 2018, 35, 51–64. [Google Scholar]
- Li, W.D.; Stein, R.W.; Wang, X.W.; Luo, X. First account of the breeding biology of Brown Parrotbill (Cholornis unicolor, Aves: Sylviidae) in Gaoligongshan National Nature Reserve, Yunnan, China. Wilson J. Ornithol. 2019, 131, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Song, Y.F.; Zhong, Y.X.; Chen, C.W.; Zhao, Y.H.; Zeng, D.; Wu, Y.R.; Ding, P. A dataset on the life-history and ecological traits of Chinese birds. Biodivers. Sci. 2021, 29, 1149–1153. [Google Scholar] [CrossRef]
- Soler, J.J.; Moreno, J. Evolution of sexual dichromatism in relation to nesting habits in European passerines: A test of Wallace’s hypothesis. J. Evol. Biol. 2012, 25, 1614–1622. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake, K.B.; Weston, M.A.; Nimmo, D.G.; Maguire, G.S.; Endler, J.A.; Küpper, C. The bright incubate at night: Sexual dichromatism and adaptive incubation division in an open-nesting shorebird. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143026. [Google Scholar] [CrossRef] [PubMed]
- Sofaer, H.R.; Sillett, T.S.; Peluc, S.I.; Morrison, S.A.; Ghalambor, C.K. Differential effects of food availability and nest predation risk on avian reproductive strategies. Behav. Ecol. 2013, 24, 698–707. [Google Scholar] [CrossRef]
- Liang, D.; Gao, G.; Pagani-Núñez, E.; Pang, H.; Liu, Y.; Luo, X.; Robinson, S.K. Incubation behaviour of a high-altitude species: The Fire-tailed Sunbird Aethopyga ignicauda. Bird Study 2018, 65, 261–265. [Google Scholar] [CrossRef]
- Badyaev, A.V. Avian life history variation along altitudinal gradients: An example with cardueline finches. Oecologia 1997, 111, 365–374. [Google Scholar] [CrossRef]
- Badyaev, A.V.; Ghalambor, C.K. Evolution of life histories along elevational gradients: Trade-off between parental care and fecundity. Ecology 2001, 82, 2948–2960. [Google Scholar] [CrossRef]
- Collar, N. Rufous-tailed Robin (Larvivora sibilans), Version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Collar, N. Rufous-headed Robin (Larvivora ruficeps), Version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Covas, R. Evolution of reproductive life histories in island birds worldwide. Proc. R. Soc. B Biol. Sci. 2012, 279, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Roff, D.A. The Evolution of Life Histories; Chapman and Hall: New York, NY, USA, 1992. [Google Scholar]
- Cody, M.L. A general theory of clutch size. Evolution 1966, 20, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Christians, J.K. Avian egg size: Variation within species and inflexibility within individuals. Biol. Rev. 2002, 77, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Owens, I.P.; Bennett, P.M. Ancient ecological diversification explains life-history variation among living birds. Philos. Trans. R. Soc. Biol. 1995, 261, 227–232. [Google Scholar]
- Jetz, W.; Sekercioglu, C.H.; Böhning-Gaese, K. The worldwide variation in avian clutch size across species and space. PLoS Biol. 2008, 6, e303. [Google Scholar] [CrossRef]
- Decker, K.L.; Conway, C.J.; Fontaine, J.J. Nest predation, food, and female age explain seasonal declines in clutch size. Evol. Ecol. 2012, 26, 683–699. [Google Scholar] [CrossRef]
- Kirwan, G.M.; Collar, N.; del Hoyo, J.; Christie, D.A.; Boesman, P.F.D. Okinawa Robin (Larvivora namiyei), Version 1.0. In Birds of the World; Keeney, B.K., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Kirwan, G.M.; Collar, N.; del Hoyo, J.; Christie, D.A.; Boesman, P.F.D. Izu Robin (Larvivora tanensis), Version 1.0. In Birds of the World; Kirwan, G.M., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar]
- Basnet, D.; Yang, J.M.; Dorji, T.; Xiao, Q.L.; Kumari, A.L.; Yue, M.W.; Wu, N.; Wei, Y.T.; Kamala, G.; Li, R.J.; et al. Bird photography tourism, sustainable livelihoods, and biodiversity conservation: A case study from China. Mt. Res. Dev. 2021, 41, D1–D9. [Google Scholar] [CrossRef]
- Harrison, T.J.; Smith, J.A.; Martin, G.R.; Chamberlain, D.E.; Bearhop, S.; Robb, G.N.; Reynolds, S.J. Does food supplementation really enhance productivity of breeding birds? Oecologia 2010, 164, 311–320. [Google Scholar] [CrossRef]
- Oro, D.; Genovart, M.; Tavecchia, G.; Fowler, M.S.; Martínez-Abraín, A. Ecological and evolutionary implications of food subsidies from humans. Ecol. Lett. 2013, 16, 1501–1514. [Google Scholar] [CrossRef]
- Robb, G.N.; McDonald, R.A.; Chamberlain, D.E.; Bearhop, S. Food for thought: Supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 2008, 6, 476–484. [Google Scholar] [CrossRef]
- Plummer, K.E.; Siriwardena, G.M.; Conway, G.J.; Risely, K.; Toms, M.P. Is supplementary feeding in gardens a driver of evolutionary change in a migratory bird species? Glob. Change Biol. 2015, 21, 4353–4363. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, J.A.; Stanley, M.C.; Jones, D.N.; Beggs, J.R. Experimental feeding regime influences urban bird disease dynamics. J. Avian. Biol. 2017, 48, 700–713. [Google Scholar] [CrossRef]
- Hanmer, H.J.; Thomas, R.L.; Fellowes, M.D.E. Provision of supplementary food for wild birds may increase the risk of local nest predation. Ibis 2017, 159, 158–167. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Galbraith, J.A.; Smith, J.A.; Jones, D.N. Garden bird feeding: Insights and prospects from a north-south comparison of this global urban phenomenon. Front. Ecol. Evol. 2017, 5, 24. [Google Scholar] [CrossRef]
- Sorensen, A.; van Beest, F.M.; Brook, R.K. Impacts of wildlife baiting and supplemental feeding on infectious disease transmission risk: A synthesis of knowledge. Prev. Vet. Med. 2014, 113, 356–363. [Google Scholar] [CrossRef]
- Murray, M.H.; Becker, D.J.; Hall, R.J.; Hernandez, S.M. Wildlife health and supplemental feeding: A review and management recommendations. Biol. Conserv. 2016, 204, 163–174. [Google Scholar] [CrossRef]
- Strandin, T.; Babayan, S.A.; Forbes, K.M. Reviewing the effects of food provisioning on wildlife immunity. Philos Trans. R. Soc. Biol. 2018, 373, 20170088. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, J.; Fan, S.; Luo, X. First Account of the Breeding Biology of Indian Blue Robin (Larvivora brunnea) in Southwest China. Animals 2024, 14, 39. https://doi.org/10.3390/ani14010039
Nie J, Fan S, Luo X. First Account of the Breeding Biology of Indian Blue Robin (Larvivora brunnea) in Southwest China. Animals. 2024; 14(1):39. https://doi.org/10.3390/ani14010039
Chicago/Turabian StyleNie, Jun, Shixiang Fan, and Xu Luo. 2024. "First Account of the Breeding Biology of Indian Blue Robin (Larvivora brunnea) in Southwest China" Animals 14, no. 1: 39. https://doi.org/10.3390/ani14010039
APA StyleNie, J., Fan, S., & Luo, X. (2024). First Account of the Breeding Biology of Indian Blue Robin (Larvivora brunnea) in Southwest China. Animals, 14(1), 39. https://doi.org/10.3390/ani14010039