
The Genetic Selection of HSPD1 and HSPE1 Reduce Inflammation of Liver and Spleen While Restraining the Growth and Development of Skeletal Muscle in Wuzhishan Pigs
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Data Sets
2.2. Resequencing and Variant Calling
2.3. Population Genetic Differentiation
2.4. Population Structure
2.5. History Demographic Estimation
2.6. Structural Variation Calling
2.7. RNA-Seq Analysis
2.8. Functional Enrichment
2.9. RT-qPCR
3. Results
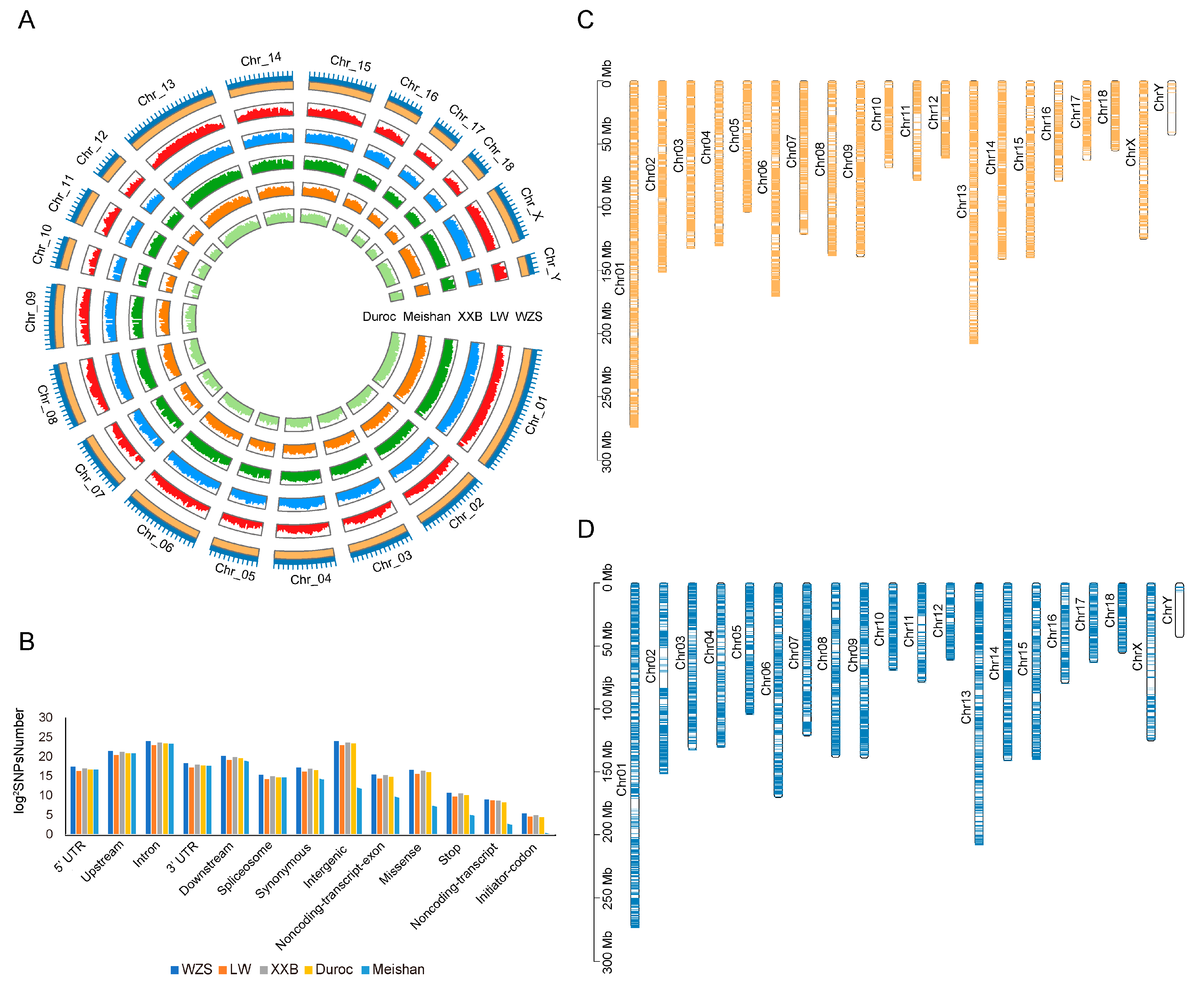
3.1. Genomic Variant Map
3.2. Genetic Relationship and Population Structure
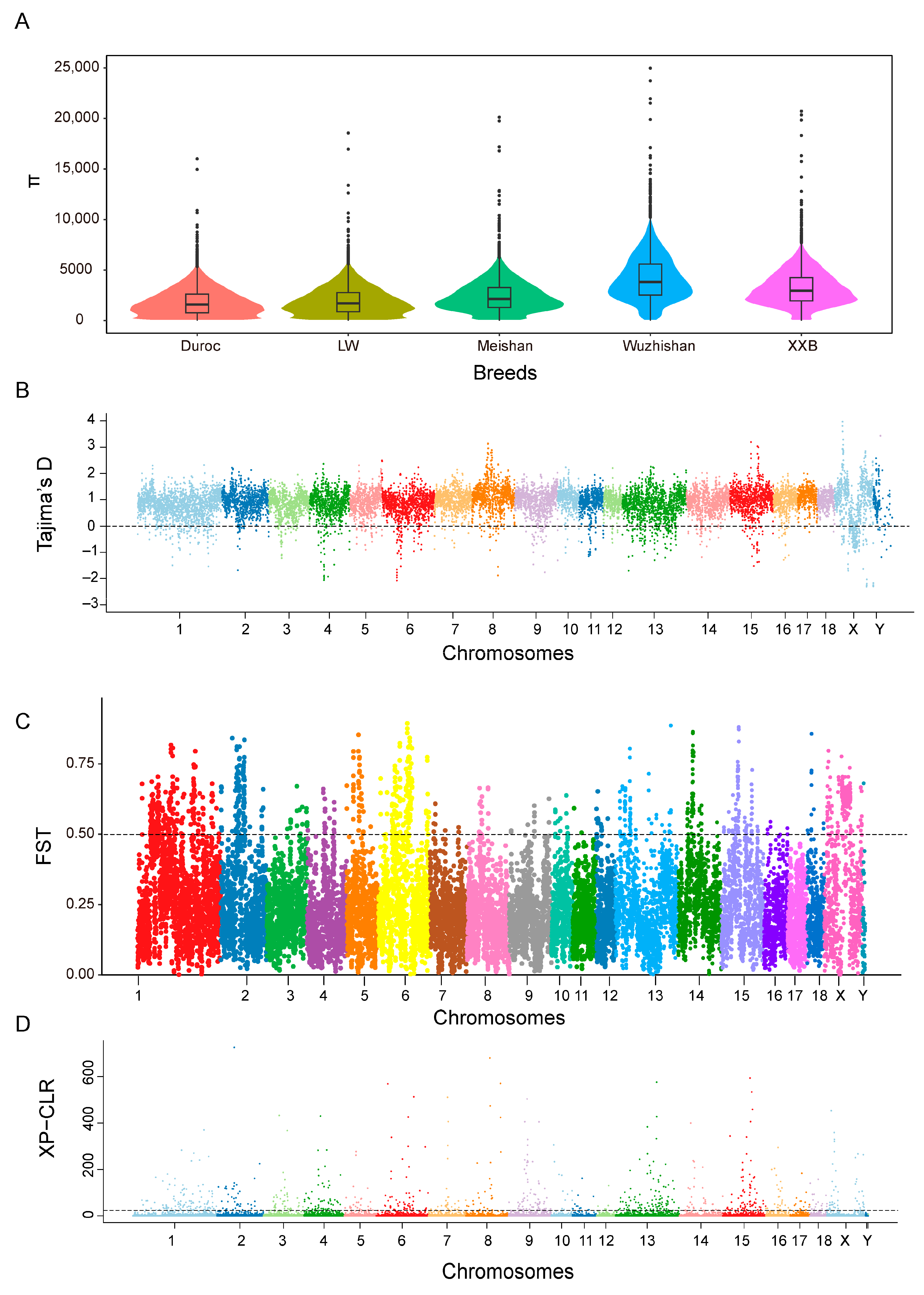
3.3. Effective Population Size
3.4. Genetic Diversity
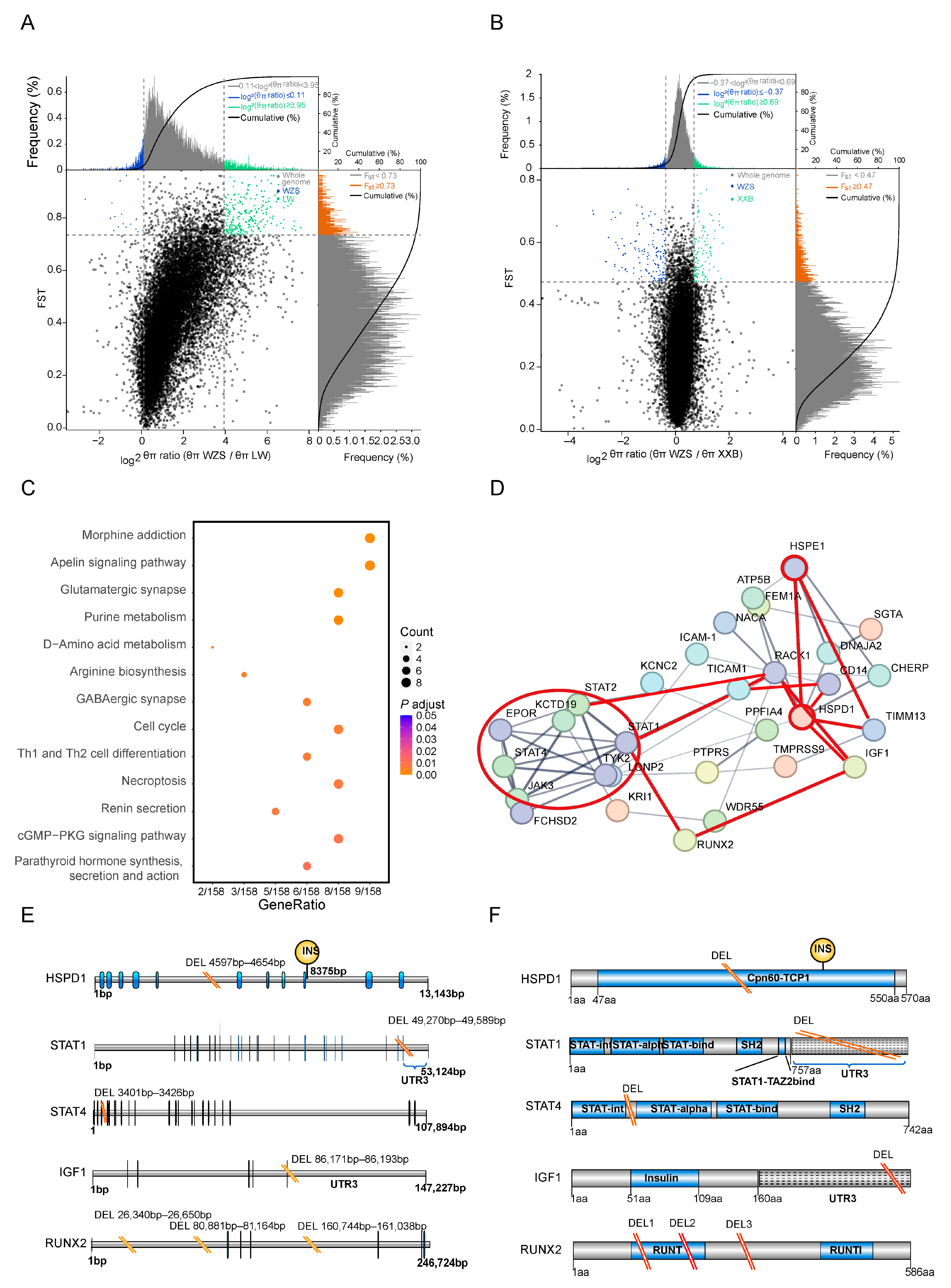
3.5. Functional Annotation of the Most 5% Distant Genes
3.6. SV Detection in Genes in the PPI Network
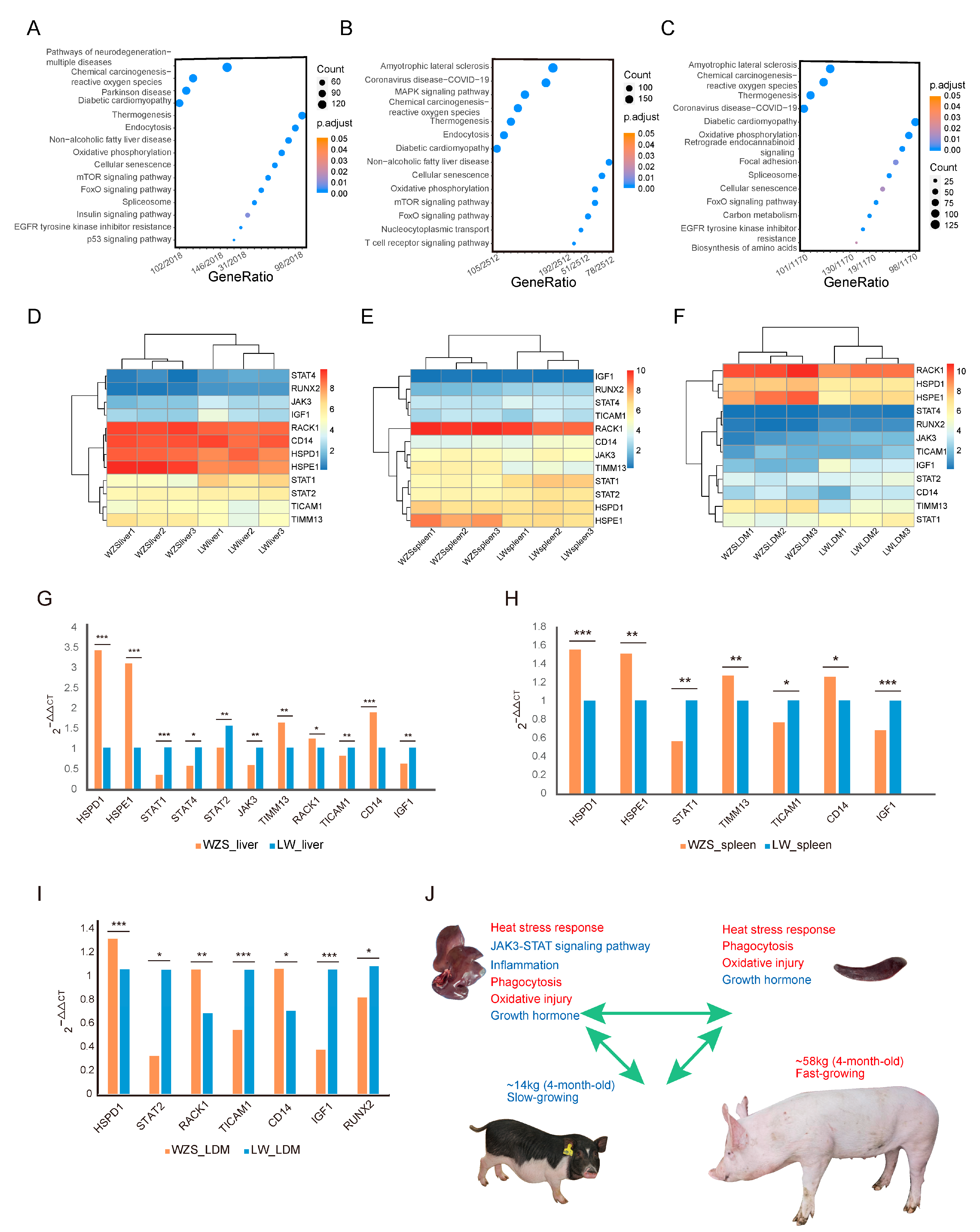
3.7. Functional Analysis of Differentially Expressed Genes between WZS and LW Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeder, M.A. The origins of agriculture in the Near East. Curr. Anthropol. 2011, 52, S221–S235. [Google Scholar] [CrossRef]
- Hofmanová, Z.; Kreutzer, S.; Hellenthal, G.; Sell, C.; Diekmann, Y.; Díez-Del-Molino, D.; van Dorp, L.; López, S.; Kousathanas, A.; Link, V.; et al. Early farmers from across Europe directly descended from Neolithic Aegeans. Proc. Natl. Acad. Sci. USA 2016, 113, 6886–6891. [Google Scholar] [CrossRef] [PubMed]
- Font-I-Furnols, M.; Guerrero, L. Consumer preference, behavior and perception about meat and meat products: An overview. Meat Sci. 2014, 98, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Pabst, R. The pig as a model for immunology research. Cell Tissue Res. 2020, 380, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Donegà, M.; Fjordbakk, C.T.; Kirk, J.; Sokal, D.M.; Gupta, I.; Hunsberger, G.E.; Crawford, A.; Cook, S.; Viscasillas, J.; Stathopoulou, T.R.; et al. Human-relevant near-organ neuromodulation of the immune system via the splenic nerve. Proc. Natl. Acad. Sci. USA 2021, 118, e2025428118. [Google Scholar] [CrossRef] [PubMed]
- Thrall, K.D.; Mahendra, S.; Jackson, M.K. A gottingen minipig model of radiation-induced coagulopathy. Int. J. Radiat. Biol. 2021, 97 (Suppl. S1), S63–S72. [Google Scholar] [CrossRef] [PubMed]
- Mahan, B.; Tacail, T.; Lewis, J.; Elliott, T.; Habekost, M.; Turner, S.; Chung, R.; Moynier, F. Exploring the K isotope composition of Göttingen minipig brain regions, and implications for Alzheimer’s disease. Metallomics 2022, 14, mfac090. [Google Scholar] [CrossRef]
- Guo, Q.; Zhu, H.Y.; Jin, L.; Gao, Q.S.; Kang, J.D.; Cui, C.D.; Yin, X.J. Production of cloned Wuzhishan miniature pigs and application for alloxan toxicity test. Anim. Biotechnol. 2015, 26, 292–297. [Google Scholar] [CrossRef]
- Jiang, X.; Qian, T.; Linn, T.; Cao, L.; Xiang, G.; Wang, Y.; Peng, H.; Xue, P.; Zhang, L.; Chen, D.; et al. Islet isolation and purification from inbred Wuzhishan miniature pigs. Xenotransplantation 2012, 19, 159–165. [Google Scholar] [CrossRef]
- Lee, J.; Park, N.; Lee, D.; Kim, J. Evolutionary and functional analysis of Korean native Pig using single nucleotide polymorphisms. Mol. Cell 2020, 43, 728–738. [Google Scholar]
- Ma, H.; Jiang, J.; He, J.; Liu, H.; Han, L.; Gong, Y.; Li, B.; Yu, Z.; Tang, S.; Zhang, Y.; et al. Long-read assembly of the Chinese indigenous Ningxiang pig genome and identification of genetic variations in fat metabolism among different breeds. Mol. Ecol. Resour. 2022, 22, 1508–1520. [Google Scholar] [CrossRef]
- Zhong, Z.; Wang, Z.; Xie, X.; Tian, S.; Wang, F.; Wang, Q.; Ni, S.; Pan, Y.; Xiao, Q. Evaluation of the genetic diversity, population structure and selection signatures of three native chinese pig populations. Animals 2023, 13, 2010. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Kong, C.; Chen, Y.; Zheng, X.; Zhou, R.; Zhang, X.; Wu, X.; Zhang, W.; Ding, Y.; Yin, Z. Copy number variation analysis revealed the evolutionary difference between Chinese indigenous pigs and Asian wild boars. Genes 2023, 14, 472. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat shock proteins as immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef] [PubMed]
- Liyanagamage, D.S.N.K.; Martinus, R.D. Role of mitochondrial stress protein HSPD1 in diabetes-induced neuroinflammation. Mediat. inflamm. 2020, 2020, 8073516. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Srivastava, A.; Jagdale, P.; Ayanur, A.; Khanna, V.K. Lambda-cyhalothrin enhances inflammation in nigrostriatal region in rats: Regulatory role of NF-κβ and JAK-STAT signaling. Neurotoxicology 2023, 96, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Fang, L.; Dong, Y.; Yang, J.; Chen, X.; Zhang, N.; Zhu, Y.; Huang, T. Mitochondrial quality control protects photoreceptors against oxidative stress in the H2O2-induced models of retinal degeneration diseases. Cell Death Dis. 2021, 12, 413. [Google Scholar] [CrossRef] [PubMed]
- Yeung, N.; Murata, D.; Iijima, M.; Sesaki, H. Role of human HSPE1 for OPA1 processing independent of HSPD1. iScience 2023, 26, 106067. [Google Scholar] [CrossRef]
- Cannon, A.S.; Holloman, B.L.; Wilson, K.; Miranda, K.; Dopkins, N.; Nagarkatti, P.; Nagarkatti, M. AhR activation leads to attenuation of murine autoimmune hepatitis: Single-cell RNA-Seq analysis reveals unique immune cell phenotypes and gene expression changes in the liver. Front. Immunol. 2022, 13, 899609. [Google Scholar] [CrossRef]
- Verhoeven, Y.; Tilborghs, S.; Jacobs, J.; De Waele, J.; Quatannens, D.; Deben, C.; Prenen, H.; Pauwels, P.; Trinh, X.B.; Wouters, A.; et al. The potential and controversy of targeting STAT family members in cancer. Semin. Cancer Biol. 2020, 60, 41–56. [Google Scholar] [CrossRef]
- Montero, P.; Milara, J.; Roger, I.; Cortijo, J. Role of JAK/STAT in interstitial lung diseases; molecular and cellular mechanisms. Int. J. Mol. Sci. 2021, 22, 6211. [Google Scholar] [CrossRef]
- Svendsen, O. The minipig in toxicology. Exp. Toxicol. Pathol. 2006, 57, 335–339. [Google Scholar] [CrossRef]
- Liu, F.; Zhao, W.; Le, H.H.; Cottrell, J.J.; Green, M.P.; Leury, B.J.; Dunshea, F.R.; Bell, A.W. Review: What have we learned about the effects of heat stress on the pig industry? Animal 2022, 16 (Suppl. S2), 100349. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Yu, Y.; Feng, W.; Du, H.; Yu, J.; Kang, H.; Zheng, X.; Wang, Z.; Liu, G.E.; Ernst, C.W.; et al. Evidence of evolutionary history and selective sweeps in the genome of Meishan pig reveals its genetic and phenotypic characterization. Gigascience 2018, 7, giy058. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A mapreduce framework for analyzing nextgeneration DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Lv, F.H.; Cao, Y.H.; Liu, G.J.; Luo, L.Y.; Lu, R.; Liu, M.J.; Li, W.R.; Zhou, P.; Wang, X.H.; Shen, M.; et al. Whole-genome resequencing of worldwide wild and domestic sheep elucidates genetic diversity, introgression, and agronomically important loci. Mol. Biol. Evol. 2022, 39, msab353. [Google Scholar] [CrossRef]
- Cingolani, P. Variant Annotation and Functional Prediction: SnpEff. Methods Mol. Biol. 2022, 2493, 289–314. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population-structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [PubMed]
- Zhang, C.; Dong, S.S.; Xu, J.Y.; He, W.M.; Yang, T.L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Yao, C.T.; Hung, C.M. Domestication obscures genomic estimates of population history. Mol. Ecol. 2022, 31, 752–766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, Q.; Ai, H.; Huang, L. Revisiting the evolutionary history of pigs via de novo mutation rate estimation in a three-generation pedigree. Genom. Proteom. Bioinf. 2022, 6, 1040–1052. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Estavoyer, M.; François, O. Theoretical analysis of principal components in an umbrella model of intraspecific evolution. Theor. Popul. Biol. 2022, 148, 11–21. [Google Scholar] [CrossRef]
- Bruger, E.L.; Marx, C.J. A decade of genome sequencing has revolutionized studies of experimental evolution. Curr. Opin. Microbiol. 2018, 45, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, Z.; Zhao, Q.; Du, H.; Yu, J.; Wang, H.; Liu, X.; Liu, H.; Jing, X.; Yang, H.; et al. Population genetic structure and selection signature analysis of Beijing Black Pig. Front. Genet. 2022, 13, 860669. [Google Scholar] [CrossRef]
- Wei, G.; Li, X.; Wang, D.; Zhao, B.; Shi, Y.; Huang, A. Discovery of specific antioxidant peptide from Chinese Dahe black pig and hybrid pig dry-cured hams based on peptidomics strategy. Food Res. Int. 2023, 166, 112610. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, M.; Wang, Y.; Wu, X.; Zhang, X.; Ding, Y.; Yin, Z. Genomic analysis reveals selection signatures of the Wannan Black pig during domestication and breeding. Asian-Australas. J. Anim. Sci. 2020, 33, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Marino Gammazza, A.; Palumbo Piccionello, A.; Campanella, C.; Pace, A.; Conway de Macario, E.; Macario, A.J. HSPD1 chaperonopathies and chaperonotherapy: Targets and agents. Expert Opin. Ther. Targets 2014, 18, 185–208. [Google Scholar] [CrossRef]
- Domínguez-Horta, M.D.C.; Serrano-Díaz, A.; Hernández-Cedeño, M.; Martínez-Donato, G.; Guillén-Nieto, G. A peptide derived from HSP60 reduces proinflammatory cytokines and soluble mediators: A therapeutic approach to inflammation. Front. Immunol. 2023, 14, 1162739. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.; Le, T.T.; Dobbin, C.A.; Banovic, T.; Howard, C.B.; Flores Fde, M.; Vanags, D.; Naylor, D.J.; Hill, G.R.; Suhrbier, A. Heat shock protein 10 inhibits lipopolysaccharide-induced inflammatory mediator production. J. Biol. Chem. 2005, 280, 4037–4047. [Google Scholar] [CrossRef]
- Zai, Z.; Xu, Y.; Qian, X.; Li, Z.; Ou, Z.; Zhang, T.; Wang, L.; Ling, Y.; Peng, X.; Zhang, Y. Estrogen antagonizes ASIC1a-induced chondrocyte mitochondrial stress in rheumatoid arthritis. J. Transl. Med. 2022, 20, 561. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Wang, X.; Fu, C.; Zhao, F.; Zou, L.; Wu, K.; Chen, W.; Li, Z.; Fan, J.; et al. The regulation of cell homeostasis and antiviral innate immunity by autophagy during classical swine fever virus infection. Emerg. Microbes Infect. 2023, 12, 2164217. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhang, Y.J. Antagonizing cytokine-mediated JAK-STAT signaling by porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.P.; Maclaren, D.P.; Cable, N.T.; Campbell, I.T.; Evans, L.; Kayani, A.C.; McArdle, A.; Drust, B. Trained men display increased basal heat shock protein content of skeletal muscle. Med. Sci. Sports Exerc. 2008, 40, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Marino Gammazza, A.; Macaluso, F.; Di Felice, V.; Cappello, F.; Barone, R. HSPD1 in skeletal muscle fiber biogenesis and homeostasis: From physical exercise to skeletal muscle pathology. Cells 2018, 7, 224. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yin, S.; He, Y.; Tang, J.; Pu, J.; Jia, G.; Liu, G.; Tian, G.; Chen, X.; Cai, J.; et al. Hydroxy-Selenomethionine mitigated chronic heat stress-induced porcine splenic damage via activation of Nrf2/Keap1 signal and suppression of NFκb and STAT signal. Int. J. Mol. Sci. 2023, 24, 6461. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Yan, P.; Song, R.; Liu, F.; Tian, Q. HOXC13-driven TIMM13 overexpression promotes osteosarcoma cell growth. Cell Death Dis. 2023, 14, 398. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Q.; Zhao, Z.; Fan, J.; Qin, L.; Alexander, D.B.; Tsuda, H.; Zhao, D.; Xu, J. Surfactant proteins A/D-CD14 on alveolar macrophages is a common pathway associated with phagocytosis of nanomaterials and cytokine production. Front. Immunol. 2021, 12, 758941. [Google Scholar] [CrossRef] [PubMed]
- Kamanga-Sollo, E.; Pampusch, M.S.; White, M.E.; Hathaway, M.R.; Dayton, W.R. Effects of heat stress on proliferation, protein turnover, and abundance of heat shock protein messenger ribonucleic acid in cultured porcine muscle satellite cells. J. Anim. Sci. 2011, 89, 3473–3480. [Google Scholar] [CrossRef]
- Cervantes, M.; Cota, M.; Arce, N.; Castillo, G.; Avelar, E.; Espinoza, S.; Morales, A. Effect of heat stress on performance and expression of selected amino acid and glucose transporters, HSP90, leptin and ghrelin in growing pigs. J. Therm. Biol. 2016, 59, 69–76. [Google Scholar] [CrossRef]
- Kim, M.; Roura, E.; Choi, Y.; Kim, J. Transcriptomic Analysis of the Porcine Gut in Response to Heat Stress and Dietary Soluble Fiber from Beet Pulp. Genes 2022, 13, 1456. [Google Scholar] [CrossRef]
- Zou, Y.; Hu, X.M.; Zhang, T.; Wei, H.K.; Zhou, Y.F.; Zhou, Z.X.; Peng, J. Effects of dietary oregano essential oil and vitamin E supplementation on meat quality, stress response and intestinal morphology in pigs following transport stress. J. Vet. Med. Sci. 2017, 79, 328–335. [Google Scholar] [CrossRef]
- Gurevich, E.; Segev, Y.; Landau, D. Growth hormone and IGF1 actions in kidney development and function. Cells 2021, 10, 3371. [Google Scholar] [CrossRef]
- Komori, T. Whole aspect of Runx2 functions in skeletal development. Int. J. Mol. Sci. 2022, 23, 5776. [Google Scholar] [CrossRef]
- Reverter, A.; Ballester, M.; Alexandre, P.A.; Mármol-Sánchez, E.; Dalmau, A.; Quintanilla, R.; Ramayo-Caldas, Y. A gene co-association network regulating gut microbial communities in a Duroc pig population. Microbiome 2021, 9, 52. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Wang, F.; Sun, R.; Zheng, X.; Liu, Y.; Lin, Y.; Hong, L.; Huang, X.; Chao, Z. The Genetic Selection of HSPD1 and HSPE1 Reduce Inflammation of Liver and Spleen While Restraining the Growth and Development of Skeletal Muscle in Wuzhishan Pigs. Animals 2024, 14, 174. https://doi.org/10.3390/ani14010174
Ren Y, Wang F, Sun R, Zheng X, Liu Y, Lin Y, Hong L, Huang X, Chao Z. The Genetic Selection of HSPD1 and HSPE1 Reduce Inflammation of Liver and Spleen While Restraining the Growth and Development of Skeletal Muscle in Wuzhishan Pigs. Animals. 2024; 14(1):174. https://doi.org/10.3390/ani14010174
Chicago/Turabian StyleRen, Yuwei, Feng Wang, Ruiping Sun, Xinli Zheng, Yuanyuan Liu, Yanning Lin, Lingling Hong, Xiaoxian Huang, and Zhe Chao. 2024. "The Genetic Selection of HSPD1 and HSPE1 Reduce Inflammation of Liver and Spleen While Restraining the Growth and Development of Skeletal Muscle in Wuzhishan Pigs" Animals 14, no. 1: 174. https://doi.org/10.3390/ani14010174
APA StyleRen, Y., Wang, F., Sun, R., Zheng, X., Liu, Y., Lin, Y., Hong, L., Huang, X., & Chao, Z. (2024). The Genetic Selection of HSPD1 and HSPE1 Reduce Inflammation of Liver and Spleen While Restraining the Growth and Development of Skeletal Muscle in Wuzhishan Pigs. Animals, 14(1), 174. https://doi.org/10.3390/ani14010174