
Long-Term Seed Dispersal within an Asymmetric Lizard-Plant Interaction

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
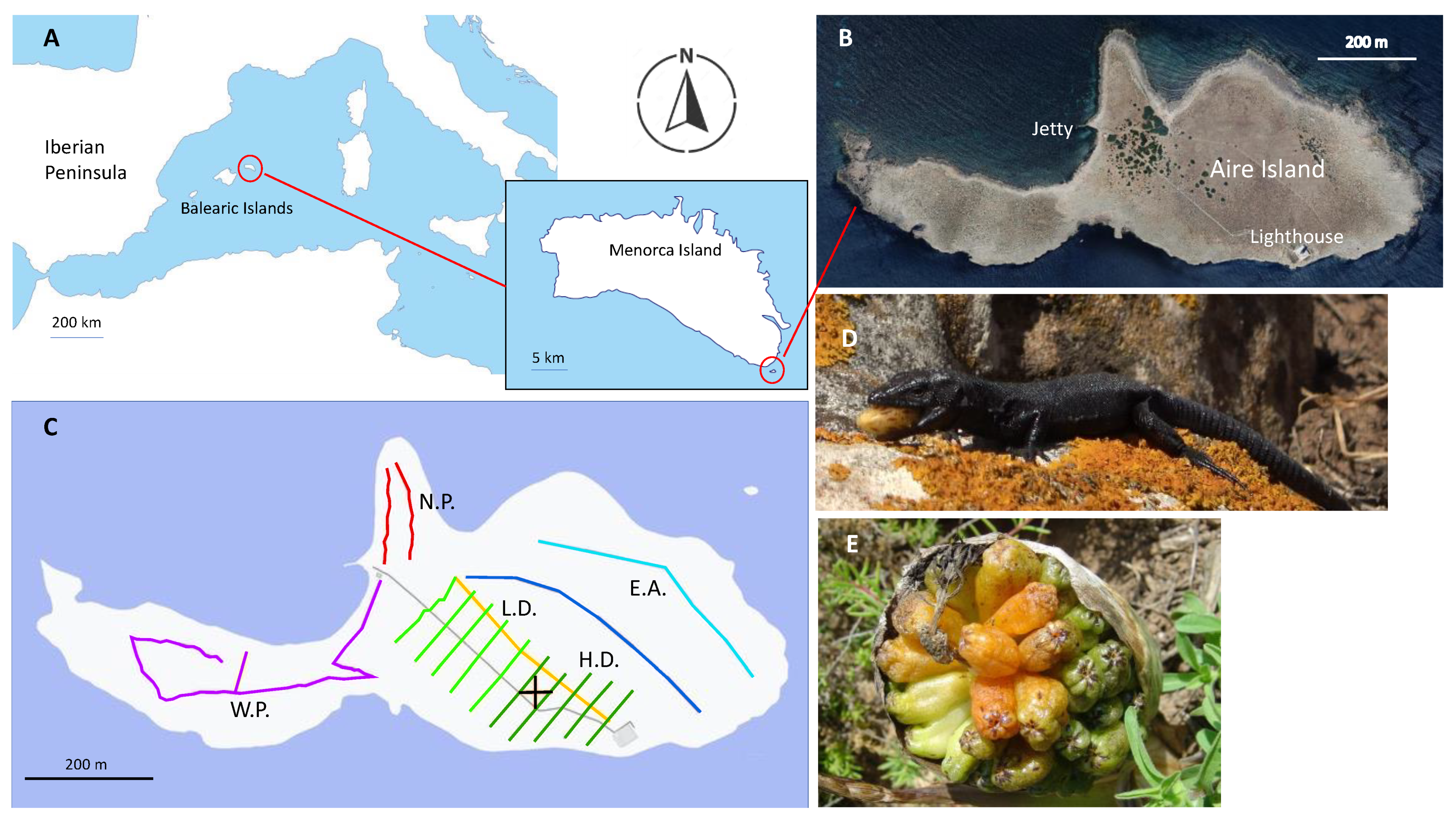
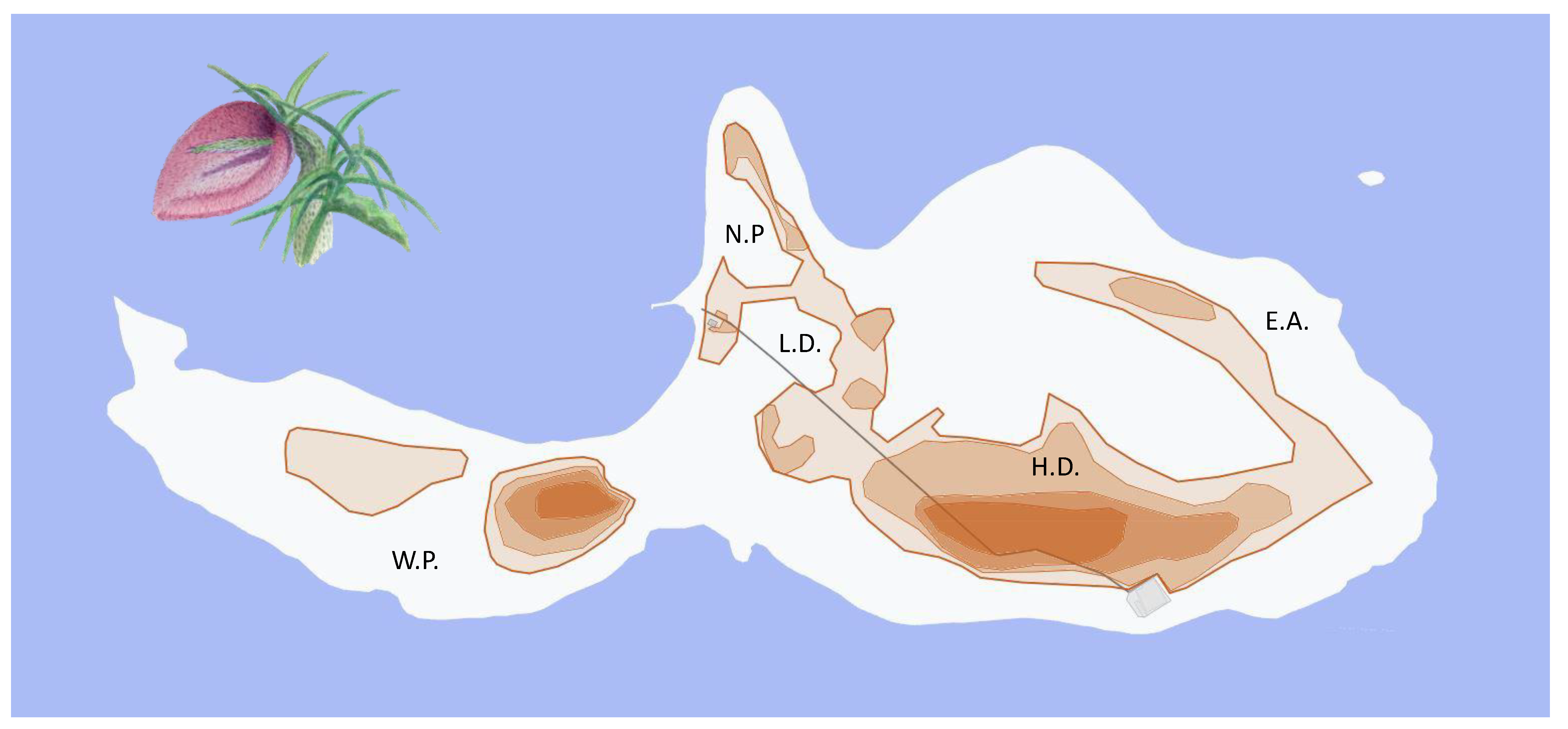
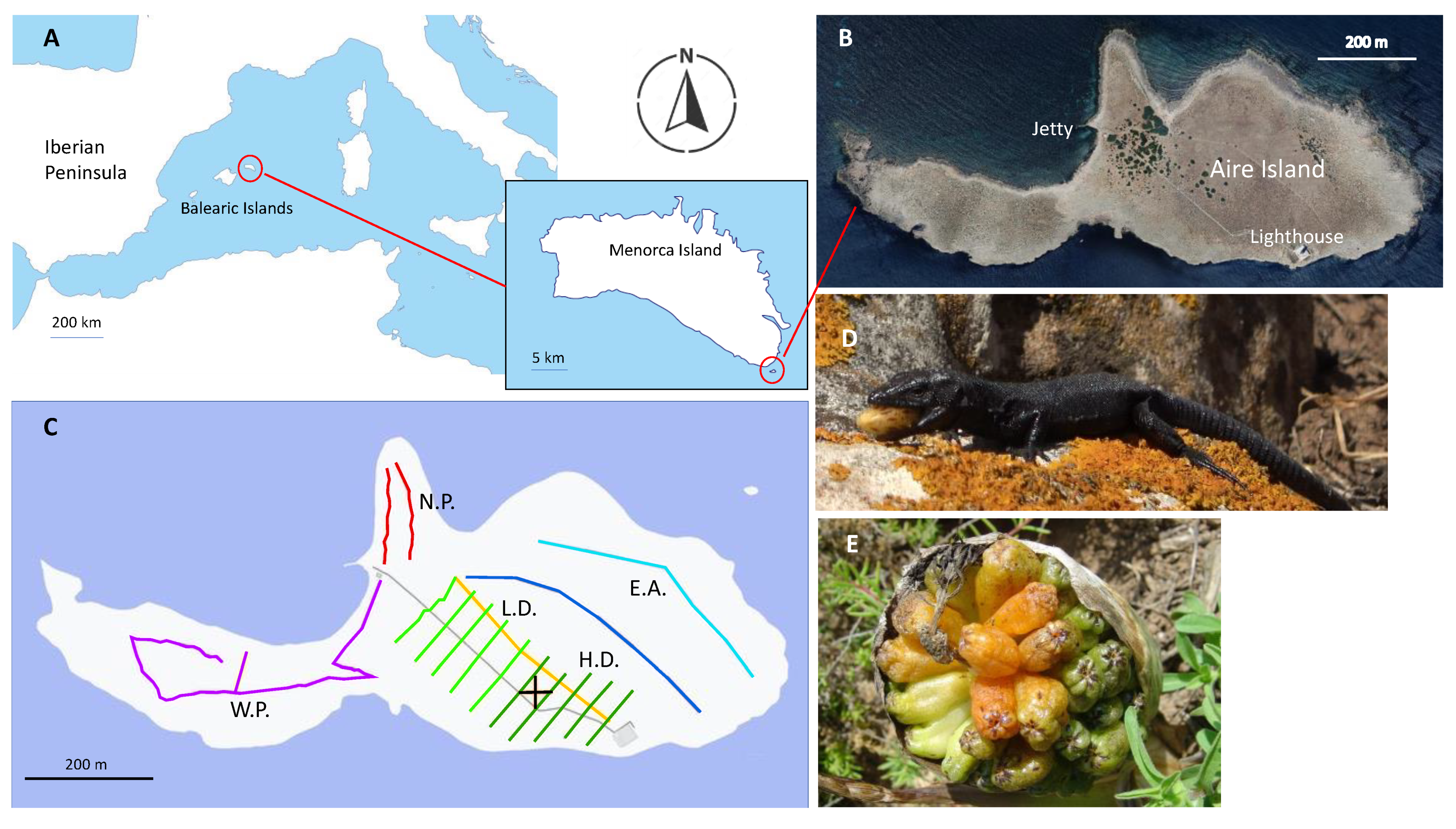
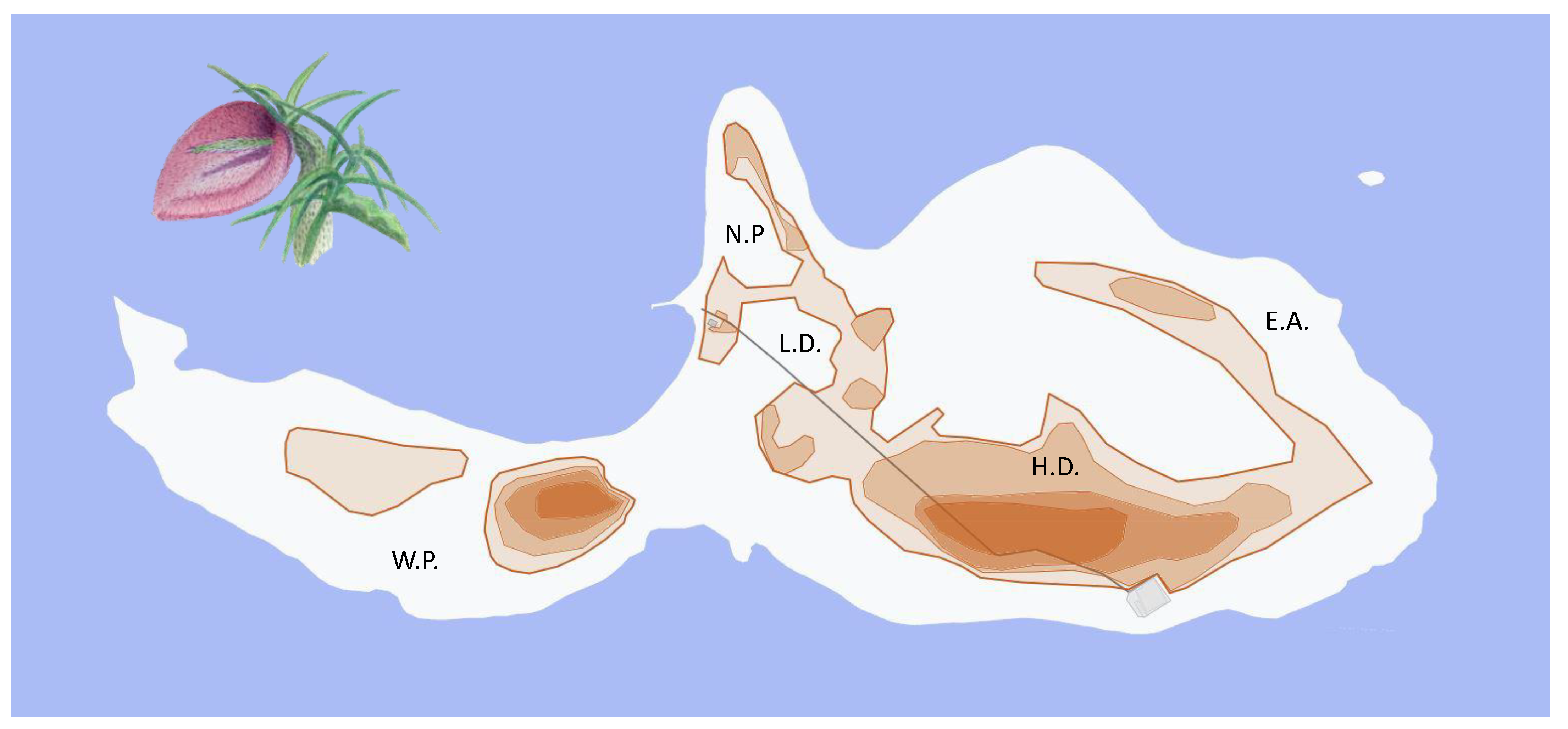
2.1. Study Period and Area
2.2. Study System
2.2.1. The Lizard
2.2.2. The Plant
2.3. Distribution and Abundance of Plants and Lizards
2.4. Seed Dispersal
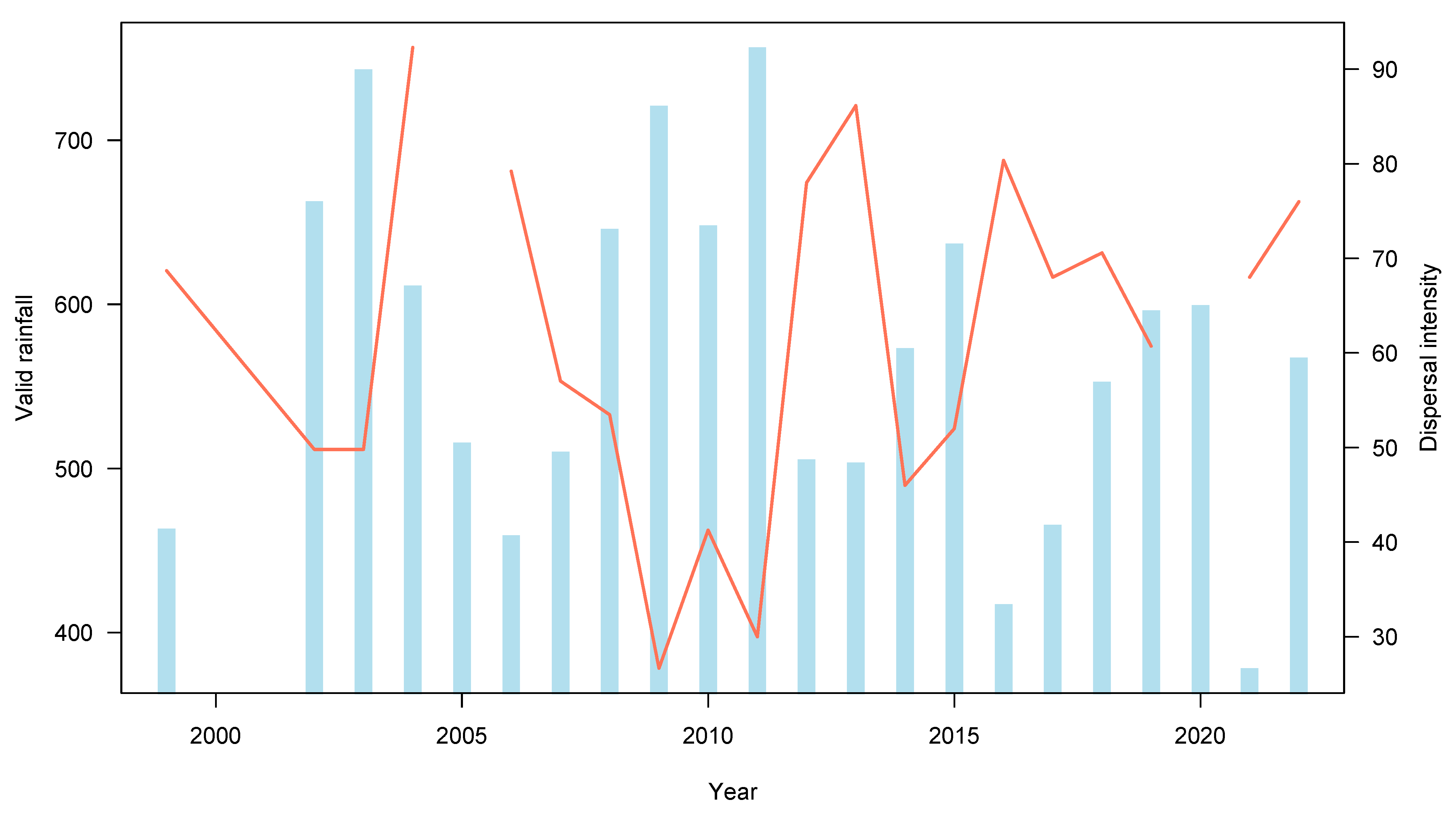
2.5. Climatic Information
2.6. Statistical Analyses
3. Results
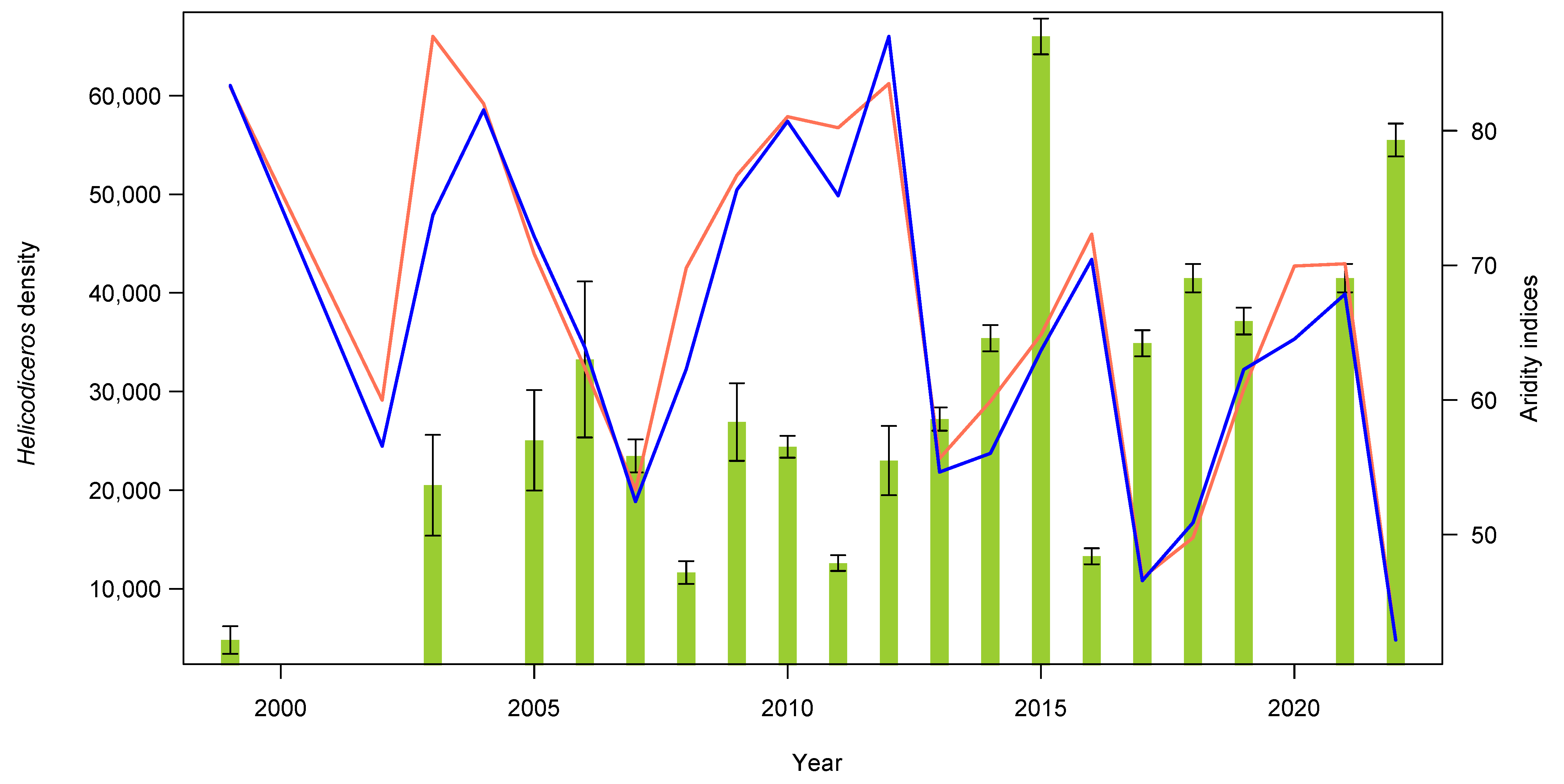
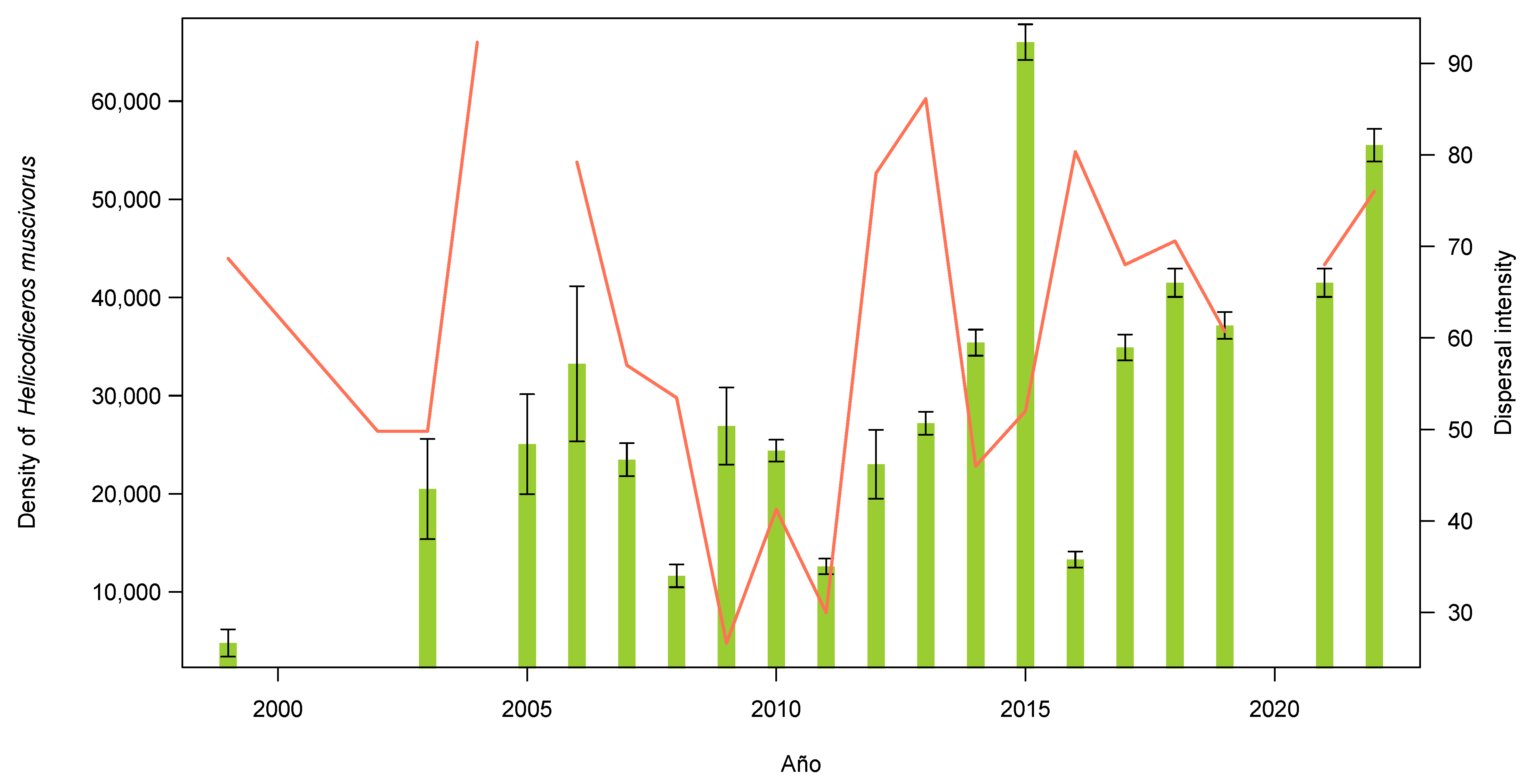
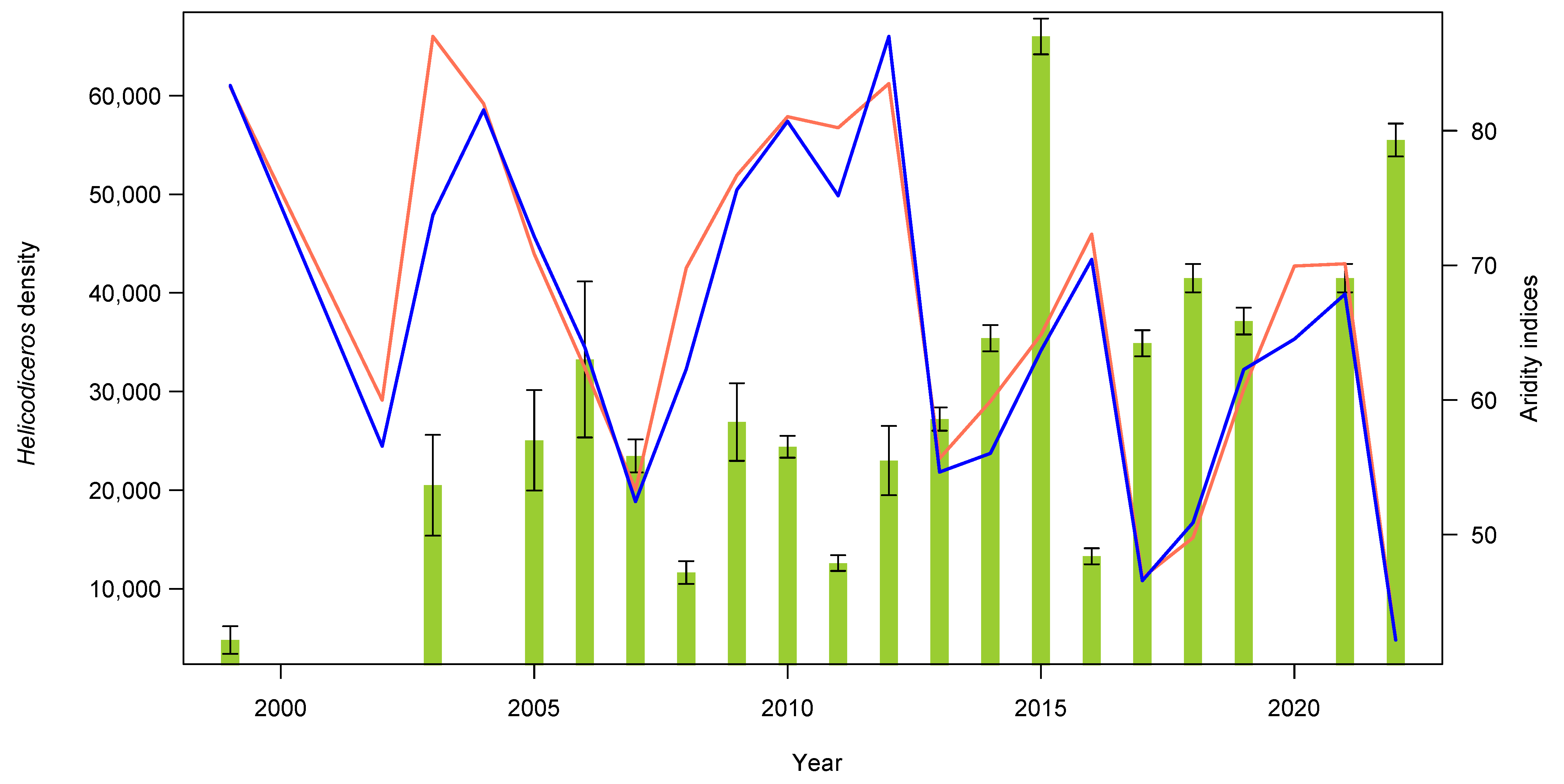
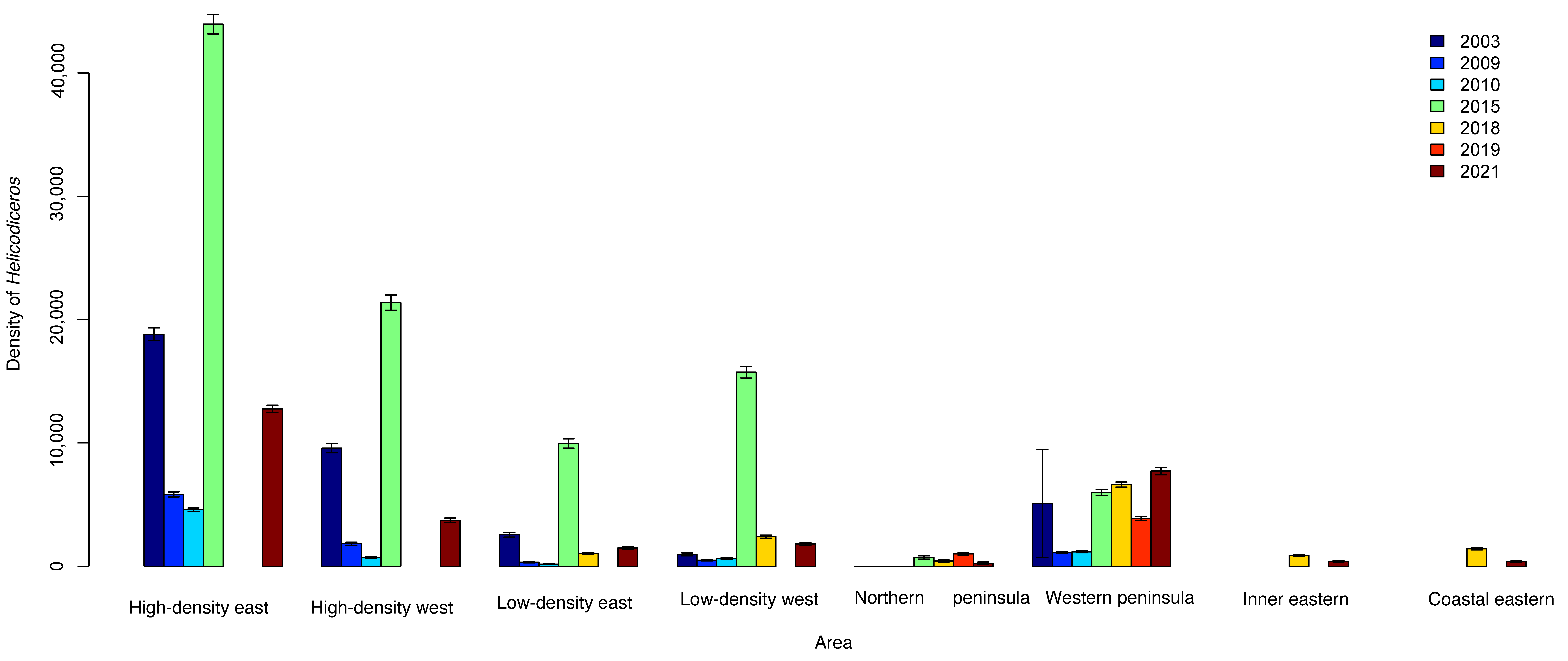
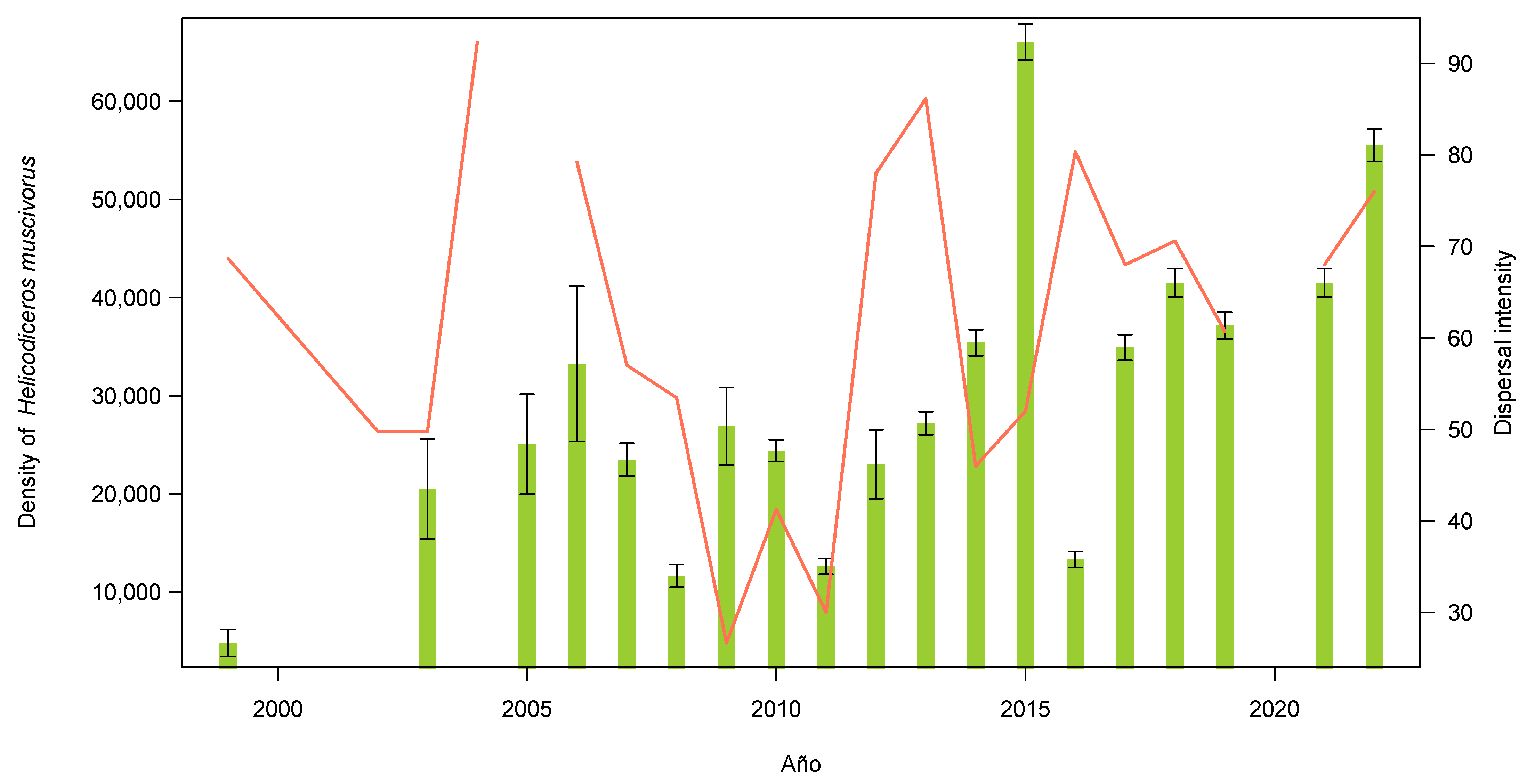
3.1. Plant and Lizard Densities
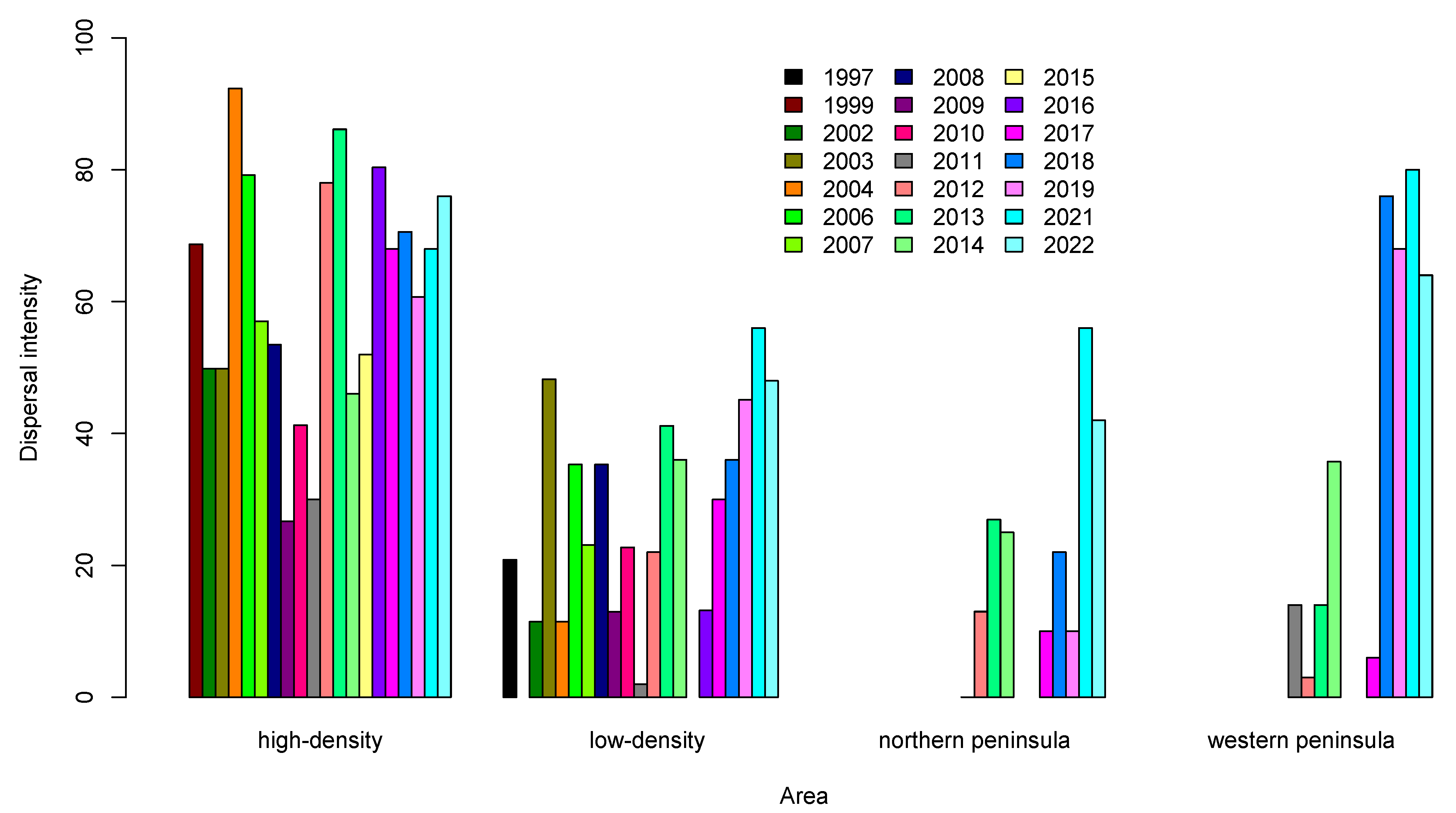
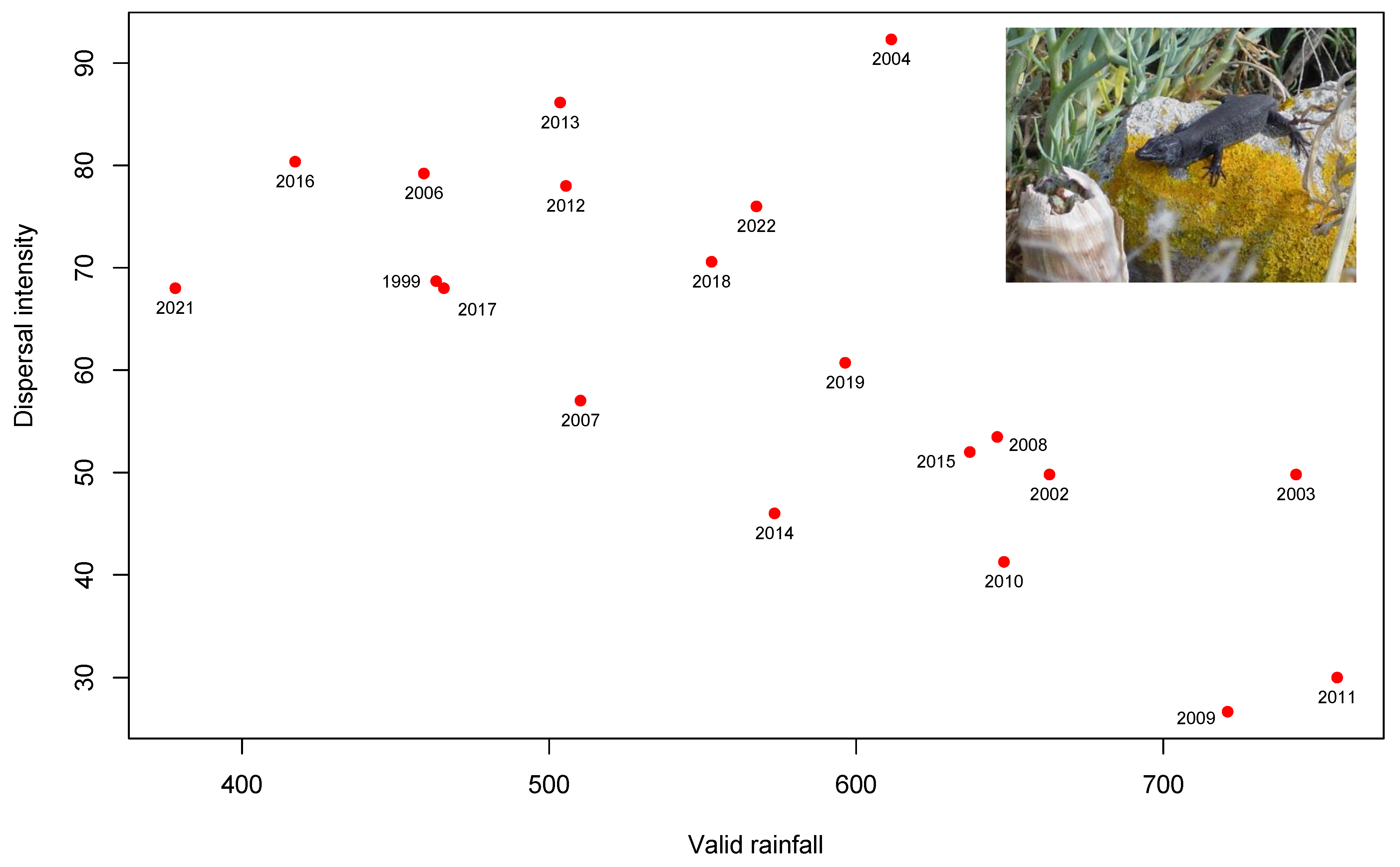
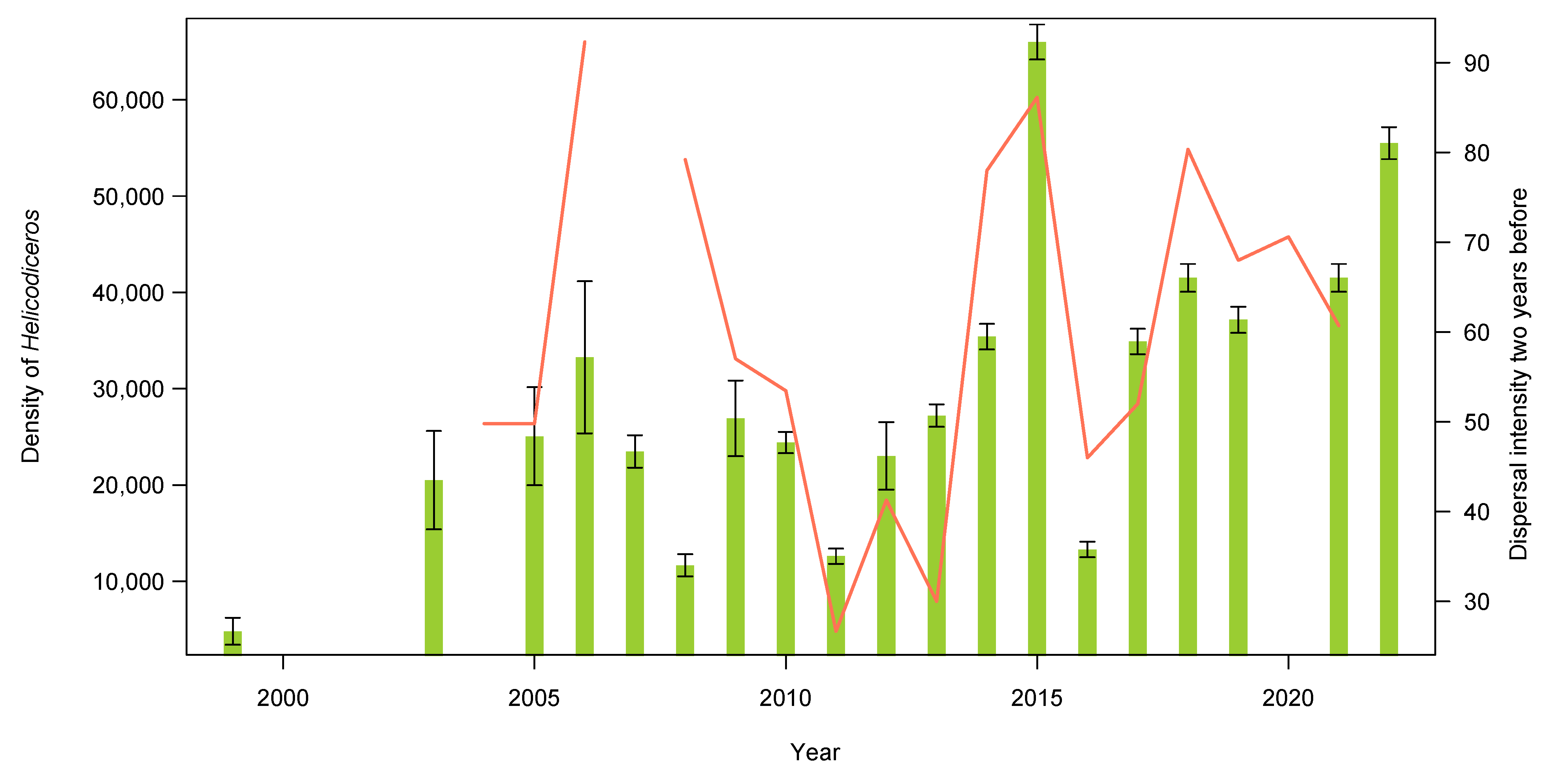
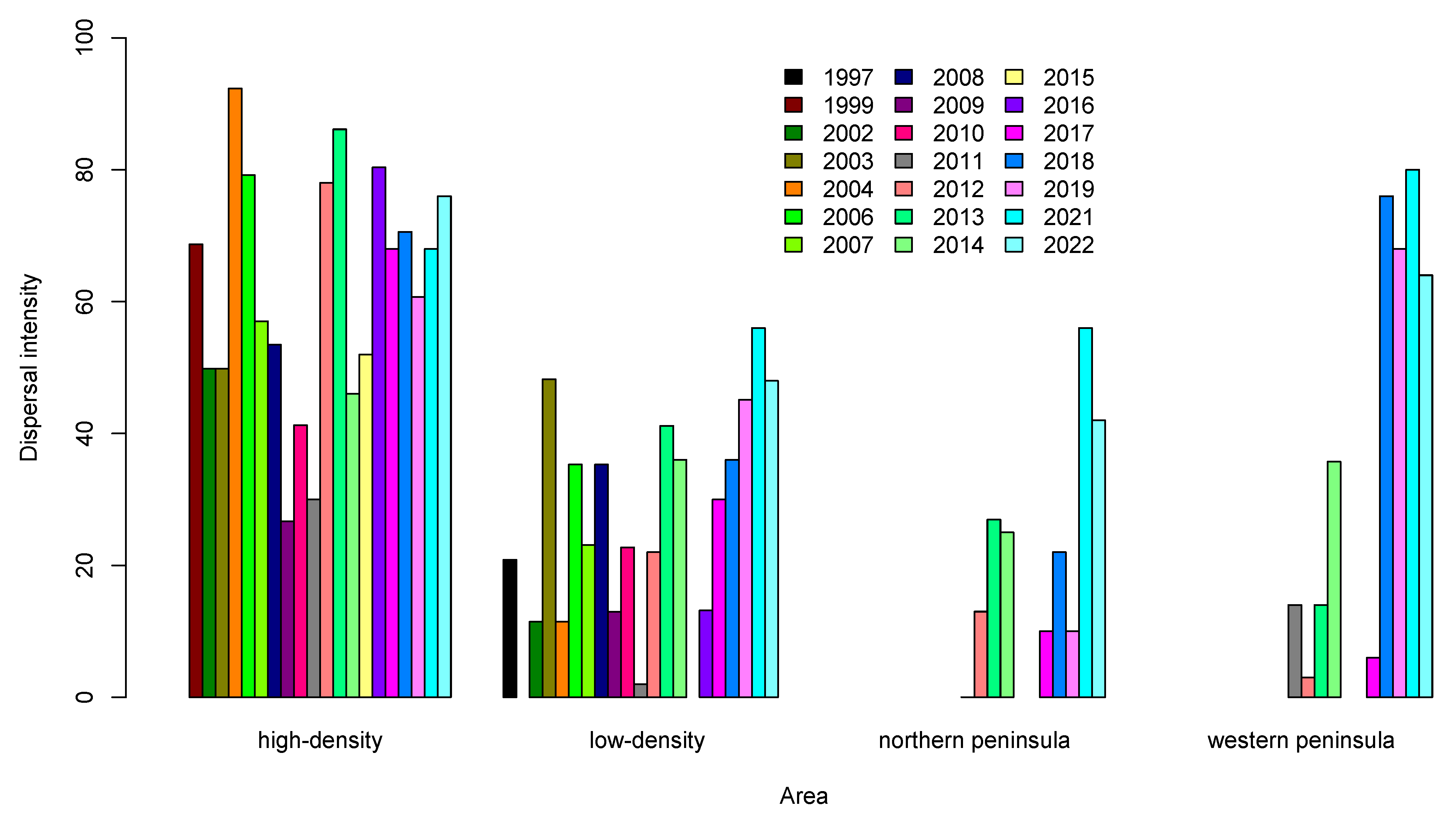
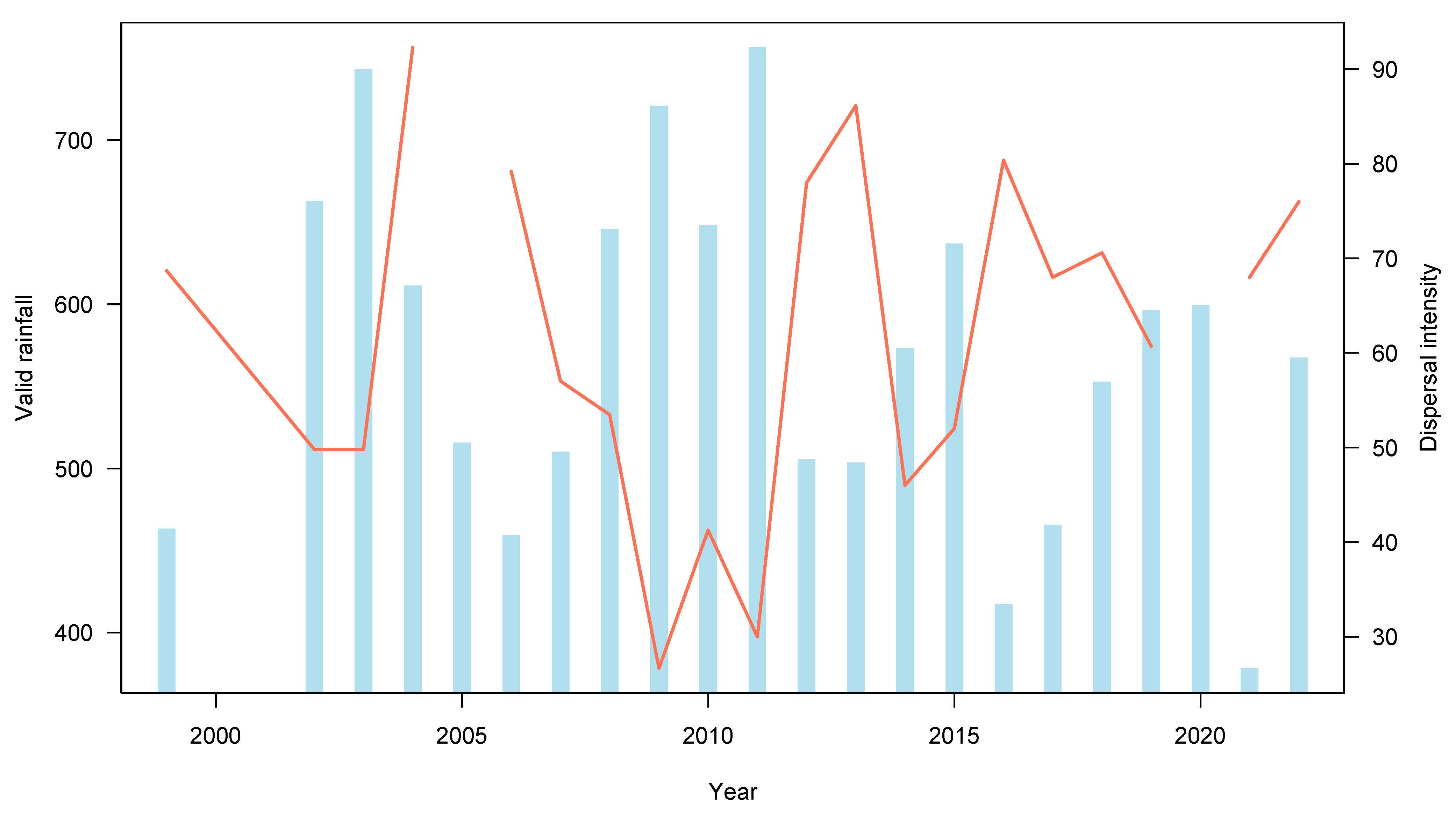
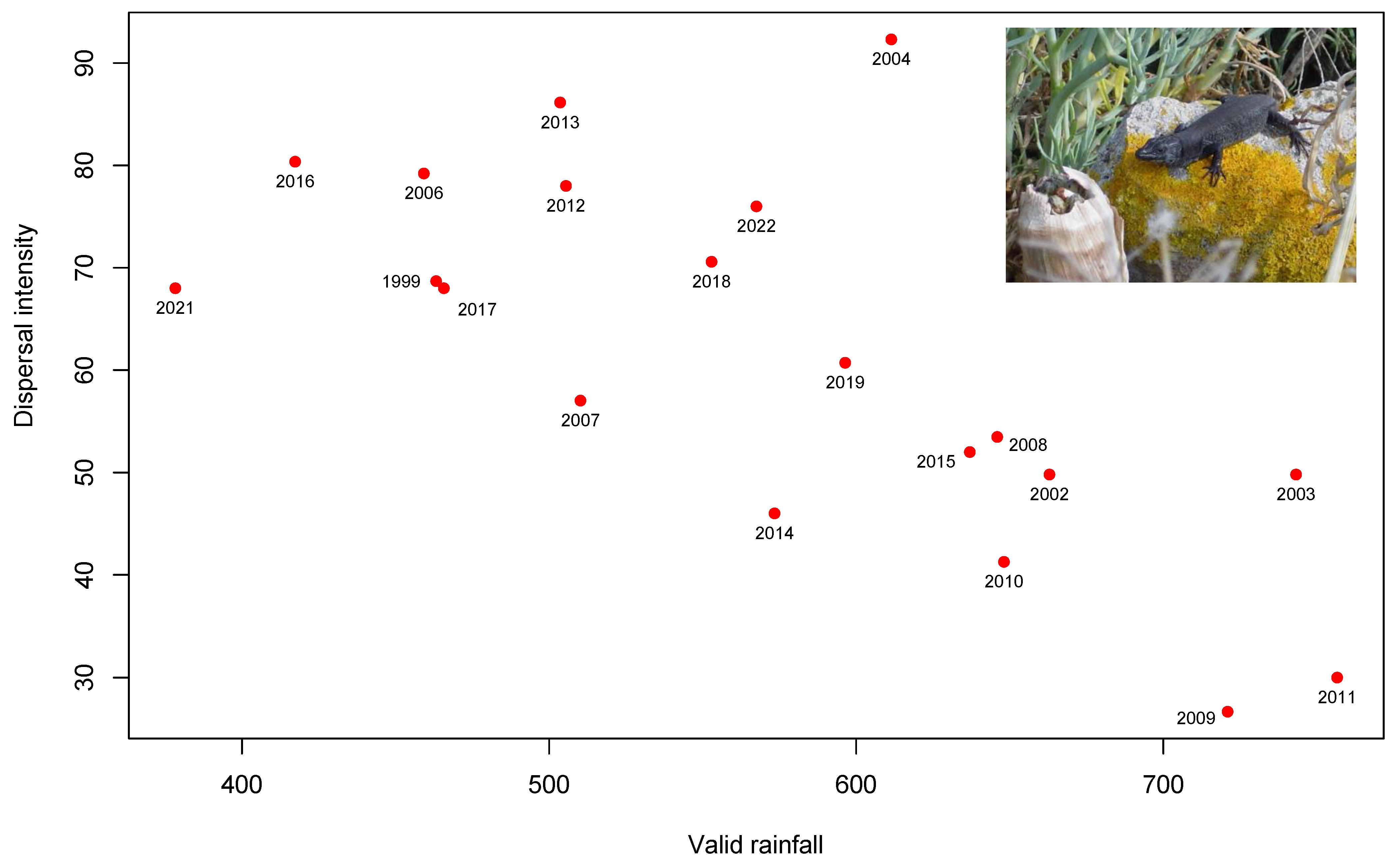
3.2. Seed Dispersal
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Density | ±SE | n |
|---|---|---|---|
| 1999 | 4800 | 1397 | 159 |
| 2003 | 20,489 | 5109 | 807 |
| 2005 | 25,052 | 5095 | 500 |
| 2006 | 33,253 | 7904 | 665 |
| 2007 | 23,472 | 1678 | 468 |
| 2008 | 11,651 | 1158 | 233 |
| 2009 | 26,903 | 3933 | 538 |
| 2010 | 24,400 | 1105 | 488 |
| 2011 | 12,600 | 794 | 252 |
| 2012 | 23,002 | 3505 | 460 |
| 2013 | 27,200 | 1166 | 544 |
| 2014 | 35,400 | 1330 | 708 |
| 2015 | 66,003 | 1817 | 1320 |
| 2016 | 13,300 | 815 | 266 |
| 2017 | 34,900 | 1321 | 698 |
| 2018 | 41,500 | 1440 | 830 |
| 2019 | 37,150 | 1363 | 743 |
| 2021 | 41,500 | 1440 | 830 |
| 2022 | 55,500 | 1666 | 1112 |
| Year | n | Lizard Density | ±SE | n | Plant Density | ±SE | Area |
|---|---|---|---|---|---|---|---|
| 2003 | 119 | 3706 | 632.11 | 0 | 0 | Northern peninsula | |
| 2003 | 214 | 2110 | 413.81 | 214 | 5095.25 | 4385.73 | Western peninsula |
| 2003 | 180 | 3282 | 304 | 1316 | 18,800 | 518 | High-density east |
| 2003 | 114 | 2007 | 233 | 670 | 9571 | 370 | High-density west |
| 2003 | 147 | 2660 | 272 | 179 | 2557 | 191 | Low-density east |
| 2003 | 135 | 2125 | 226 | 68 | 971 | 118 | Low-density west |
| 2009 | 196 | 3410 | 303 | 815 | 5821 | 204 | High-density east |
| 2009 | 123 | 2567 | 300 | 217 | 1833 | 124 | High-density west |
| 2009 | 112 | 2780 | 322 | 46 | 329 | 48.4 | Low-density east |
| 2009 | 128 | 2420 | 264 | 69 | 493 | 59.3 | Low-density west |
| 2009 | 133 | 2258 | 240 | 220 | 1105 | 74.9 | Western peninsula |
| 2009 | 118 | 4258 | 482 | 0 | 0 | Northern peninsula | |
| 2010 | 193 | 3416 | 302 | 1026 | 4580 | 143 | High-density east |
| 2010 | 135 | 2578 | 273 | 122 | 699 | 64.1 | High-density west |
| 2010 | 117 | 1696 | 193 | 29 | 167 | 31.5 | Low-density east |
| 2010 | 171 | 2854 | 268 | 106 | 631 | 61.3 | Low-density west |
| 2010 | 117 | 3911 | 445 | 0 | 0 | Northern peninsula | |
| 2010 | 184 | 3063 | 277 | 230 | 1174 | 77.4 | Western peninsula |
| 2015 | 142 | 2408 | 252 | 3077 | 43,957 | 792 | High-density east |
| 2015 | 202 | 2451 | 226 | 697 | 9957 | 377 | Low-density east |
| 2015 | 87 | 1719 | 232 | 1197 | 21,373 | 618 | High-density west |
| 2015 | 113 | 1529 | 185 | 1102 | 15,742 | 474 | Low-density west |
| 2015 | 103 | 1534 | 195 | 30 | 714 | 130 | Northern peninsula |
| 2015 | 58 | 758 | 123 | 529 | 5971 | 260 | Western peninsula |
| 2018 | 161 | 1295 | 141 | 152 | 893 | 72.9 | Inner Eastern |
| 2018 | 154 | 1446 | 156 | 239 | 1423 | 92 | Coastal Eastern |
| 2018 | 110 | 2661 | 351 | 24 | 429 | 87.5 | Northern peninsula |
| 2018 | 280 | 2089 | 170 | 1059 | 6619 | 203 | Western peninsula |
| 2018 | 230 | 1643 | 108 | High-density east | |||
| 2018 | 146 | 1673 | 195 | High-density west | |||
| 2018 | 190 | 1357 | 98.4 | 148 | 1021 | 85.4 | Low-density east |
| 2018 | 201 | 2188 | 207 | 337 | 2407 | 131 | Low-density west |
| 2019 | 270 | 1598 | 126 | 626 | 3862 | 155 | Western peninsula |
| 2019 | 129 | 1969 | 227 | 120 | 1007 | 91.9 | Northern peninsula |
| 2021 | 86 | 1458 | 202 | 7 | 250 | 94.5 | Northern peninsula |
| 2021 | 154 | 1623 | 166 | 626 | 7716 | 309 | Western peninsula |
| 2021 | 53 | 495 | 90.8 | 49 | 411 | 60.6 | Inner Eastern |
| 2021 | 66 | 670 | 109 | 65 | 381 | 47.6 | Coastal Eastern |
| 2021 | 204 | 2023 | 195 | 1786 | 12,757 | 302 | High-density east |
| 2021 | 138 | 2110 | 239 | 432 | 3724 | 179 | High-density west |
| 2021 | 172 | 1602 | 171 | 208 | 1486 | 103 | Low-density east |
| 2021 | 152 | 1807 | 193 | 254 | 1814 | 114 | Low-density west |
| Year | Validrainfall | Iqindexvalid | Qembergervalid | Faecal Sample | % Dispersal | Area |
|---|---|---|---|---|---|---|
| 1999 | 463.2 | 83.35618994 | 52.01351893 | 98 | 68.6868687 | high-density |
| 2002 | 662.9 | 56.5574049 | 37.39399188 | 492 | 49.796748 | high-density |
| 2003 | 743.2 | 73.74305344 | 54.36904352 | 492 | 49.796748 | high-density |
| 2004 | 611.4 | 81.54212812 | 51.23287728 | 65 | 92.3076923 | high-density |
| 2006 | 459.2 | 63.89507521 | 38.88416469 | 101 | 79.2079208 | high-density |
| 2007 | 510.2 | 52.4549868 | 33.20414184 | 114 | 57.02 | high-density |
| 2008 | 645.9 | 62.29556038 | 43.56115424 | 101 | 53.4653465 | high-density |
| 2009 | 720.9 | 75.60845234 | 47.88530657 | 75 | 26.6666667 | high-density |
| 2010 | 648.1 | 80.70159783 | 50.62556883 | 63 | 41.2698413 | high-density |
| 2011 | 756.6 | 75.1625245 | 50.10932663 | 50 | 30 | high-density |
| 2012 | 505.5 | 87.00107253 | 52.17241379 | 100 | 78 | high-density |
| 2013 | 50.6 | 54.64305975 | 34.67078189 | 101 | 86.1386139 | high-density |
| 2014 | 573.4 | 56.02777172 | 37.33664292 | 50 | 46 | high-density |
| 2015 | 637 | 63.67560676 | 40.4475043 | 50 | 52 | high-density |
| 2016 | 417.3 | 70.4336912 | 45.14352736 | 56 | 80.3571429 | high-density |
| 2017 | 465.7 | 46.58086127 | 29.0967661 | 50 | 68 | high-density |
| 2018 | 552.9 | 46.58086127 | 30.9718387 | 51 | 70.5882353 | high-density |
| 2019 | 596.4 | 62.25900377 | 37.89581905 | 56 | 60.7143 | high-density |
| 2021 | 378.3 | 67.86940073 | 43.75732056 | 50 | 68 | high-density |
| 2022 | 567.5 | 26.2916 | 50 | 76 | high-density | |
| 2011 | 756.6 | 75.1625245 | 50.10932663 | 50 | 0 | northern peninsula |
| 2012 | 505.5 | 87.00107253 | 52.17241379 | 100 | 13 | northern peninsula |
| 2013 | 503.6 | 54.64305975 | 34.67078189 | 52 | 26.923077 | northern peninsula |
| 2014 | 573.4 | 56.02777172 | 37.33664292 | 48 | 25 | northern peninsula |
| 2017 | 465.7 | 46.58086127 | 29.0967661 | 50 | 10 | northern peninsula |
| 2018 | 552.9 | 46.58086127 | 30.9718387 | 50 | 22 | northern peninsula |
| 2019 | 596.4 | 62.25900377 | 37.89581905 | 50 | 10 | northern peninsula |
| 2021 | 378.3 | 67.86940073 | 43.75732056 | 50 | 56 | northern peninsula |
| 2022 | 567.5 | 26.2916 | 50 | 42 | northern peninsula | |
| 2011 | 756.6 | 75.1625245 | 50.10932663 | 50 | 14 | western peninsula |
| 2012 | 505.5 | 87.00107253 | 52.17241379 | 100 | 3 | western peninsula |
| 2013 | 503.6 | 54.64305975 | 34.67078189 | 50 | 14 | western peninsula |
| 2014 | 573.4 | 56.02777172 | 37.33664292 | 42 | 35.7142857 | western peninsula |
| 2017 | 465.7 | 46.58086127 | 29.0967661 | 50 | 6 | western peninsula |
| 2018 | 552.9 | 46.58086127 | 30.9718387 | 25 | 76 | western peninsula |
| 2019 | 596.4 | 62.25900377 | 37.89581905 | 25 | 68 | western peninsula |
| 2021 | 378.3 | 67.86940073 | 43.75732056 | 25 | 80 | western peninsula |
| 2022 | 567.5 | 26.2916 | 25 | 64 | western peninsula | |
| 1997 | 490 | 93.98154827 | 71.57307191 | 48 | 20.8333333 | low-density |
| 2002 | 662.9 | 56.5574049 | 37.39399188 | 262 | 11.4503817 | low-density |
| 2003 | 743.2 | 73.74305344 | 54.36904352 | 56 | 48.2142857 | low-density |
| 2004 | 611.4 | 81.54212812 | 51.23287728 | 262 | 11.4503817 | low-density |
| 2006 | 459.2 | 63.89507521 | 38.88416469 | 102 | 35.2941177 | low-density |
| 2007 | 510.2 | 52.4549868 | 33.20414184 | 91 | 23.08 | low-density |
| 2008 | 645.9 | 62.29556038 | 43.56115424 | 51 | 35.2941177 | low-density |
| 2009 | 720.9 | 75.60845234 | 47.88530657 | 54 | 12.962963 | low-density |
| 2010 | 648.1 | 80.70159783 | 50.62556883 | 88 | 22.7272727 | low-density |
| 2011 | 756.6 | 75.1625245 | 50.10932663 | 50 | 2 | low-density |
| 2012 | 505.5 | 87.00107253 | 52.17241379 | 100 | 22 | low-density |
| 2013 | 503.6 | 54.64305975 | 34.67078189 | 102 | 41.1764706 | low-density |
| 2014 | 573.4 | 56.02777172 | 37.33664292 | 50 | 36 | low-density |
| 2016 | 417.3 | 70.4336912 | 45.14352736 | 38 | 13.1578947 | low-density |
| 2017 | 465.7 | 46.58086127 | 29.0967661 | 50 | 30 | low-density |
| 2018 | 552.9 | 46.58086127 | 30.9718387 | 50 | 36 | low-density |
| 2019 | 596.4 | 62.25900377 | 37.89581905 | 51 | 45.098 | low-density |
| 2021 | 378.3 | 67.86940073 | 43.75732056 | 50 | 56 | low-density |
| 2022 | 567.5 | 26.2916 | 50 | 48 | low-density |
References
- Traveset, A. El paper dels mutualismes planta-animal als ecosistemes insulars. In Ecologia de Les Illes; Alcover, J.A., del Hoyo, J., Eds.; Monografies de la Societat d’Historia Natural de les Balears: Palma de Mallorca, Spain, 1999; Volume 6, pp. 9–33. [Google Scholar]
- Fricke, E.C.; Ordóñez, A.; Rogers, H.S.; Svenning, J.C. Effects of defaunation on plant’s capacity to track climatic change. Science 2022, 375, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Castilla, A.M. Podarcis lilfordi from the Balearic Islands as a potential disperser of the rare Mediterranean plant Withania frutescens. Acta Oecol. 1999, 20, 103–107. [Google Scholar] [CrossRef]
- Pérez-Mellado, V.; Traveset, A. Relationships between plants and Mediterranean lizards. Nat. Croat. 1999, 8, 275–285. [Google Scholar]
- Olesen, J.M.; Valido, A. Lizards and birds as generalized pollinators and seed dispersers of island plants. In Island Ecology; Fernández-Palacios, J.M., Morici, M., Eds.; AEET/Cabildo Insular de La Palma: Canary Islands, Spain, 2004; pp. 229–249. [Google Scholar]
- Valido, A.; Olesen, J.M. The Importance of Lizards as Frugivores and Seed Dispersers. In Seed Dispersal. Theory and Its Application in a Changing World; Dennis, A.J., Schupp, E.W., Green, R.J., Wescott, D.A., Eds.; CAB International: Wallingford, UK, 2007; pp. 124–147. [Google Scholar]
- Valido, A.; Olesen, J.M. Frugivory and seed dispersal by lizards: A global review. Front. Ecol. Evol. 2019, 7, 1–97. [Google Scholar] [CrossRef] [Green Version]
- Correcher, E.J.; Hervías-Parejo, S.; Ruíz, R.; Sauroy-Toucouère, S.; Traveset, A. Environmental and morphological drivers of mutualistic plant-lizard interactions: A global review. Ecography 2023, 2023, e06425. [Google Scholar] [CrossRef]
- Herrera, C.M. Mediterranean plant-bird seed dispersal systems: The roles of history and adaptation. In Proceedings of the 6th International Conference on Mediterranean Climate Ecosystems, Plant-Animal Interactions in Mediterranean-Type Ecosystems, Maleme (Crete), Greece, 23–27 September 1991; Thanos, C.A., Ed.; University of Athens: Athens, Greece, 1991; pp. 241–250. [Google Scholar]
- Herrera, C.M. Seed dispersal by vertebrates. In Plant-Animal Interactions. An Evolutionary Approach; Herrera, C.M., Pellmyr, O., Eds.; Wiley-Blackwell Publishing: Maiden, NC, USA, 2002; pp. 185–208. [Google Scholar]
- Roselló, J.A.; Sáez, L. Notes on some Balearic Araceae. Acta Bot. Barcinon. 1997, 44, 169–174. [Google Scholar]
- Fridlender, A. Répartition, écologie et conservation de l’Arum mange-mouches (Helicodiceros muscivorus (L.fil.) Engler (Araceae). Webbia 2000, 55, 7–35. [Google Scholar] [CrossRef]
- Mansion, G.; Rosenbaum, G.; Schonenberger, N.; Bachetta, G.; Rosselló, J.A.; Conti, E. Phylogenetic Analysis Informed by Geological History Supports Multiple, Sequential Invasions of the Mediterranean Basin by the Angiosperm Family Araceae. Syst. Biol. 2008, 57, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Nauheimer, L.; Metzler, D.; Renner, S.S. Global history of the ancient monocot family Araceae inferred with models accounting for past continental positions and previous ranges based on fossils. New Phytol. 2012, 195, 938–950. [Google Scholar] [CrossRef]
- Boyce, P.C. The genera Dracunculus and Helicodiceros (Araceae: Aroideae). Thaiszia 1994, 4, 175–182. [Google Scholar]
- Contandriopoulos, J.M.; Cardona, M.A. Original character of the endemic flora in the Balearic Islands. Bot. Helv. 1984, 94, 101–132. [Google Scholar]
- Pérez-Cembranos, A.; León, A.; Pérez-Mellado, V. Omnivory of an insular lizard: Sources of variation in the diet of Podarcis lilfordi (Squamata, Lacertidae). PLoS ONE 2016, 11, e0148947. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Mellado, V.; Riera, N.; Hernández-Estévez, J.A.; Piccolo, V.; Potter, C. A complex case of interaction between lizards and plants. The dead horse arum (Dracunculus muscivorus) and the Balearic lizard (Podarcis lilfordi). In Mainland and Insular Lacertid Lizards: A Mediterranean Perspective; Corti, C., Lo Cascio, P., Biaggini, M., Eds.; Firenze University Press: Firenze, Italy, 2007; pp. 133–160. [Google Scholar]
- Pérez-Mellado, V.; Cortázar, G.; López-Vicente, M.; Perera, A.; Sillero, N. Interactions between the Balearic lizard, Podarcis lilfordi and the plant Dracunculus muscivorus. Amphibia-Reptilia 2000, 21, 223–226. [Google Scholar] [CrossRef]
- Pérez-Mellado, V.; Riera, N.; Piccolo, V.; Potter, C. Mutualismo en ecosistemas insulares: La interacción de dos endemismos mediterráneos, la Rapa mosquera, Dracunculus muscivorus y la lagartija balear, Podarcis lilfordi. Rev. Menorca 2006, 87, 45–77. [Google Scholar]
- Jansà i Clar, A. Climatologia de Menorca. In Enciclopèdia de Menorca I. Geografia Física; Vidal-Hernández, J.M., Ed.; Obra Cultural Balear de Menorca: Maó, Spain, 1979; pp. 89–160. [Google Scholar]
- Pérez-Mellado, V. Podarcis lilfordi (Günther, 1874). In Reptiles; Salvador, A., Ramos, M.A., Eds.; In Fauna Ibérica, Volume 10. Ramos, M.A. et al., Eds.; Museo Nacional de Ciencias Naturales, CSIC: Madrid, Spain, 1998; pp. 272–282. [Google Scholar]
- Pérez-Mellado, V.; Hernández-Estévez, J.A.; García-Díez, T.; Terrassa, B.; Ramón, M.M.; Castro, J.A.; Picornell, A.; Martín-Vallejo, F.J.; Brown, R.P. Population density in Podarcis lilfordi (Squamata, Lacertidae), a lizard species endemic to small islets in the Balearic Islands (Spain). Amphibia-Reptilia 2008, 29, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Angioy, A.M.; Stensmyr, M.C.; Urru, I.; Puliafito, M.; Collu, I.; Hansson, B.S. Function of the heater: The dead horse arum revisited. Proc. R. Soc. B-Biol. Sci. 2004, 271 (Suppl. S3), S13–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castroviejo, S.; Galán, A. Helicodiceros Schott ex Koch [nom. Cons.]. In Flora Ibérica; Castroviejo, S., Galán, A., Eds.; Real Jardín Botánico (C.S.I.C.): Madrid, Spain, 2008; Volume XVIII, pp. 293–296. [Google Scholar]
- Traveset, A. La lagartija balear, una eficaz polinizadora y dispersante de plantas. Quercus 1997, 139, 20–22. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to distance sampling. Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; Vienna, Austria. 2022. Available online: https://www.R-project.org/ (accessed on 21 March 2022).
- Ficetola, G.F.; Barzaghi, B.; Melotto, A.; Muraro, M.; Lunghi, E.; Canedoli, C.; Lo Parrino, E.; Nanni, V.; Silva-Rocha, I.; Urso, A.; et al. N-mixture models reliably estimate the abundance of small vertebrates. Sci. Rep. 2018, 8, 10357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Royle, J.A.; Dawson, D.K.; Bates, S. Modeling abundance effects in distance sampling. Ecology 2004, 85, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- AEMET Open Data, Agencia Estatal de Meteorología. Available online: https://opendata.aemet.es/centrodedescargas/inicio (accessed on 20 September 2022).
- Sahin, S. An aridity index defined by precipitation and specific humidity. J. Hydrol. 2012, 444–445, 199–208. [Google Scholar] [CrossRef]
- Emberger, L. Sur une formule climatique et ses applications en botanique. La Météorologie 1932, nov.dic, 423–432. [Google Scholar]
- Crawley, M.J. The R Book, 2nd ed.; Wiley: Chichester, UK, 2013. [Google Scholar]
- Koenker, R.W. Quantile Regression; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Koenker, R.W.; Chernozhukov, V.; Xuming, H.; Limin, P. Handbook of Quantile Regression; Chapman and Hall/CR: Boca Raton, FL, USA, 2017. [Google Scholar]
- García-Febrero, O.; free-lance biologist, Maó, Menorca, Balearic Islands, Spain; Flora Vascular de l’Illa de l’Aire; A Catalogue of the flowering plants of Aire Island, Menorca. Unpublished work. 1998.
- Rodríguez Femenías, J.J. Catálogo Razonado de Las Plantas Vasculares de Menorca; Imprenta Fàbregues: Maó, Spain, 1868. [Google Scholar]
- Rodríguez Femenías, J.J. Flórula de Menorca; Imprenta Fàbregues: Maó, Spain, 1904. [Google Scholar]
- Pérez-Mellado, V. Estudio ecológico de la Lagartija Balear Podarcis lilfordi (GÜNTHER, 1874) en Menorca. Rev. Menorca 1989, 80, 455–511. [Google Scholar]
- Pérez-Mellado, V. Les Sargantanes de Les Balears; Quaderns de Natura de les Balears; Edicions Documenta Balear: Palma de Mallorca, Spain, 2009. [Google Scholar]
- Gillespie, R.G. Ecological release. In Encyclopedia of Islands; Gillespie, R.G., Clague, D.A., Eds.; University of California Press: Berkeley, CA, USA, 2009; pp. 251–253. [Google Scholar]
- Kjellsson, G. Seed fate in a population of Carex pilulifera L. 1. Seed dispersal and ant-seed mutualism. Oecologia 1985, 67, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Lidén, M. Fumariaceae. In The Families and Genera of Vascular Plants. V. 2: Dicotyledons. Magnoliid, Hamamelid and Caryophyllid Families; Kubitzki, K., Ed.; Springer: Berlin, Germany, 1993; pp. 310–318. [Google Scholar]
- Van der Pijl, L. Principles of Dispersal in Higher Plants; Springer: Berlin, Germany, 1972. [Google Scholar]
- Tinaut, A.; Universidad de Granada, Granada, Spain. Personal communication, 2015.
- Vogel, P. On an ant-lizard interaction. Oecologia 1983, 58, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Mellado, V.; Corti, C. Dietary adaptations and herbivory in lacertid lizards of the genus Podarcis from western Mediterranean islands (Reptilia: Sauria). Bonn. Zool. Beitr. 1993, 4, 193–220. [Google Scholar]
- Aronne, C.; Wilcock, C.C. First evidence of myrmecochory in fleshyfruited shrubs of the Mediterranean region. New Phytol. 1994, 127, 781–788. [Google Scholar] [CrossRef]
- Herrera, C.M. Plant-vertebrate seed dispersal systems in the Mediterranean: Ecological, Evolutionary, and Historical Determinants. Annu. Rev. Ecol. Syst. 1995, 26, 705–727. [Google Scholar] [CrossRef]
- Lord, J.M.; Markey, A.S.; Marshall, J. 4. Have Frugivores Influenced the Evolution of Fruit Traits in New Zealand? In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CABI Publishing: Wallingford, UK, 2002; pp. 55–68. [Google Scholar]
- Valido, A.; Nogales, M. Frugivory and seed dispersal by the lizard Gallotia galloti (Lacertidae) in a xeric habitat of the Canary Islands. Oikos 1994, 70, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Traveset, A. Seed dispersal of Cneorum tricoccon L. (Cneoraceae) by lizards and mammals in the Balearic Islands. Acta Oecol. 1995, 16, 171–178. [Google Scholar]
- Adler, P.B.; Levine, J.M. Contrasting relationships between precipitation and species richness in space and time. Oikos 2007, 116, 221–232. [Google Scholar] [CrossRef]
- Mayo, S.L.; Bogner, J.; Boyce, P.C. The Genera of Araceae; Royal Botanic Gardens: Kew, UK, 1997. [Google Scholar]
- Maurer, G.E.; Hallmark, A.J.; Brown, R.F.; Sala, O.E.; Collins, S.L. Sensitivity of primary production to precipitation across the United States. Ecol. Lett. 2020, 23, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Le Houéron, H.N.; Hoste, C.H. Rangeland production and annual rainfall relations in the Mediterranen Basin and the African Sahelian and Sudanian Zones. J. Range Manag. 1977, 30, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Le Houéron, H.N.; Bingham, R.L.; Skerjek, W. Relationship between the variability of primary production and the variability of annual precipitation in world arid lands. J. Arid Environ. 1988, 15, 1–18. [Google Scholar] [CrossRef]
- Griffiths, A.D.; Christian, K.A. Diet and habitat use of frillneck lizards in a seasonal tropical environment. Oecologia 1996, 106, 39–48. [Google Scholar] [CrossRef]
- Christian, K.A.; Webb, J.K.; Schultz, T.J. Energetics of blue-tongue lizards (Tiliqua scincoides) in a seasonal tropical environment. Oecologia 2003, 136, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Barnett, K.L.; Facey, S.L. Grasslands, Invertebrates, and Precipitation: A Review of the Effects of Climatic Change. Front. Plant Sci. 2016, 7, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Clarke, I.P.; Taylor, M.E.; Whitehouse, A.T. Changing precipitation patterns alter plant community dynamics and succession in an ex-arable grassland. Funct. Ecol. 2004, 18, 648–655. [Google Scholar] [CrossRef]
- Lister, B.C.; García, A. Climate-driven declines in arthropod abundance restructure a rainforest food web. Proc. Natl. Acad. Sci. USA 2018, 115, 10397–10406. [Google Scholar] [CrossRef] [Green Version]
- Buckland, S.M.; Grime, J.P.; Hodgson, J.G.; Thompson, K. A comparison of plant responses to the extreme drought of 1995 in northern England. J. Ecol. 1997, 85, 875–882. [Google Scholar] [CrossRef]
- Morecroft, M.D.; Bealey, C.E.; Howells, E.; Rennie, S.; Woiwod, I.P. Effects of drought on contrasting insect and plant species in the UK in the mid-1990s. Global Ecol. Biogeogr. 2002, 11, 7–22. [Google Scholar] [CrossRef]
- Staley, J.T.; Hodgson, C.J.; Mortimer, S.R.; Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Taylor, M.E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. Eur. J. Soil Biol. 2007, 43, 189–198. [Google Scholar] [CrossRef]
- Anderson, D.C. Below-ground herbivory in natural communities: A review emphasising fossorial animals. Q. Rev. Biol. 1987, 62, 261–286. [Google Scholar] [CrossRef]
- Brown, V.K.; Gange, A.C. Insect herbivory below ground. Adv. Ecol. Res. 1990, 20, 1–58. [Google Scholar] [CrossRef]
- Branquart, R.D.; Kime, M.; Dufrene, J.; Tavernier, M.; Wauthy, G. Macroarthropod-habitat relationships in oak forests in South Belgium. 1. Environments and communities. Pedobiologia 1995, 39, 243–263. [Google Scholar]
- McNaugton, S.J.; Oesterheld, M.; Frank, D.A.; Williams, K.J. Primary and Secondary Production in Terrestrial Ecosystems. In Comparative Analyses of Ecosystems; Cole, J., Lovett, G., Findlay, S., Eds.; Springer: New York, NY, USA, 1991; pp. 120–139. [Google Scholar]
- Cooper, W.E.; Vitt, L.J. Distribution, extent and evolution of plant consumption by lizards. J. Zool. 2002, 257, 487–517. [Google Scholar] [CrossRef] [Green Version]
- Herrel, A.; Vanhooydonck, B.; Van Damme, R. Omnivory in lacertid lizards: Adaptive evolution or constraint? J. Evol. Biol. 2004, 17, 974–984. [Google Scholar] [CrossRef]
- Van Damme, R. Evolution of herbivory in lacertid lizards: Effects of insularity and body size. J. Herpetol. 1999, 33, 663–674. [Google Scholar] [CrossRef]
- Debussche, M.; Cortez, J.; Rimbault, I. Variation in fleshy fruit composition in the Mediterranean region: The importance of ripening season, life-form, fruit type and geographical distribution. Oikos 1987, 49, 244–252. [Google Scholar] [CrossRef]
- Herrera, C.M. A study of avian frugivores, bird-dispersed plants, and their interaction in Mediterranean scrublands. Ecol. Monogr. 1984, 54, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M. Capítulo 5. Dispersión de semillas por animales en el Mediterráneo. Ecología y evolución. In Ecosistemas Mediterráneos. Análisis Funcional; Zamora, R., Pugnaire, F.I., Eds.; Servicio de Publicaciones del C.S.I.C.: Madrid, Spain, 2001; pp. 132–152. [Google Scholar]
- Herrera, C.M. Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits: A 12-years study. Ecol. Monogr. 1998, 68, 511–538. [Google Scholar] [CrossRef]
- Suc, J.P. Origin and evolution of the Mediterranean vegetation and climate in Europe. Nature 1984, 307, 429–432. [Google Scholar] [CrossRef]
- Herrera, C.M. Determinants of plant-animal coevolution: The case of mutualistic dispersal of seeds by vertebrates. Oikos 1985, 44, 132–141. [Google Scholar] [CrossRef]
- Herrera, C.M. Interspecific variation in fruit shape: Allometry, phylogeny, and adaptation to dispersal agents. Ecology 1992, 73, 1832–1841. [Google Scholar] [CrossRef]
- Herrera, C.M. Historical effects and sorting processes as explanations for contemporary ecological patterns: Character syndromes in Mediterranean woody plants. Am. Nat. 1992, 140, 421–446. [Google Scholar] [CrossRef] [Green Version]
- Jordano, P. Patterns of mutualistic interactions on pollination and seed dispersal: Connectance, dependence asymmetries, and coevolution. Am. Nat. 1987, 129, 657–677. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Cembranos, A.; Pérez-Mellado, V. Long-Term Seed Dispersal within an Asymmetric Lizard-Plant Interaction. Animals 2023, 13, 973. https://doi.org/10.3390/ani13060973
Pérez-Cembranos A, Pérez-Mellado V. Long-Term Seed Dispersal within an Asymmetric Lizard-Plant Interaction. Animals. 2023; 13(6):973. https://doi.org/10.3390/ani13060973
Chicago/Turabian StylePérez-Cembranos, Ana, and Valentín Pérez-Mellado. 2023. "Long-Term Seed Dispersal within an Asymmetric Lizard-Plant Interaction" Animals 13, no. 6: 973. https://doi.org/10.3390/ani13060973
APA StylePérez-Cembranos, A., & Pérez-Mellado, V. (2023). Long-Term Seed Dispersal within an Asymmetric Lizard-Plant Interaction. Animals, 13(6), 973. https://doi.org/10.3390/ani13060973