1. Introduction
The productivity of extensive beef production systems relies on the sum of several traits. Of these, fertility is one of the most crucial, since it has a major effect on the number of calves born and, therefore, the quantity of weaned calves produced per year [
1]. This has been demonstrated by several studies, which went so far as to quantify reproductive traits as three times more important than production traits in semi-intensive cattle production systems [
2,
3,
4,
5]. Despite the existence of a sizeable environmental effect, it was also demonstrated that there is a considerable genetic component affecting fertility in cows, thus enabling us to select more fertile individuals in order to increase the reproductive efficiency of the whole system [
6]. In this context, there is a general consensus that the ideal beef cow, in terms of fertility, should be precocious (i.e., first calving at 24 months in Retinta) and produce one calf per year [
7,
8]. Therefore, the possibility of selecting individuals that have a genetic predisposition to meet such objectives could represent a key advantage from a breeding point of view.
An increase in the calving interval is one of the most common causes of culling in beef cattle [
9]. However, this reproductive trait, closely associated with overall fertility, is characterized by low heritability and slow genetic progress in cattle [
10]. For this reason, it is rarely included as a selection criterion in breeding programs of beef breeds raised under extensive environmental conditions. Instead, breeding programs focus more on increasing the live weight of the individuals, using growth traits such as weight and average daily weight gain, which correlate negatively with fertility [
11]. However, the inclusion of reproductive traits, either individually or in combination through indexes, has been considered in certain breeds, although it is not yet a very widespread practice [
12]. Cammack, et al. [
10] summarized average values and heritabilities in thirteen different reproduction traits in beef cattle raised under extensive systems reported over 25 years, showing that most of them focused on the effects of the bull and precocity. However, few studies have looked into the individual fertility of cows, most likely due to the difficulty in obtaining a large and reliable dataset in extensive or grazing production systems.
During the last few years, the use of an indirect fertility criterion, including precocity and the calving interval, which can be estimated indirectly based on reproductive records, has been demonstrated as an interesting option for selecting more fertile females in extensively-bred livestock species [
13,
14]. This parameter, called reproductive efficiency (Re), is estimated as the percentage deviation of the number of calvings that a cow has at each age, from the number of calvings that this cow could have had in optimal conditions. In the Retinta breed, the optimum age at first calving was considered two years and one year for the optimal calving interval [
15]. It has demonstrated increased heritability and reliability in comparison with other reproductive traits in beef cattle, such as calving interval or age at first calving [
16]. However, to our knowledge, it has only been analyzed in a few studies in cattle [
17,
18]. In addition, reproductive traits have mostly been evaluated from a genetic point of view, using repeatability models. This methodology assumes that the Re values at the different calvings are manifestations of the same trait, and therefore the (co)variance across the trajectory of the parities of the cow is constant. Despite the fact that this methodology is widely used, Wilson and Réale [
19] concluded that this assumption is not correct since the genetic parameters for this kind of effect vary at different ages. However, there is still a shortage of reports evaluating the changes in such parameters over the lifetime of the cows.
In recent years, the genomic revolution has allowed us to identify single nucleotide polymorphisms (SNPs) associated with phenotypic traits, leading to a better understanding of complex traits. One of the most powerful tools for analyzing such associations is the genome-wide association study (GWAS). In beef, a large number of GWAS analyses have been reported for different traits such as longitudinal, carcass, fatty acid profiles, meat tenderness and quality, and growth traits [
20,
21,
22,
23,
24,
25,
26]. However, GWAS studies on reproductive traits in beef cows are still scarce [
27,
28,
29,
30].
Retinta is an autochthonous breed with unique adaptive characteristics linked to an extensive regime in the Dehesa ecosystem. Nowadays, it is widely bred in the south of Spain, due to its excellent adaptation to the harsh environment characterized by marginal pasturelands and the hot, dry climate [
31]. In 2018, the breed reported more than 42,000 breeding cows, of which 16,850 were enrolled in the Genealogical Book [
32]. Of these, 25% were raised as purebreds, with the rest used in crossbreeding as a maternal line with other continental breeds, such as Charolais and Limousin, with the aim of increasing profitability by using crossbred individuals [
33]. For over 40 years now, the Retinta breeders’ association has not only gathered pedigree records but also a large phenotypic and environmental dataset, including birth and culling records from all the individuals enrolled in the breed [
15]. For this reason, the Retinta breed is an interesting model by which to evaluate quantitative traits accurately, including those related to fertility.
In this study, we aimed to determine the genetic influence on the fertility of Retinta breeding cows by measuring the variance component patterns of reproductive efficiency across the calvings in the Retinta beef breed using random regression models (RRM). Additionally, we performed a preliminary genome-wide association study for reproductive efficiency traits to identify genetic variants, genomic regions, and candidate genes associated with fertility.
3. Results and Discussion
Fertility is one of the key factors in any extensive livestock production system. However, at the same time, it is one of the most neglected from a genetic point of view, since it is difficult to study models in such production conditions. For this reason, the use of indirect traits obtained from pedigree records is becoming an interesting alternative to evaluating fertility at the population level since it allows us to obtain large phenotypic datasets, which are crucial for making reliable estimations [
13,
47]. Using such an approach, we were able to determine the genetic and environmental effects in a large cohort of beef cows bred in extensive conditions. To our knowledge, no previous studies have employed this methodology in cattle.
Re is a novel trait that estimates the deviation between the optimal and real parity number of females at each age of the cows (considering precocity and calving interval) throughout their entire productive lives. Our results showed a wide variability in average Re (across calving) among Retinta cows (
Table 1).
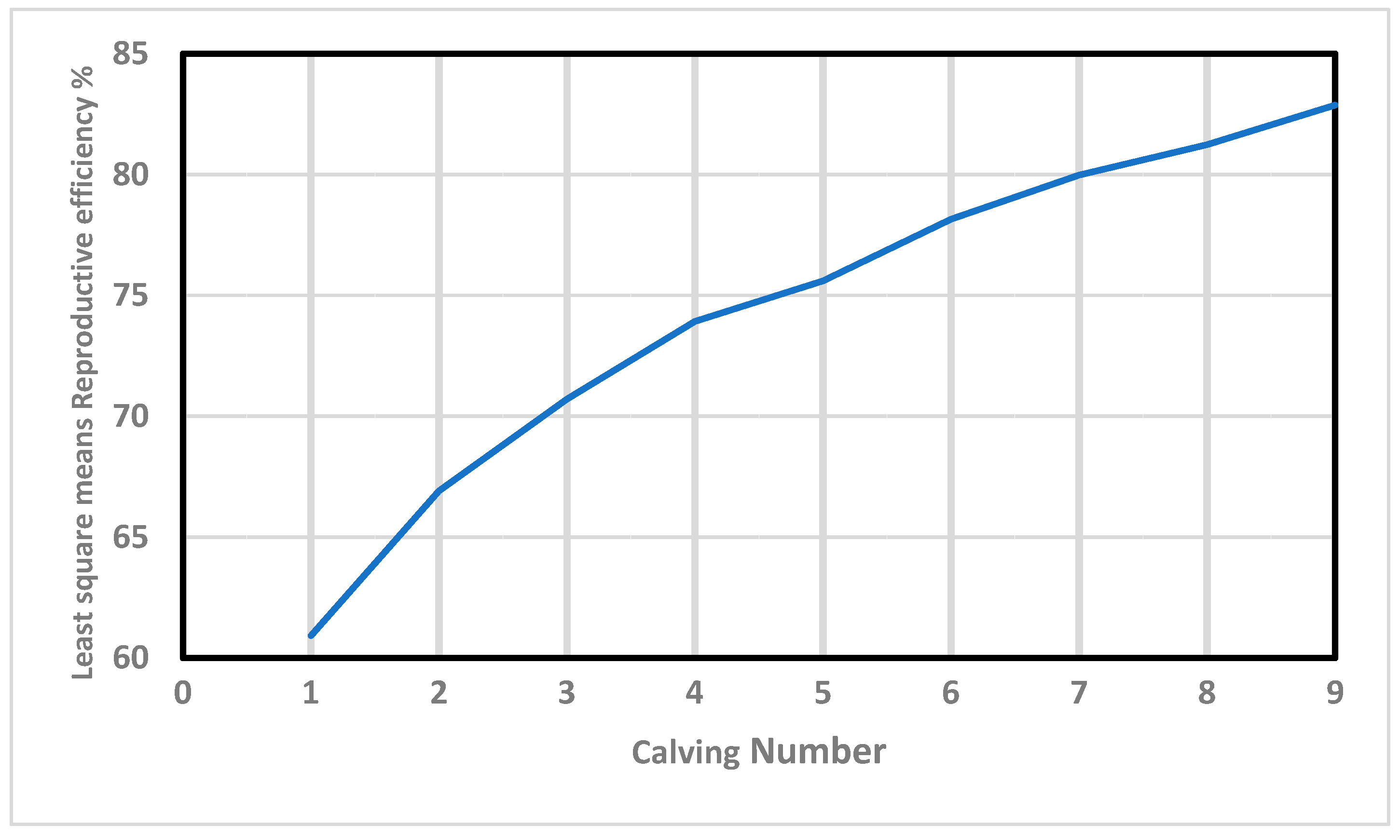
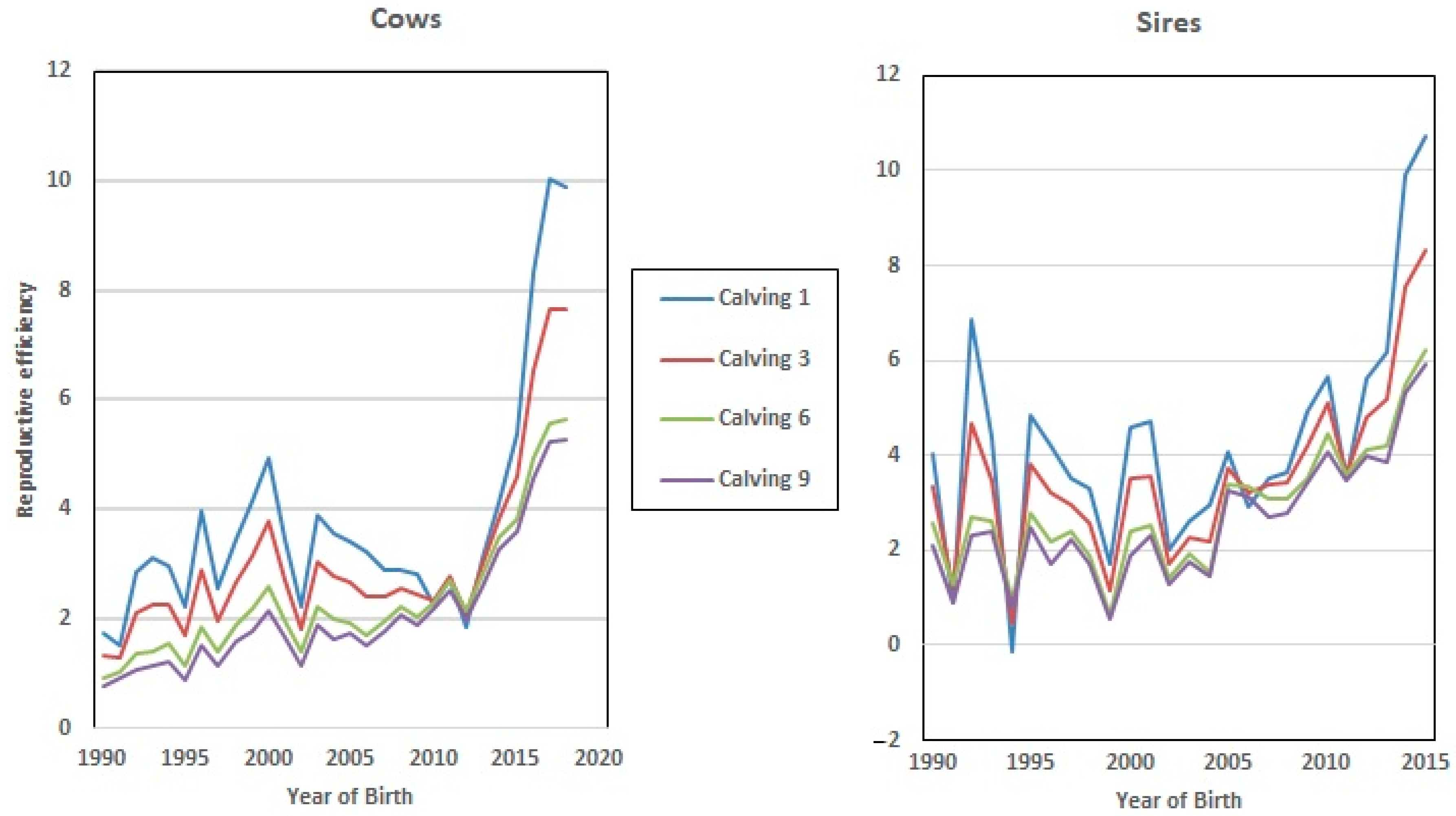
Furthermore, Re increased in value almost linearly with the calving number (
Figure 1). This positive correlation (0.97) agrees with the results obtained by Mercadante, et al. [
48], and Swanepoel and Hoogenboezem [
49], who concluded that the cows that remain longer in the herd present better reproductive behavior during their productive lives. In this context, Balieiro, et al. [
50] demonstrated that the most fertile cows that do not “miss” any breeding seasons are the ones most likely to be allowed to remain in the herd in extensively-raised beef breeds. Interestingly, those “regular and reliable” individuals show high values of Re, demonstrating the validity of this trait as an indirect estimator of fertility.
The same analysis revealed lower variability in the age of first calving (33.9 ± 6.18 months, on average), which can be associated with differences in precocity, as has been convincingly proven in beef breeds [
51,
52]. This fact is particularly important since precocity is negatively related to longevity, and cows starting their reproductive lives at later stages are less likely to be culled for reproductive reasons [
53]. Since precocity is not commonly included in models of calving intervals in beef cattle, we hypothesize that Re may be an interesting estimator of fertility in cows bred in extensive conditions, reducing a potential cause of bias within the evaluations.
3.1. Modeling the Genetic Influence on Re Using a Repeatability Model
In this study, we employed two different approaches to estimate the genetic component affecting Re: the classical Rep model and the RRM. The former is the most common methodology employed, having been used for over 20 years [
16]. However, the latter has been increasingly employed during recent years since it allows us to estimate changes in the variance components of Re (or any reproductive trait) across calvings and, therefore, to select individuals with increased fertility values at any particular age [
54]. However, regardless of which model is employed, fertility traits in livestock bred in extensive conditions must be analyzed using large reproductive datasets, such as the dataset used with Re as the trait of interest.
The variance components obtained using the Rep model are shown in
Table 2. Heritability was particularly high (near 0.3) for a reproductive character in comparison with previous reports on beef cows, which are closer to 0.1 (reviewed by Cammack, et al. [
10]). However, none of these employed Re as a fertility estimator for the cows. Likewise, similar h
2 values (near 0.25) were recently reported in goats and horses [
13,
47]. In addition, it was also noticeable that the permanent environmental effect of the cow accounted for nearly 35% of the total variance and the contemporary group (HYS) for only 9%. This result for the permanent environmental effect is expected in any extensive system in which environmental factors (such as metritis or retained placentas) with poor treatment can permanently delay the cow’s fertility within the season [
55]. Regarding HYS, the lower value fits in with the homogeneity of the production conditions across herds and years in the Spanish Dehesa in which the Retinta cows are bred [
33].
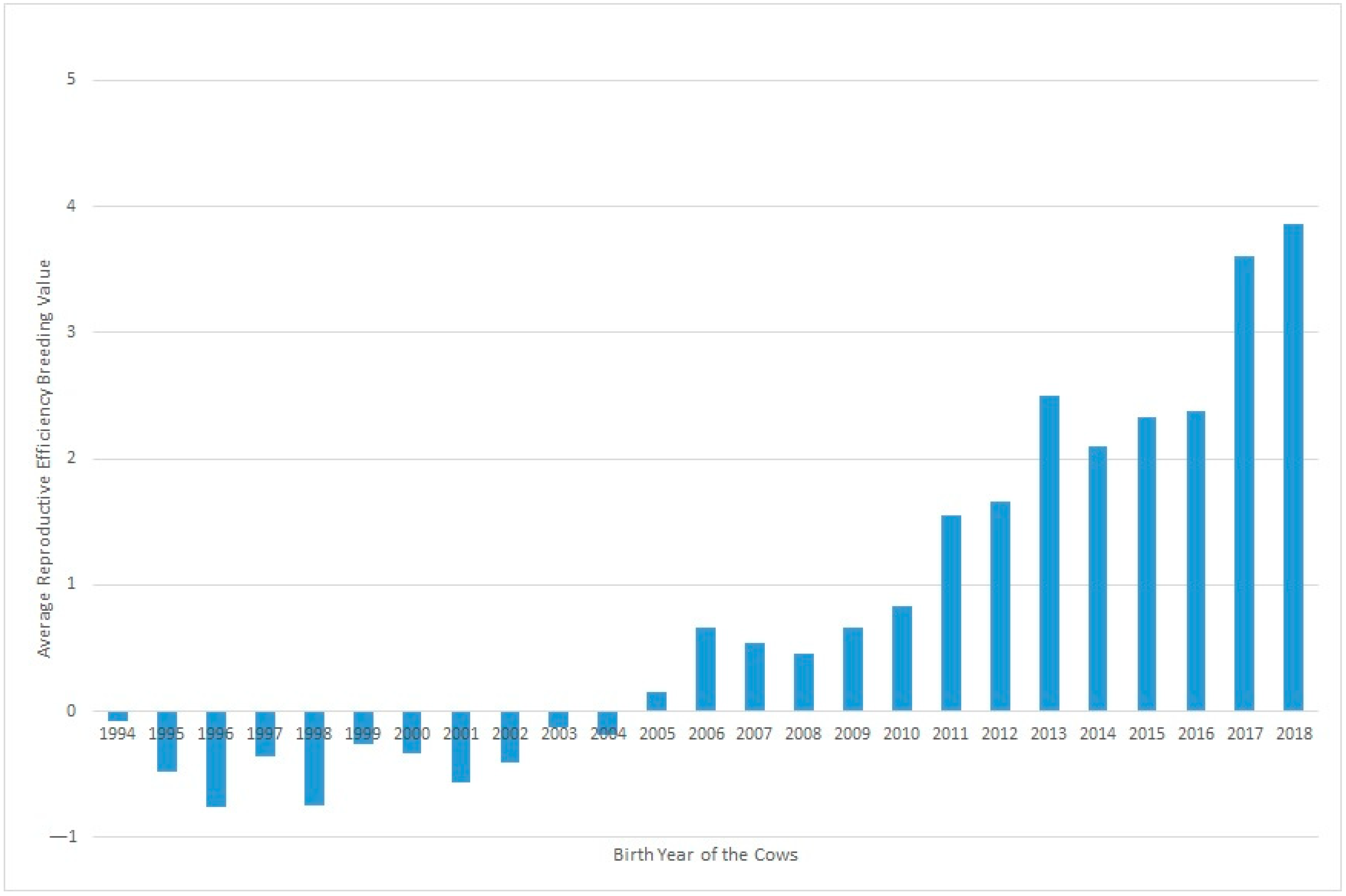
Figure 2 shows the evolution of Re over the years under study, where a clear positive trend in the breeding values can be seen over recent decades. This could be partly due to selection against carriers of the Robertsonian translocation 1/29 (rob(1;29)). Despite the incidence of carrier individuals in the whole population decresing during the last 30 years from 15.73% in 1992 to 1.06% in 2020, an increase was observed in Re phenotypic values in the same period. Although this increase in Re can be partially related to the decrease in the translocation rate, it may also be explained partly by other effects, such as improvements in animal management [
15].
3.2. Modeling Fertility across the Lifespan of the Cow
One of the drawbacks of the Rep model is the assumption of homogeneity of the variance components of Re across calvings. This hypothesis can be problematic for traits with low-moderate heritability, such as fertility-related traits. In contrast, RA models take into account the possible h
2 variations across the stage of production of the individual (the calving number of the cow, in our case) [
54]. In our study, both RA models showed better adjustments than those obtained using the Rep model (lower logL, AIC, and BIC,
Table 3), which supports the hypothesis that Re behavior is not homogeneous for each calving of the cow. Among the RA models, the second-order RA (RA2) showed the best adjustment to the data, suggesting that it can produce the most unbiased estimation of the (co)variance components.
In the RA model, heritabilities rose gradually with increased calving number, from 0.246 in the 1st calving to 0.583 in the 9th, mostly due to a slower pace of reduction observed in genetic variance in comparison with phenotypic variance (
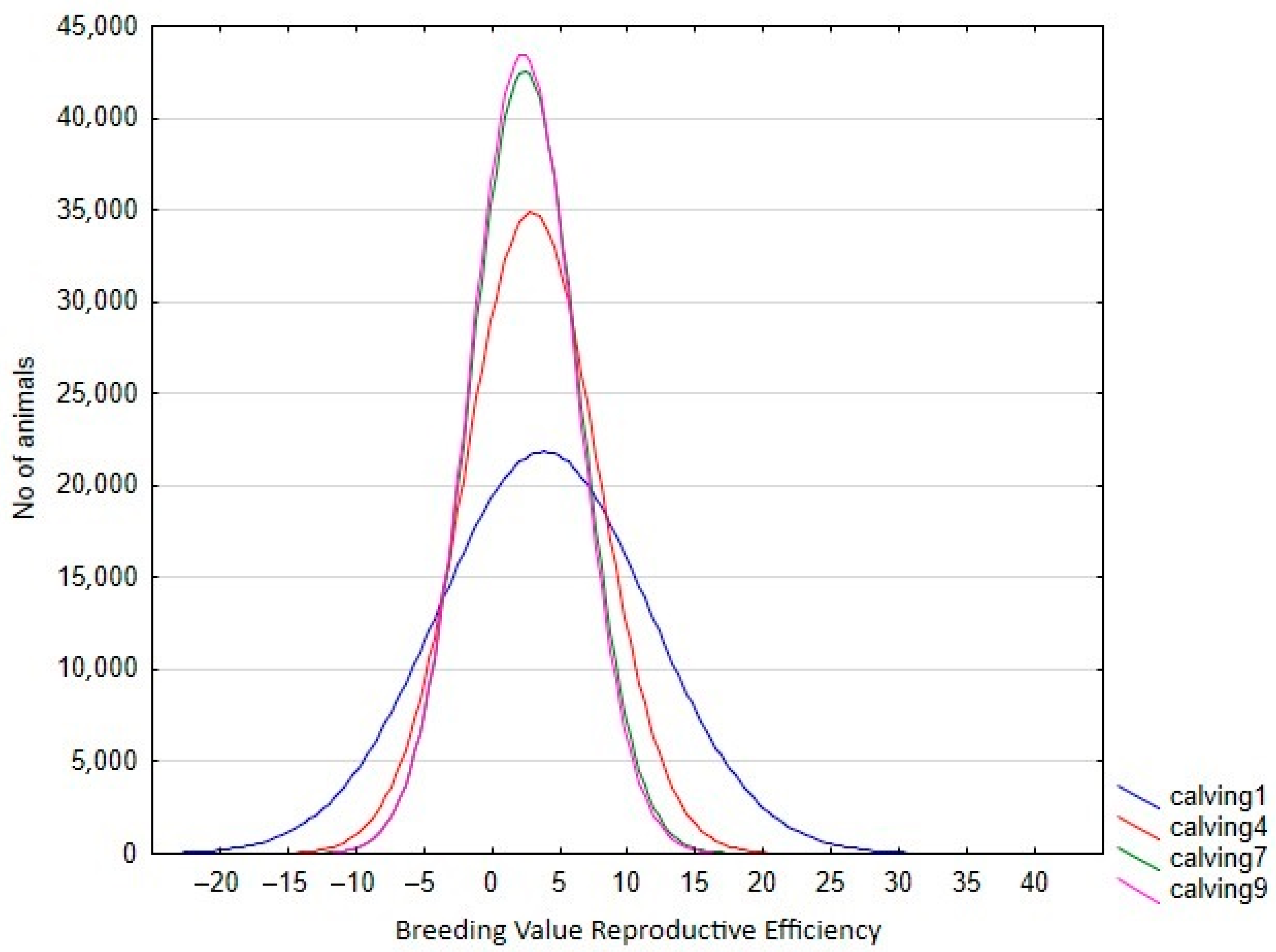
Table 4). In this context, only the estimated heritability at the first calving in RA was lower than the estimated value using the Rep model. Therefore, the use of RA models allows us to give a better explanation of the intrinsic variation of the trait, which is not possible using Re. However, the reduction in the total variance observed for the later calvings can limit the selection possibilities of the trait, despite the higher heritabilities observed. This can be seen clearly in
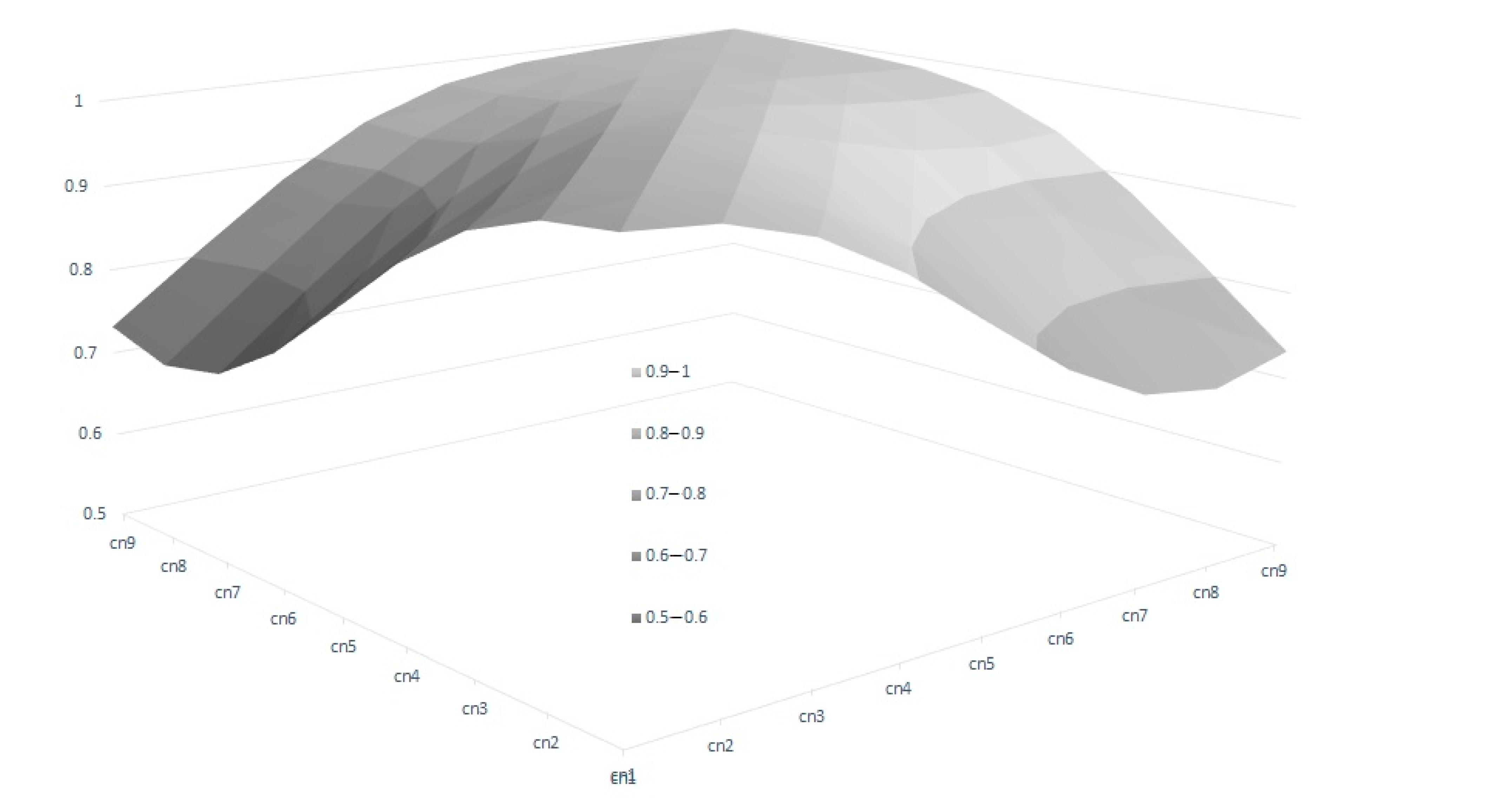
Figure 3, which shows the range of Re variation per calving. The size of this variation is close to 40% for the first calving but falls to 29% and 24% in the fourth and rest of the calvings, respectively, which implies that a better response to the selection based on EGVs is expected in the first calving. However, it is also worth mentioning that the correlated responses for the rest of the calvings will be lower, due to the decreasing pattern observed in the genetic correlations between the first and other calvings (
Figure 4), despite the fact that all the correlations estimated among the genetic values for different calvings were high (none were lower than 0.6,
Figure 4), and even higher (>0.83) when three or more calvings were considered. For this reason, our results suggest that the Re estimation for the third calving could be the best trait for selecting cows for fertility since it combines precocity in the selection practices, genetic variability, and reliability. In addition, it demonstrates that the order of merit for Re will vary among calvings, and therefore, selective practices should take this into account.
Finally, the dataset employed in this study allows us to analyze the evolution of EGV for Re during the last 3 decades, since the beginning of the improvement program (from 1992 to 2018,
Figure 5), which showed a positive trend towards increased Re genetic values in both sexes (7% on average). However, this value was higher in the EGV of the 1st and 3rd calving, which implies that breeding decisions based on fertility were made by breeders at the early stages of the lives of these cows and bulls.
3.3. Practical Implications of the RA Results
One of the problems of RA models is the vast amount of information produced in each analysis. In our case, we estimated 9 EGVs for 38,058 individuals, making it difficult to obtain reliable and clear conclusions. To cope with this situation, we performed a multivariate principal component analysis (PCA) in which all the available information was transformed into vectors (principal components, PC), which account for a certain % of the (co)variances existing among the variables studied. In this procedure, originally developed by Togashi and Lin [
44], PCs are ordered by the percentage of the total variance explained across all the response surfaces (all the calvings analyzed, in our case), in order to summarize the information produced by the RA analysis in a simple and useful way.
Our analysis revealed that 99.7% of EGV (co)variance was explained by two major
(PC1 and PC2), which demonstrates that all the variability determined by the RA analysis can be interpreted in a reliable way using this methodology (
Table 5). In our case, all the eigenvectors (EVs) in PC1 showed a very similar positive trend (close to 0.3), suggesting that any variation observed will be linear across the calvings (size vector). In contrast, PC2 (known as the vector of shape) showed positive values until the 4th calving, when it became negative.
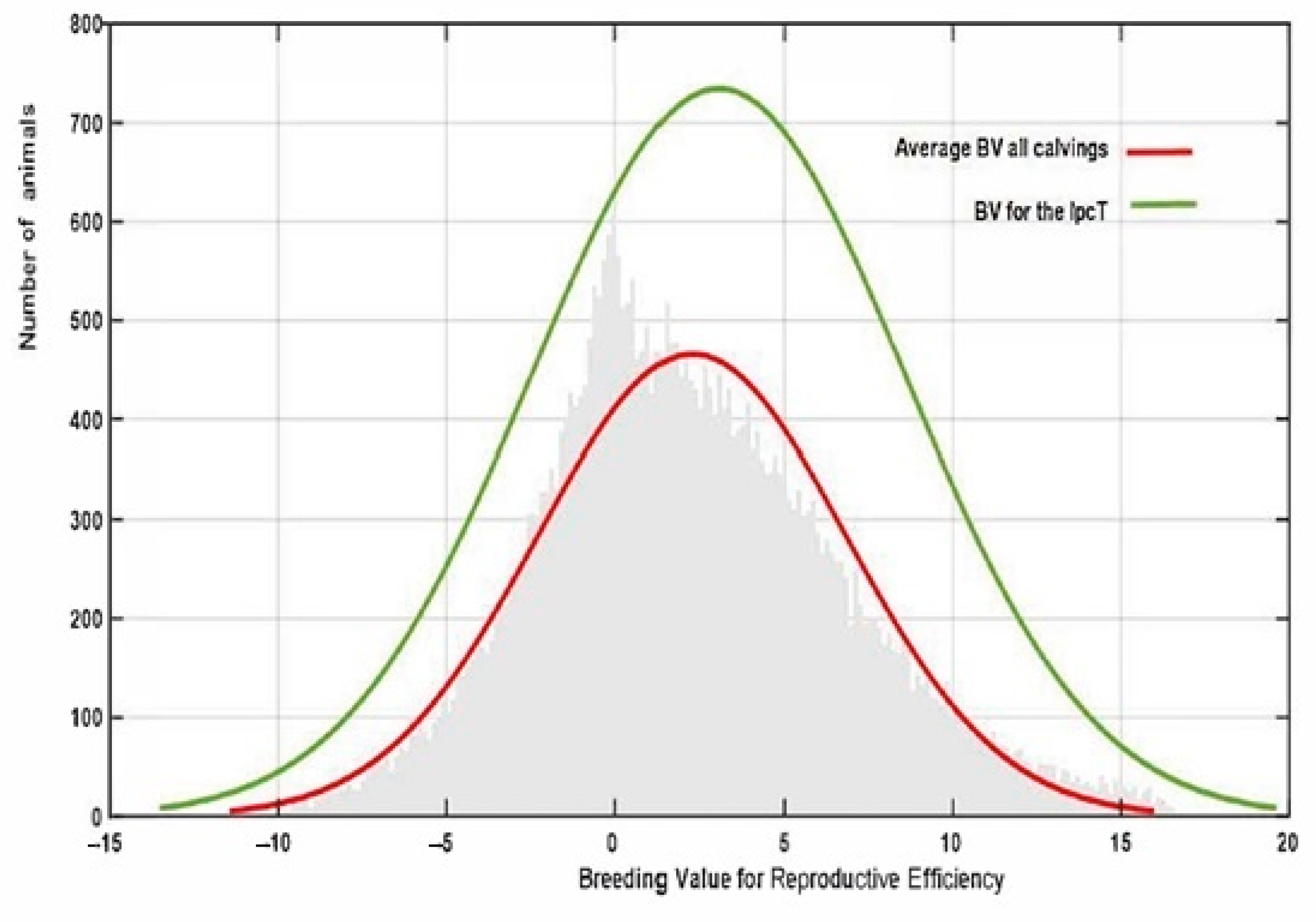
Finally, the estimated IpcT (calculated as the combination of Ipc values for PC1 and 2) showed a much more accurate fit to the breeding value distribution observed in all the population in comparison to that obtained by estimating the average EGV values for each individual using the RA model (
Figure 6). This value has an average of 0 ± 0.014 varying from −6.812 to 9.70. However, IpcT also allows us to summarize in a single value all the Re variability per individual across the calvings, thus allowing us to increase the accuracy of the breeding decisions. The correlation between the breeding values for Rep and IpcT shows a significant relationship of medium magnitude (0.45), which supports the use of this index as a pseudophenotype instead of the global EGV of the animal for this trait.
In addition, the use of IpcT permits a significant reduction in the risk of the possible interaction genotype x environment interaction problems (a different genetic potential in each calving number, as shown the
Table 5) by adding the eingvectors, which by definition are uncorrelated. The use of PCA in different species and breed selection [
43,
56,
57,
58] and the results of the correlations between the EGV of each calving and the IpcT demonstrate it (
Table 6).
3.4. Differences among Individuals in Re Development and Variation in Calvings
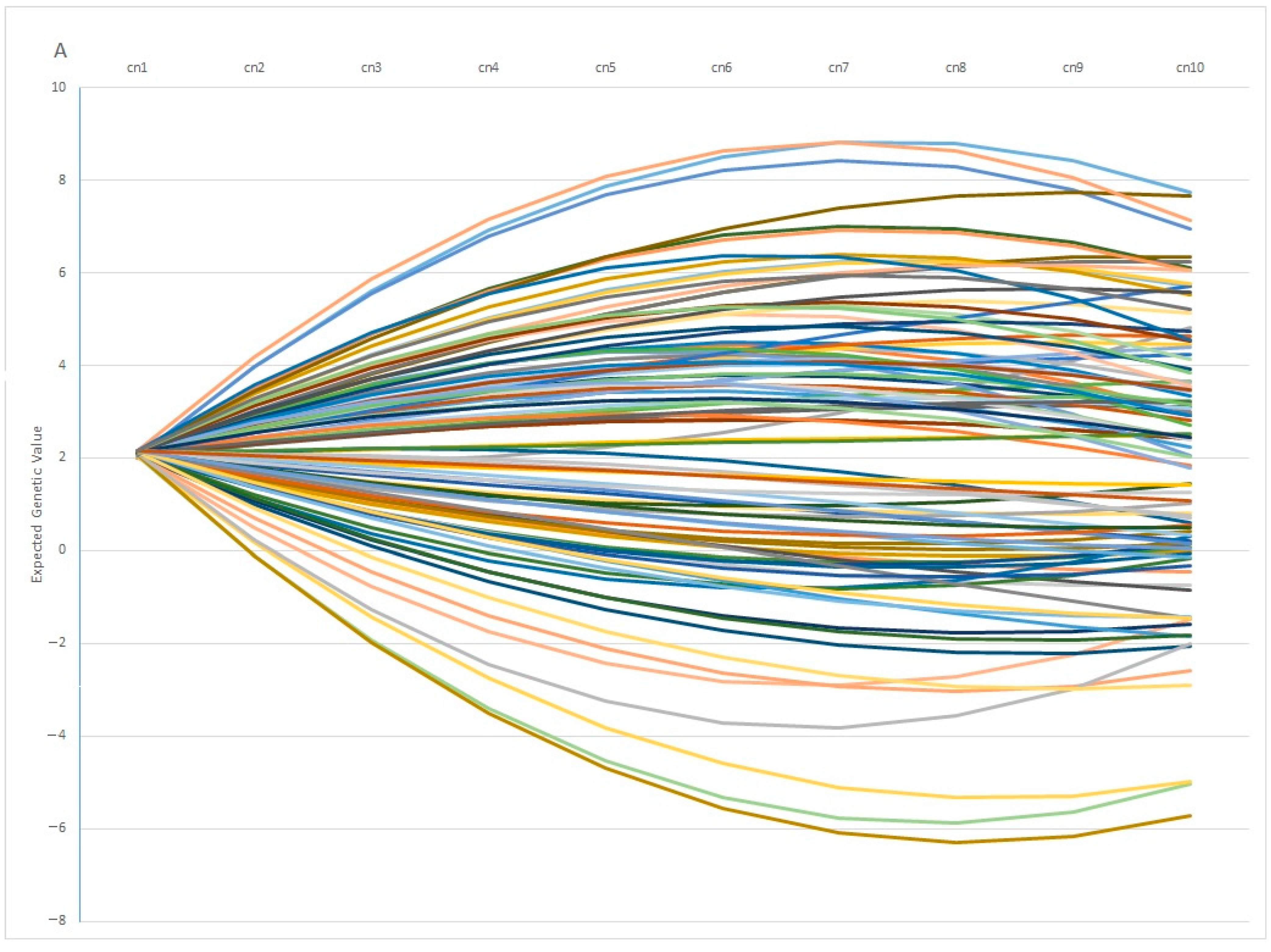
One interesting finding is the fact that individuals with similar EGV values at early ages can show opposite behaviors in terms of fertility at later stages. This pattern is observed in
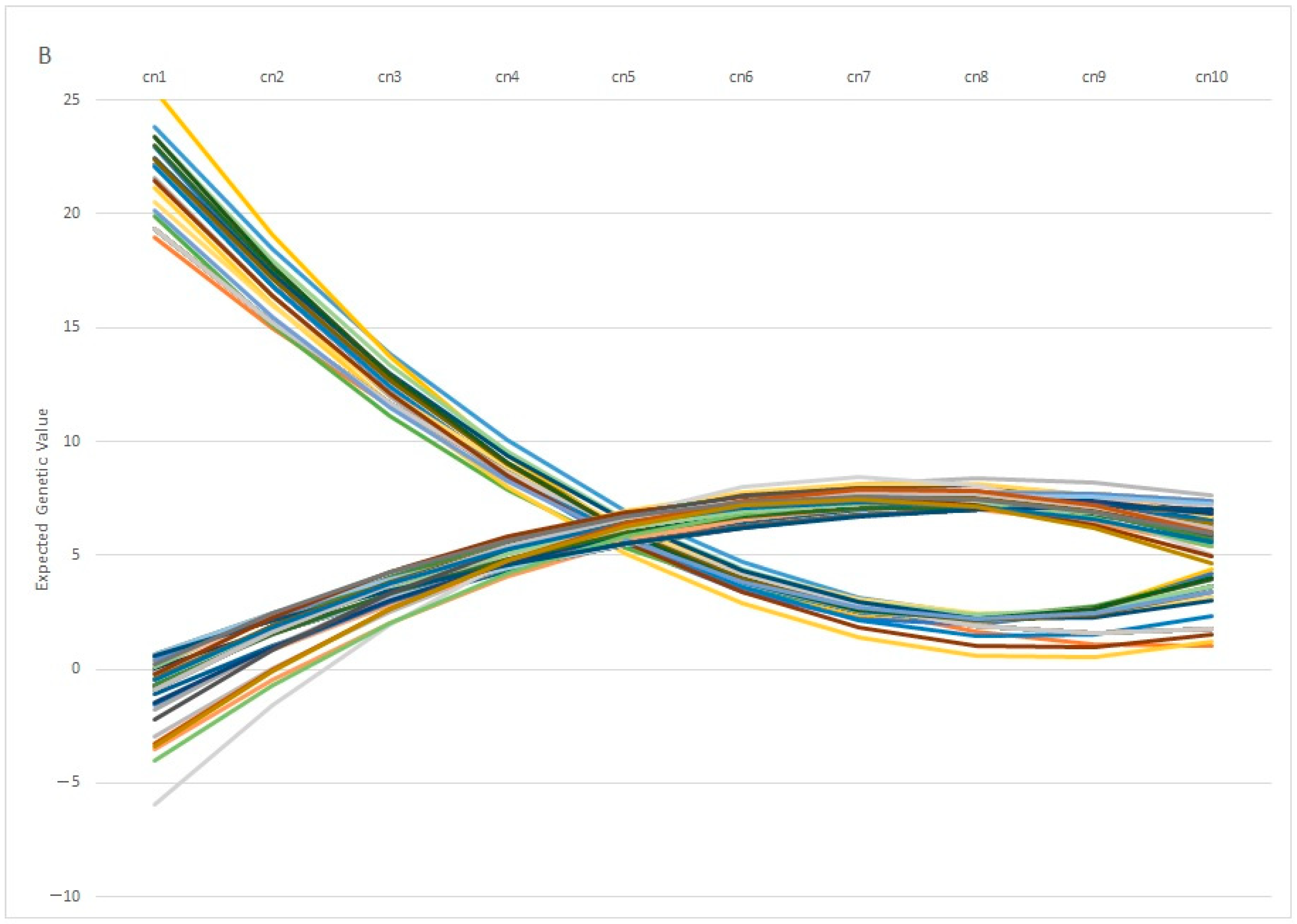
Figure 7A, in which 100 individuals with the same genetic estimation for Re at the first calving diverge profoundly at the fifth calving, in which some individuals showed a positive EGV while others showed the opposite. In the same way,
Figure 7B showed the Re estimation per calving of 100 individuals with the same value at the 5th calving, in which some of them showed a highly positive estimation at the 1st calving (with values ranging around 20%), whereas another group of individuals showed negative values at the same calving. From a biological point of view, the differences observed among individuals can be explained by the phenomenon known as “plasticity”, described by de Jong and Bijma [
59] and largely employed in modeling production traits in livestock systems [
60,
61]. However, it is worth mentioning that 82% of the best 100 individuals ranked using EGV1 and IpcT were coincident, and the divergent behaviors were shown by less of the population, which supports the use of PCA analysis as a robust methodology to perform better matings.
In our case, this parameter, defined as the difference between the genetic values obtained in the same character in the 1st and 9th calving, showed wide variability over the whole population. In addition, we determined an asymmetrical distribution with an increased proportion of negative values, which suggests increased reproductive efficiency at the first calving. We hypothesize that this deviation could be caused by the increased environmental variability affecting the Re values at the first calving caused by the age of the cow, since the age at first calving is influenced not only by the age at puberty but also by when each breeder decides to include the heifer in the mating lot. This breeder’s influence is reduced when the calving number increases (in addition to reducing the variability of the parameter with the elimination of less fertile animals).
3.5. Genome-Wide Association Study (GWAS) and Functional Analysis
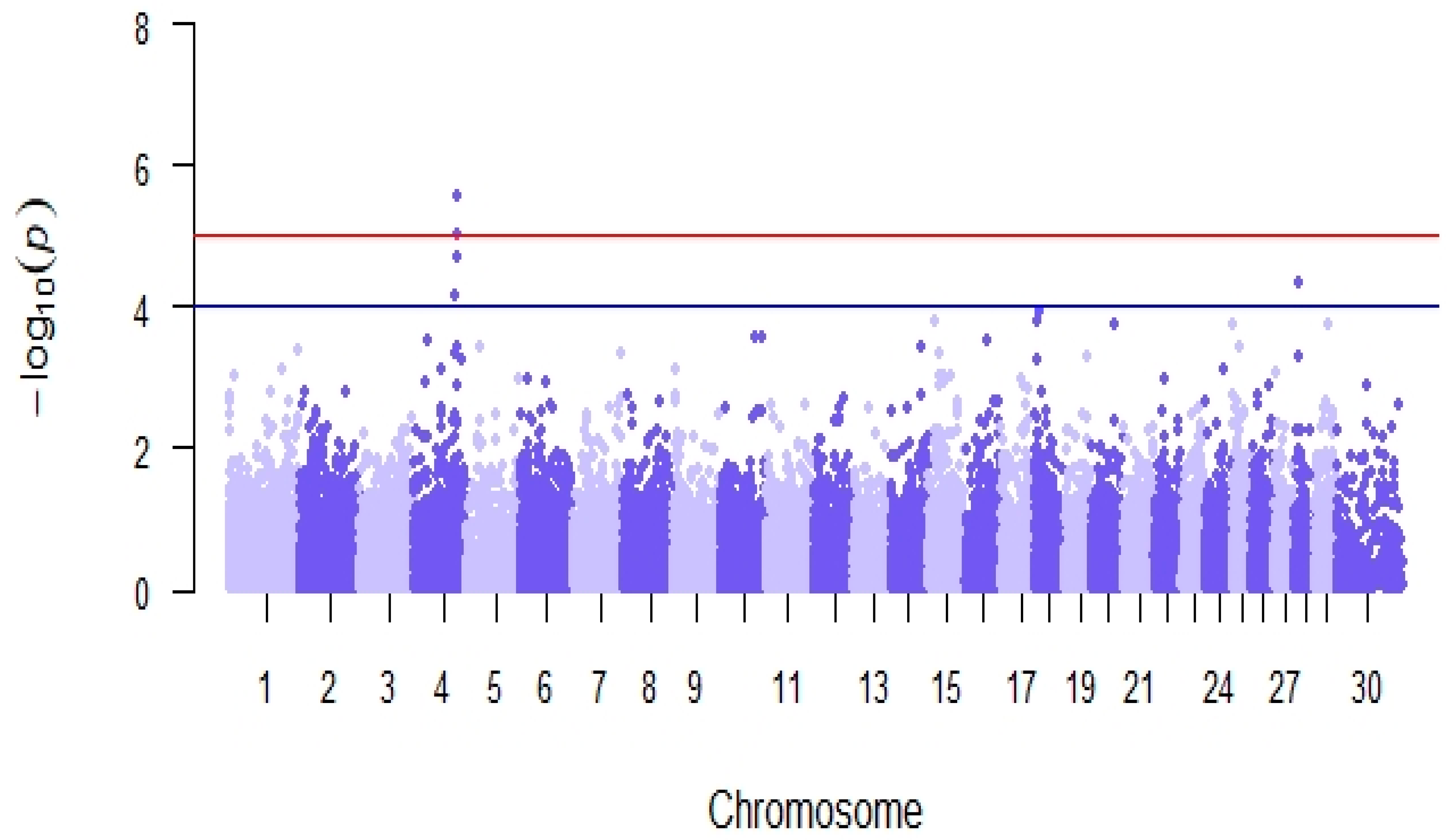
Genome-wide association analyses detected 5 SNPs significantly associated with reproductive efficiency (
p-value < 10
−4,
Table 7), located in 2 genomic regions on chromosomes BTA4 and BTA28 (
Figure 8). The estimated average effect of the significant SNPs in absolute values was 4.91. The in-silico functional analysis revealed the presence of 5 candidate genes (
NRF1, SSMEM1, CPA5, RYR2, and
ZP4) previously linked with known biological processes, molecular functions, and pathways related to fertility located within ± 500 Kb of the significant SNPs (
Table 7), among which there were processes related to steroidogenesis, embryonic development, and spermatogenesis in cattle, goat, rabbit, mouse, and human models.
The candidate genes located within the significant regions of the SNP AX-124382279 have previously been described in important biological pathways. One of these genes was nuclear respiratory factor 1 (
NRF1), which is involved in mitochondrial biogenesis, signal transduction, and protein synthesis. In mice, this gene has been related to late gestational embryonic lethality when a loss of function occurs [
62]. Recently, Zhang, et al. [
63] demonstrated the effects of
NRF1 on steroidogenesis and cell apoptosis in goat luteinized granulosa cells. An attenuated expression of
NRF1 led to mitochondrial dysfunction, disrupted the cellular redox balance, impaired steroid synthesis, and finally resulted in granulosa cell apoptosis through the mitochondria-dependent pathway. The other two candidate genes found in this region were
SSMEM1 and
CPA5, both related to male fertility. Serine-rich single-pass membrane protein 1 (
SSMEM1) is a conserved testis-specific gene in mammals. Nozawa, et al. [
64] demonstrated that
SSMEM1 is essential for male fertility in mice and found that the
SSMEM1 protein is expressed during spermatogenesis but not in mature sperm. The sterility of the
SSMEM1 KO mice was associated with globozoospermia and a loss of sperm motility, which is crucial for fertilization. Similarly,
CPA5 (carboxypeptidase A5) is involved in spermiogenesis and in the regulation of sperm function, as it is exclusively or highly expressed in spermatogenetic cells, especially from the secondary spermatocyte to elongated spermatid stages. However, Zhou, et al. [
65] reported that there were no perceptible changes in reproductive phenotypes in
CPA5-KO mice, suggesting that it is a dispensable factor for spermatogenesis and male fertility in mice.
On BTA28, we found the
RYR2 and
ZP4 genes located very close to the significant AX-169372743 marker.
RYR2 encodes an intracellular calcium-release channel and is expressed in many tissues, including the ovaries. In humans, some variants of
RYR2 were significantly associated with weight loss in early pregnancy [
66], providing evidence that it may play a role in fertility. However, its expression was also associated with amphiregulin, a key mediator of the effect of LH/hCG and a marker for oocyte competence [
67]. Finally, the
ZP4 gene codified a glycoprotein that formed the zona pellucida (ZP), the extracellular matrix that houses mammalian oocytes and embryos. In rabbits, Lamas-Toranzo, et al. [
68] reported that
ZP4 has a structural role in the zona pellucida and that rabbits without
ZP4 were subfertile. In vitro, the loss of
ZP4 did not affect ovulation, fertilization, or the early stages of the development of embryos. However, in vivo development was severely impaired in embryos covered by a ZP4-devoid zone. In addition, two recent studies in rats showed that
ZP4 is not responsible for gamete-specific interaction [
69,
70].

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}