

Transcriptome Analysis Reveals Effect of Dietary Probiotics on Immune Response Mechanism in Southern Catfish (Silurus meridionalis) in Response to Plesiomonas shigelloides
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Bacterial Strain
2.2. Animal and Experimental Design
2.3. RNA Extraction, Library Construction, and Sequencing
2.4. Data Processing and Assembly
2.5. Functional Annotation
2.6. Analysis of Differentially Expressed Genes (DEGs), Cluster Analysis, GO and KEGG Enrichment
2.7. Validation of DEGs by q-PCR
2.8. Validation of DEGs by ELISA
3. Results
3.1. Clinical Symptoms
3.2. Data Analysis of Transcriptome
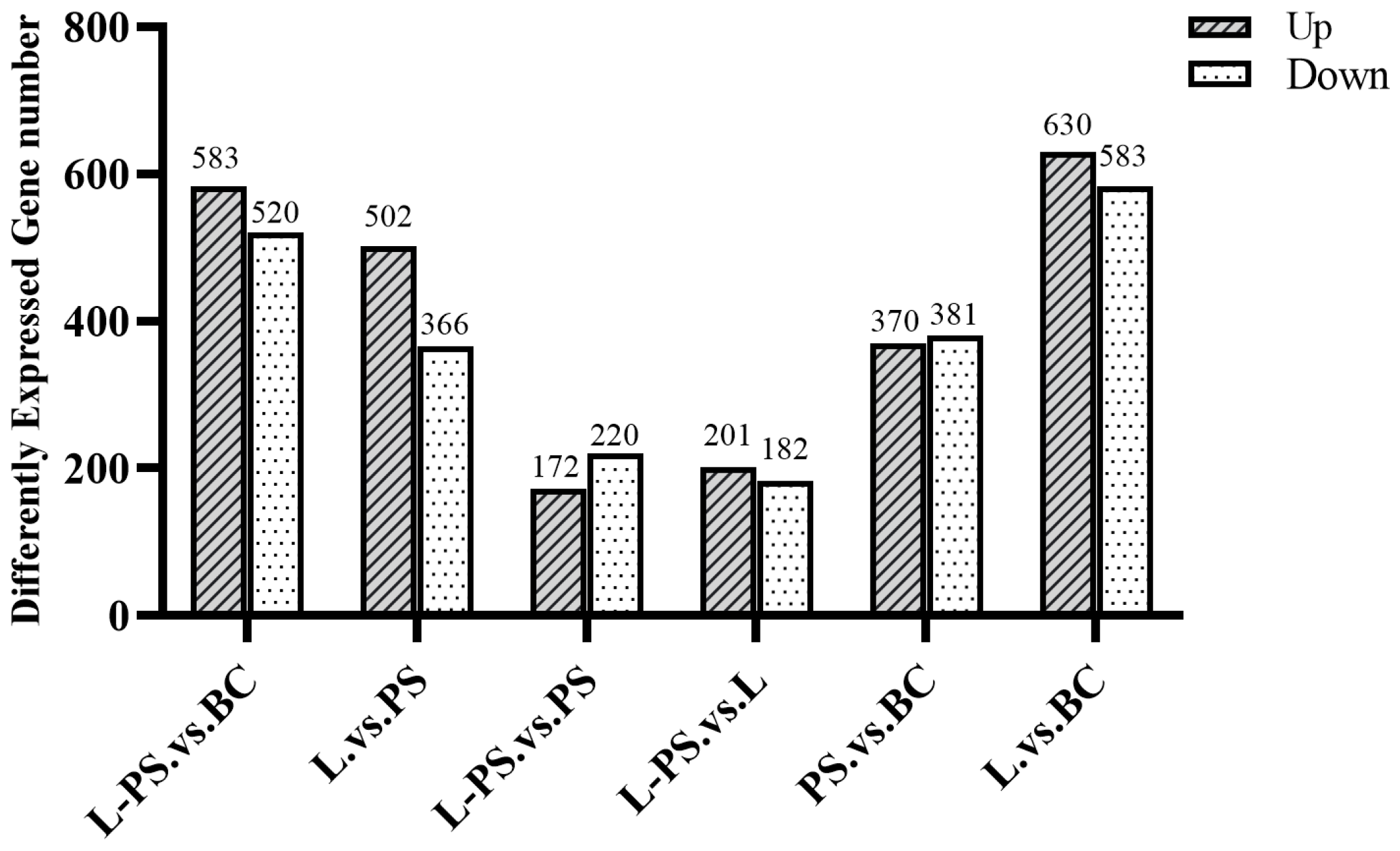
3.3. Differentially Expressed Genes (DEGs) Regulated by Exposure of P. shigelloides and Lactobacillus
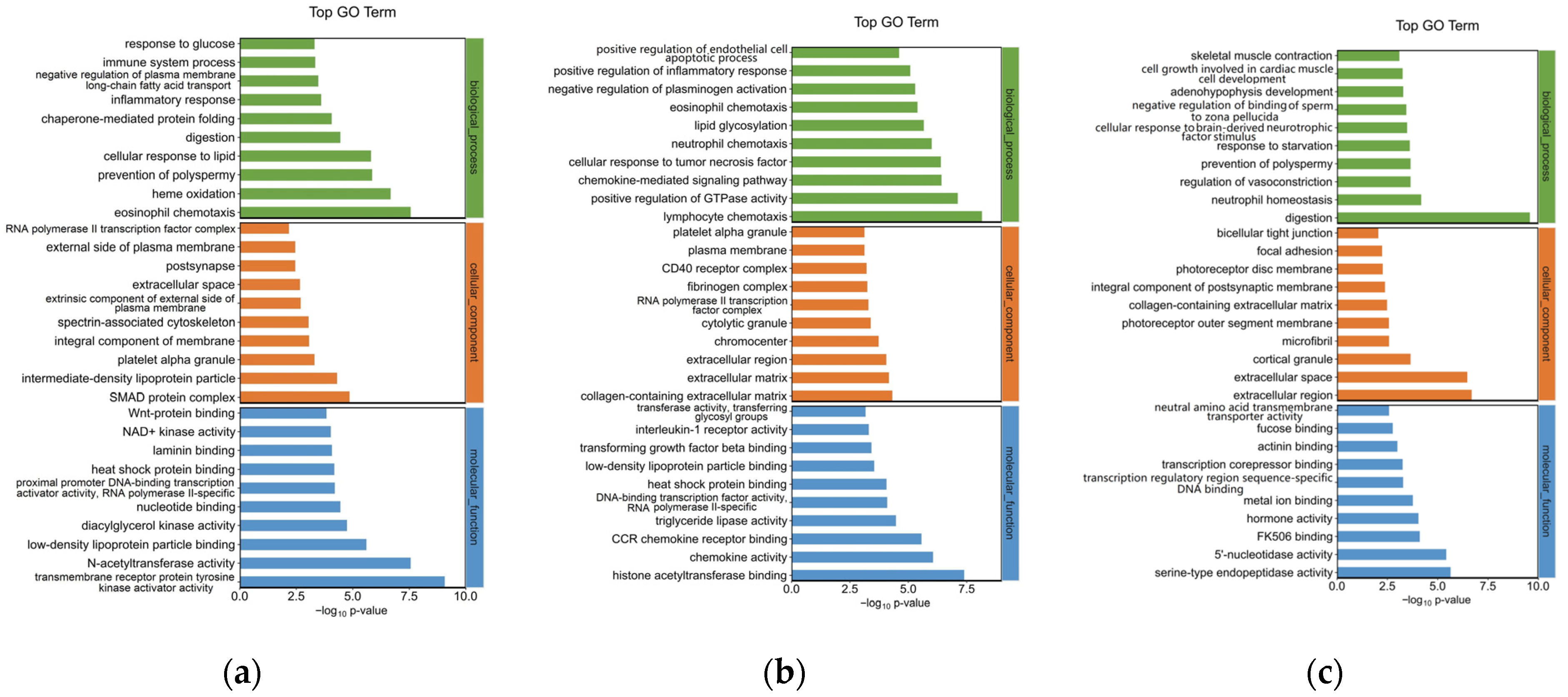
3.4. Gene Ontology Enrichment Analysis for DEGs
3.5. KEGG Analysis for DEGs
3.6. Analysis of Immune-Related Pathways
3.6.1. IL-17 Signalling Pathway
3.6.2. TNF Signalling Pathway
3.6.3. Toll-like Receptor Signalling Pathway
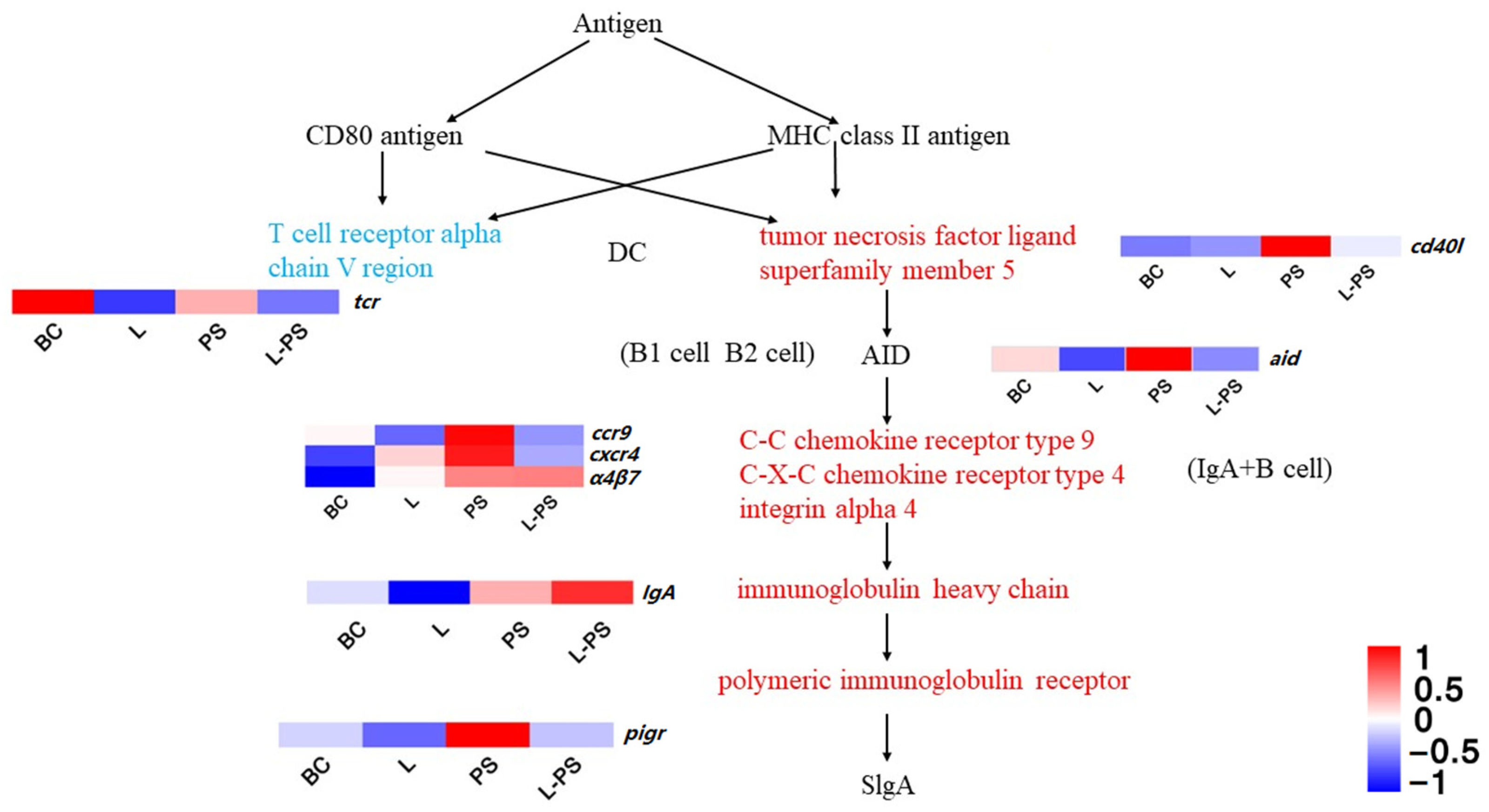
3.6.4. Intestinal Immune Network for IgA Production
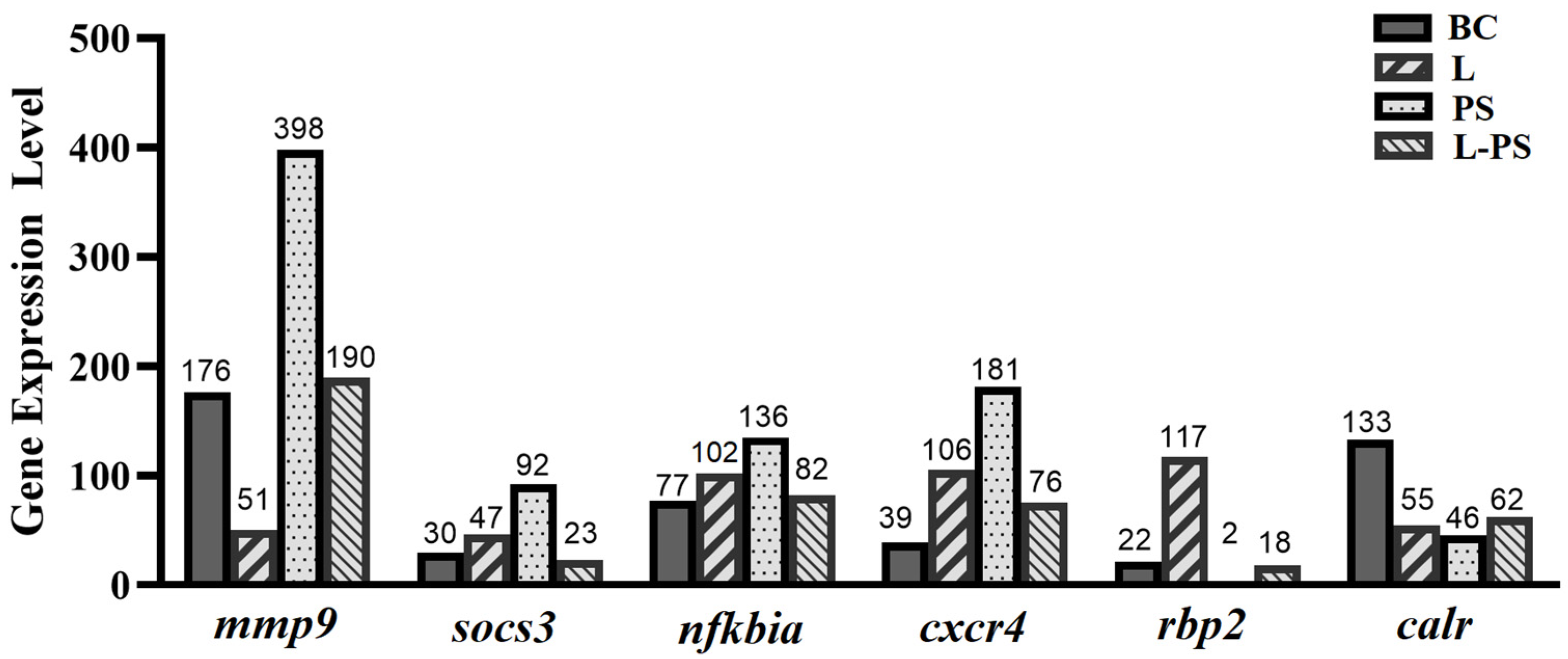
3.7. Validation of DEGs by qRT-PCR
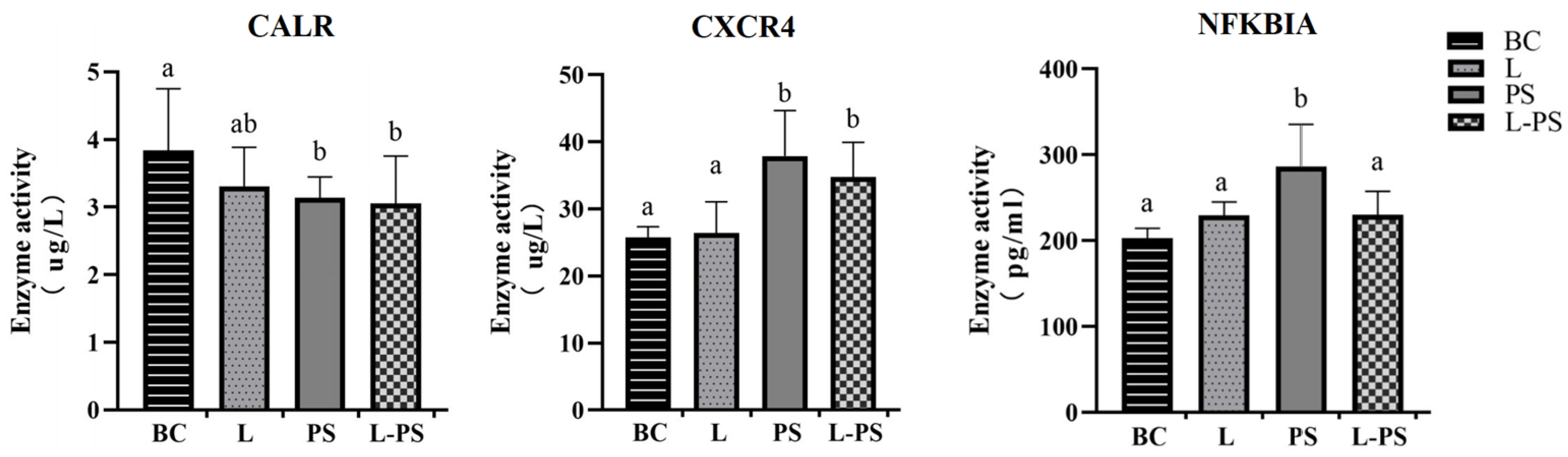
3.8. Validation of DEGs by ELISA
4. Discussion
4.1. Probiotic Complex Had Inhibitory Effects on Inflammatory Response Caused by P. shigelloides in Southern Catfish
4.1.1. IL-17 Signalling Pathway
4.1.2. TNF Signalling Pathway
4.1.3. Toll-like Receptor Signalling Pathway
4.1.4. Intestinal Immune Network for IgA Production
4.2. Other Related Key Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, R.A.; Oliva-Teles, A.; Pousão-Ferreira, P.; Jerusik, R.; Saavedra, M.J.; Enes, P.; Serra, C.R. Isolation and Characterization of Fish-Gut Bacillus spp. As Source of Natural Antimicrobial Compounds to Fight Aquaculture Bacterial Diseases. Mar. Biotechnol. 2021, 23, 276–293. [Google Scholar] [CrossRef]
- Liu, C.; Mai, K.; Zhang, W.; Chen, Q.; Leng, Y. Studies on the nutrition of two species of catfish, Silurus meridionalis Chen and S. asotus Linnaeus. I. Effects of dietary protein and lipid on growth performance and feed utilization. Aquaculture 2013, 404–405, 71–76. [Google Scholar] [CrossRef]
- Li, X.M.; Liu, L.; Yuan, J.M.; Xiao, Y.Y.; Fu, S.J.; Zhang, Y.G. The effect of aerobic exercise and starvation on growth performance and postprandial metabolic response in Juvenile southern catfish (Silurus meridionalis). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 193, 36–44. [Google Scholar] [CrossRef]
- Liu, W.; Qiu, H.; Yan, Y.; Xie, X. Acute Cd Toxicity, Metal Accumulation, and Ion Loss in Southern Catfish (Silurus meridionalis Chen). Toxics 2021, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.J.; Pang, X.; Cao, Z.D.; Peng, J.L.; Yan, G. The effects of fasting on the metabolic interaction between digestion and locomotion in Juvenile southern catfish (Silurus meridionalis). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.D.; Huang, Y.H.; Liang, J.Z.; Long, S.; Niu, Z.W.; Fan, H.L.; Huang, J.; Zeng, G.Z. Isolation, identification and drug sensitivity test of pathogenic bacteria from Silurus meridionalis infected with tail-rotted disease. J. South. Agric. 2015, 46, 1720–1725. [Google Scholar]
- Luo, Y.; Xie, X. Effects of temperature on the specific dynamic action of the southern catfish, Silurus meridionalis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 149, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Ai, Q.; Xie, X. Effects of replacement of fish meal by soybean meal and supplementation of methionine in fish meal/soybean meal-based diets on growth performance of the southern catfish Silurus meridionalis. J. World Aquac. Soc. 2005, 36, 498–507. [Google Scholar] [CrossRef]
- Fu, S.J.; Xie, X.J.; Cao, Z.D. Effect of dietary composition on specific dynamic action in southern catfish Silurus meridionalis Chen. Aquac. Res. 2005, 36, 1384–1390. [Google Scholar] [CrossRef]
- Li, Y.W.; Zhu, W.L.; Gu, J.R.; Wu, C.F.; Wu, J.; Xie, X.L.; Xu, H. Isolation of bdellovibrio infacting Silurus meridionalis pathogens and its biological characteristics. Freshw. Fish 2006, 36, 9–12. [Google Scholar]
- Cao, H.J.; Li, Y.W.; Lei, Y.; Wu, J.; Xu, H.; Zhang, T.; Zhang, X.J. Isolation, identification, phylogenetic analysis and related properties of a pathogen in Silurus meridionalis Chen. Acta Microbiol. Sin. 2007, 47, 1–6. [Google Scholar]
- Ji, L.L.; Wang, K.Y.; Xiao, D.; Yang, T. Isolation and identification of pathogenic bacteria causing ulcer disease of Silurus meriordinalis Chen. Freshw. Fish 2008, 38, 68–72. [Google Scholar]
- Chen, Z.; Cheng, K.; Zhou, X.; Zhang, Q. Pathogenic bacterium identification and histopathology of septicemia of Juvenile southern catfish, Silurus meridionalis. J. Fish Sci. China 2011, 18, 360–370. [Google Scholar]
- Li, C.W. Lsolation and Characterization of Edwardsiella ctalurid in South Catfish and Pathology and Immunohistochemistry after Experimental Infection; Sichuan Agricultural University: Chengdu, China, 2012. [Google Scholar]
- Jiang, L.P.; Liang, Q.L.; Li, Y.L.; She, R.; Wu, Z.W.; Qi, Z.L.; Zhang, J.L. Identification of pathogens in an outbreak deaths of southern catfish (Silurus meridionalis Chen) Juveniles. In Proceedings of the 2013 Annual Conference of the Chinese Fisheries Society, Sichuan, China. 2013, p. 117. Available online: https://en.cnki.com.cn/Article_en/CJFDTotal-DSYY201403010.htm (accessed on 23 October 2022).
- Guo, X. Isolation and Identification of Edwardsiellain South Catfish and Establishment of the Duplex PCR for Detection of Edwardsiella; Sichuan Agricultural University: Chengdu, China, 2014. [Google Scholar]
- Lin, C.Y.; Jin, B.Q.; Fan, Z.Z. A description of common diseases of southern largemouth catfish farmed in Tianjin. J. Hydroecology 2004, 24, 74–75. [Google Scholar]
- Wei, M.L.; Li, S.M.; Liang, J.Z. Isolation and identification and susceptibility test of pathogenic mild aeromonas in southern catfish. Guangxi, J. Anim. Husb. Vet. Med. 2020, 36, 195–198. [Google Scholar]
- Yasui, H.; Kiyoshima, J.; Hori, T.; Shida, K. Protection against influenza virus infection of mice fed Bifidobacterium breve YIT4064. Clin. Diagn. Lab. Immunol. 1999, 6, 186–192. [Google Scholar] [CrossRef]
- Agarwal, K.N.; Bhasin, S.K. Feasibility studies to control acute diarrhoea in children by feeding fermented milk preparations Actimel and Indian Dahi. Eur. J. Clin. Nutr. 2002, 56, S56–S59. [Google Scholar] [CrossRef]
- Pedone, C.A.; Arnaud, C.C.; Postaire, E.R.; Bouley, C.F.; Reinert, P. Multicentric study of the effect of milk fermented by Lactobacillus casei on the incidence of diarrhoea. Int. J. Clin. Pract. 2000, 54, 568–571. [Google Scholar]
- Pedone, C.A.; Bernabeu, A.O.; Postaire, E.R.; Bouley, C.F.; Reinert, P. The effect of supplementation with milk fermented by Lactobacillus casei (strain DN-114 001) on acute diarrhoea in children attending day care centres. Int. J. Clin. Pract. 1999, 53, 179–184. [Google Scholar]
- Szajewska, H.; Skórka, A.; Ruszczyński, M.; Gieruszczak-Białek, D. Meta-analysis: Lactobacillus GG for treating acute diarrhoea in children. Aliment. Pharmacol. Ther. 2007, 25, 871–881. [Google Scholar] [CrossRef]
- Segers, M.E.; Lebeer, S. Towards a better understanding of Lactobacillus rhamnosus GG—Host interactions. Microb. Cell Fact. 2014, 13, S7. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Zhang, X.; Hao, H.; Liu, Q.; Zhou, Z.; Liang, X.; Liu, T.; Gong, P.; Zhang, L.; Zhai, Z.; et al. Lactobacillus rhamnosus GG derived extracellular vesicles modulate gut microbiota and attenuate inflammatory in DSS-induced colitis mice. Nutrients 2021, 13, 3319. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, S.; Anjali, P.G.; Somashekaraiah, R.; Sreenivasa, M.Y. Probiotic properties of Lactobacillus casei–MYSRD 108 and Lactobacillus plantarum-MYSRD 71 with potential antimicrobial activity against Salmonella paratyphi. Biotechnol. Rep. 2021, 32, e00672. [Google Scholar] [CrossRef] [PubMed]
- Rengpipat, S.; Rueangruklikhit, T.; Piyatiratitivorakul, S. Evaluations of lactic acid bacteria as probiotics for Juvenile seabass Lates calcarifer. Aquac. Res. 2008, 39, 134–143. [Google Scholar] [CrossRef]
- Lee, J.S.; Cheng, H.; Damte, D.; Lee, S.J.; Kim, J.C.; Rhee, M.H.; Suh, J.W.; Park, S.C. Effects of dietary supplementation of Lactobacillus pentosus PL11 on the growth performance, immune and antioxidant systems of Japanese eel Anguilla japonica challenged with Edwardsiella tarda. Fish Shellfish. Immunol. 2013, 34, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Nikoskelainen, S.; Ouwehand, A.C.; Bylund, G.; Salminen, S.; Lilius, E.M. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish. Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Lin, H.L.; Shiu, Y.L.; Chiu, C.S.; Huang, S.L.; Liu, C.H. Screening probiotic candidates for a mixture of probiotics to enhance the growth performance, immunity, and disease resistance of Asian seabass, Lates calcarifer (Bloch), against Aeromonas hydrophila. Fish Shellfish. Immunol. 2017, 60, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.R.; Kim, D.; Jeon, J.; Lee, S.M.; Kim, H.K.; Kim, O.J.; Lee, J.I.; Suh, B.S.; Do, H.K.; Lee, K.H.; et al. The effects of combined dietary probiotics Lactococcus lactis BFE920 and Lactobacillus plantarum FGL0001 on innate immunity and disease resistance in olive flounder (Paralichthys olivaceus). Fish Shellfish. Immunol. 2015, 42, 177–183. [Google Scholar] [CrossRef]
- Feng, J.G.; Cai, Z.L.; Chen, Y.Y.; Chang, X.L.; Liu, H.F.; Liu, Y.J.; Zhang, J.X. Effects of an exopolysaccharide from Lactococcus lactis Q-9 on innate immune response, antioxidant activity, and disease resistance against Aeromonas hydrophila in Cyprinus carpio. J. Fish. China 2020, 44, 1477–1487. [Google Scholar] [CrossRef]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef]
- Memon, F.U.; Yang, Y.; Leghari, I.H.; Lv, F.; Soliman, A.M.; Zhang, W.; Si, H. Transcriptome Analysis Revealed Ameliorative Effects of Bacillus Based Probiotic on Immunity, Gut Barrier System, and Metabolism of Chicken under an Experimentally Induced Eimeria tenella Infection. Genes 2021, 12, 536. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Biotechnol. 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; Smyth, G.K. Moderated statistical tests for assessing differences in tag abundance. Bioinformatics 2007, 2, 2881–2887. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An r package for comparing biological themes among gene clusters. Omics-A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F. A cold-blooded view of adaptive immunity. Nat. Rev. Immunol. 2018, 18, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Miossec, P.; Kolls, J.K. Targeting IL-17 and TH17 cells in chronic inflammation. Nat. Rev. Drug Discov. 2012, 11, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Ritzmann, F.; Lunding, L.P.; Bals, R.; Wegmann, M.; Beisswenger, C. IL-17 Cytokines and Chronic Lung Diseases. Cells 2022, 11, 2132. [Google Scholar] [CrossRef]
- Ruiz de Morales, J.M.G.; Puig, L.; Daudén, E.; Cañete, J.D.; Pablos, J.L.; Martín, A.O.; Juanatey, C.G.; Adán, A.; Montalbán, X.; Borruel, N.; et al. Critical role of interleukin (IL)-17 in inflammatory and immune disorders: An updated review of the evidence focusing in controversies. Autoimmun. Rev. 2020, 19, 102429. [Google Scholar] [CrossRef]
- Amatya, N.; Garg, A.V.; Gaffen, S.L. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017, 38, 310–322. [Google Scholar] [CrossRef]
- Vandooren, J.; Van Den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Wu, L.; Wang, X.; He, X.; Li, Q.; Hua, Q.; Liu, R.; Qiu, Z. MMP9 Expression Correlates with Cisplatin Resistance in Small Cell Lung Cancer Patients. Front. Pharmacol. 2022, 13, 868203. [Google Scholar] [CrossRef]
- Xu, T.; Gao, S.; Liu, J.; Huang, Y.; Chen, K.; Zhang, X. MMP9 and IGFBP1 Regulate Tumor Immune and Drive Tumor Progression in Clear Cell Renal Cell Carcinoma. J. Cancer 2021, 12, 2243–2257. [Google Scholar] [CrossRef]
- Xu, L.; Cai, Z.; Yang, F.; Chen, M. Activation-induced upregulation of MMP9 in mast cells is a positive feedback mediator for mast cell activation. Mol. Med. Rep. 2017, 15, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zeng, H.; Gao, C. The role of neutrophil extracellular traps in central nervous system diseases and prospects for clinical application. Oxidative Med. Cell. Longev. 2021, 2021, 9931742. [Google Scholar] [CrossRef] [PubMed]
- Nakabo, S.; Romo-Tena, J.; Kaplan, M.J. Neutrophils as drivers of immune dysregulation in autoimmune diseases with skin manifestations. J. Investig. Dermatol. 2022, 142, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Amoras, E.; De Brito, W.B.; Queiroz, M.A.F.; Conde, S.; Vallinoto, I.M.V.C.; Ishak, R.; Vallinoto, A.C.R. The genetic profile and serum level of IL-8 are associated with chronic hepatitis B and C virus infection. Biomolecules 2021, 11, 1664. [Google Scholar] [CrossRef]
- Horiuchi, T.; Mitoma, H.; Harashima, S.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-alpha: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef]
- Hao, L.X.; Sun, L. Comparative analysis of the expression patterns of eight suppressors of cytokine signaling in tongue sole, Cynoglossus semilaevis. Fish Shellfish. Immunol. 2016, 55, 595–601. [Google Scholar] [CrossRef]
- Thanasaksiri, K.; Hirono, I.; Kondo, H. Identification and expression analysis of suppressors of cytokine signaling (SOCS) of Japanese flounder Paralichthys olivaceus. Fish Shellfish. Immunol. 2016, 58, 145–152. [Google Scholar] [CrossRef]
- Yao, J.; Mu, W.; Liu, S.; Zhang, J.; Wen, H.; Liu, Z. Identification, phylogeny and expression analysis of suppressors of cytokine signaling in channel catfish. Mol. Immunol. 2015, 64, 276–284. [Google Scholar] [CrossRef]
- Stoiber, D.; Stockinger, S.; Steinlein, P.; Kovarik, J.; Decker, T. Listeria monocytogenes modulates macrophage cytokine responses through STAT serine phosphorylation and the induction of suppressor of cytokine signaling 3. J. Immunol. 2001, 166, 466–472. [Google Scholar] [CrossRef]
- Kolesnichenko, M.; Mikuda, N.; Höpken, U.E.; Kärgel, E.; Uyar, B.; Tufan, A.B.; Milanovic, M.; Sun, W.; Krahn, I.; Schleich, K.; et al. Transcriptional repression of NFKBIA triggers constitutive IKK- and proteasome-independent p65/RelA activation in senescence. EMBO J. 2021, 40, e104296. [Google Scholar] [CrossRef]
- Ji, C. Transcriptome Analysis of Notch1a in Innate Immune Response in Zebrafish (Danio rerio) Larvae Challaged by Vibrio parahaemolyticus; Shanghai Ocean University: Shanghai, China, 2017. [Google Scholar]
- Song, T. Molecular Characteristics of RIG-I and MAVS of the Lamprey and their Signaling Pathways in Antiviral Immunity; Liaoning Normal University: Dalian, China, 2018. [Google Scholar]
- Tan, K.A. Transcriptomic Analysis and Determination of Immune-Related Genes in Channa Argus after Challenge with Nocardia Seriolae. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- Chen, J.; Qiao, Y.; Chen, G.; Chang, C.; Dong, H.; Tang, B.; Cheng, X.; Liu, X.; Hua, Z. Salmonella flagella confer anti-tumor immunological effect via activating Flagellin/TLR5 signalling within tumor microenvironment. Acta Pharm. Sin. B 2021, 11, 3165–3177. [Google Scholar] [CrossRef] [PubMed]
- Kantele, J.M.; Kurk, S.; Jutila, M.A. Effects of continuous exposure to stromal cell-derived factor-1 alpha on T cell rolling and tight adhesion to monolayers of activated endothelial cells. J. Immunol. 2000, 164, 5035–5040. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.M.; Nilsson, H.; Nyström, J.; Lindgren, D.; Leandersson, K.; Swärd, K.; Johansson, M.E. Localization and Regulation of Polymeric Ig Receptor in Healthy and Diseased Human Kidney. Am. J. Pathol. 2019, 189, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Maruthachalam, B.V.; Zwolak, A.; Lin-Schmidt, X.; Keough, E.; Tamot, N.; Venkataramani, S.; Geist, B.; Singh, S.; Ganesan, R. Discovery and characterization of single-domain antibodies for polymeric Ig receptor-mediated mucosal delivery of biologics. MAbs 2020, 12, 1708030. [Google Scholar] [CrossRef] [PubMed]
- Ohlmeier, S.; Mazur, W.; Linja-Aho, A.; Louhelainen, N.; Rönty, M.; Toljamo, T.; Bergmann, U.; Kinnula, V.L. Sputum proteomics identifies elevated PIGR levels in smokers and mild-to-moderate COPD. J. Proteome Res. 2012, 11, 599–608. [Google Scholar] [CrossRef]
- Hamuro, K.; Suetake, H.; Saha, N.R.; Kikuchi, K.; Suzuki, Y. A teleost polymeric Ig receptor exhibiting two Ig-like domains transports tetrameric IgM into the skin. J. Immunol. 2007, 178, 5682–5689. [Google Scholar] [CrossRef]
- Rombout, J.H.; van der Tuin, S.J.; Yang, G.; Schopman, N.; Mroczek, A.; Hermsen, T.; Taverne-Thiele, J.J. Expression of the polymeric Immunoglobulin Receptor (pIgR) in mucosal tissues of common carp (Cyprinus carpio L.). Fish Shellfish. Immunol. 2008, 24, 620–628. [Google Scholar] [CrossRef]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef]
- Ding, B.J.; Sheng, X.Y.; Tang, X.Q.; Xing, J.; Zhan, W.B. Molecular cloning and expression analysis of the plgR gene in Scophthalmus. J. Fish. Sci. China 2013, 20, 792–801. [Google Scholar]
- Kortum, A.N.; Rodriguez-Nunez, I.; Yang, J.; Shim, J.; Runft, D.; O’Driscoll, M.L.; Haire, R.N.; Cannon, J.P.; Turner, P.M.; Litman, R.T.; et al. Differential expression and ligand binding indicate alternative functions for zebrafish polymeric immunoglobulin receptor (pIgR) and a family of pIgR-like (PIGRL) proteins. Immunogenetics 2014, 66, 267–279. [Google Scholar] [CrossRef]
- Gold, L.I.; Eggleton, P.; Sweetwyne, M.T.; Van Duyn, L.B.; Greives, M.R.; Naylor, S.M.; Michalak, M.; Murphy-Ullrich, J.E. Calreticulin: Non-endoplasmic reticulum functions in physiology and disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 665–683. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.G.; Leach, M.R.; Brooke, K.W.M.; Wang, C.; Wheeler, L.W.; Hanly, E.K.; Rowley, C.W.; Levin, M.S.; Wagner, M.; Li, E.; et al. Epithelial expression of the cytosolic retinoid chaperone cellular retinol binding protein II is essential for in vivo imprinting of local gut dendritic cells by lumenal retinoids. Am. J. Pathol. 2012, 180, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Jaratlerdsiri, W.; Deakin, J.; Godinez, R.M.; Shan, X.; Peterson, D.G.; Marthey, S.; Lyons, E.; McCarthy, F.M.; Isberg, S.R.; Higgins, D.P.; et al. Comparative genome analyses reveal distinct structure in the saltwater crocodile MHC. PLoS ONE 2014, 9, e114631. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, D.; Wang, P.; Wu, W.; Fang, Q. Alox-5 as a potent therapeutic target on overcoming tki-resistance in chronic myeloid leukemia with t315i mutation in bcr-abl. Blood 2015, 126, 4835. [Google Scholar] [CrossRef]
- Connor, A.E.; Baumgartner, R.N.; Baumgartner, K.B.; Pinkston, C.M.; Boone, S.D.; John, E.M.; Torres-Mejía, G.; Hines, L.M.; Giuliano, A.R.; Wolff, R.K.; et al. Associations between ALOX, COX, and CRP polymorphisms and breast cancer among Hispanic and non-Hispanic white women: The breast cancer health disparities study. Mol. Carcinog. 2015, 54, 1541–1553. [Google Scholar] [CrossRef]
- Sun, Q.Y.; Zhou, H.H.; Mao, X.Y. Emerging Roles of 5-Lipoxygenase Phosphorylation in Inflammation and Cell Death. Oxidative Med. Cell. Longev. 2019, 2019, 2749173. [Google Scholar] [CrossRef]
- Lopes, D.E.M.; Jabr, C.L.; Decani, N.N.; Saraiva, A.C.; de Aquino, S.G.; Medeiros, A.I.; Rossa Junior, C. Inhibition of 5-lipoxygenase attenuates inflammation and BONE resorption in lipopolysaccharide-induced periodontal disease. J. Periodontol. 2017, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. A Publ. Protein Soc. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Forward Primer (5′->3′) | Reverse Primer (5′->3′) |
|---|---|---|
| 18s rRNA | GTGTCCGGTCCCTTTCAG | CAGGTATTCAGGCGAGTT |
| β-actin | CAGACGCTACTTCGAGTTT | CTCATCCATGGCGATGAAT |
| mmp9 | TGAATTGTCCCAACTACCGAA | ACATCTGGGATTTGCATTTAAG |
| socs3 | CAGTACCCAAATGACTAGGAAC | CCCTGAAAGGCATAATGCT |
| nfkbia | ACACCAAACTACAACGGTC | CGTTACACTGCTCCTGTTC |
| cxcr4 | CCGTTCCTGATCTGGTCTT | TTGGCTTCGTGAGGGTAG |
| rbp2 | GTGCGCAATGAGAACTTTGA | GTTTGTGTGAGGTGTGCT |
| calr | TCAACTCGGTTTGAGGACTT | ATCCTCCTCCACAGTCTATG |
| Group | Number | Patients | Prevalence |
|---|---|---|---|
| BC | 20 | 0 | 0% |
| L | 20 | 0 | 0% |
| PS | 20 | 17 | 85% |
| L-PS | 20 | 3 | 15% |
| Sample | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Valid Bases | Q30 | GC |
|---|---|---|---|---|---|---|---|
| BC1 | 48.21M | 7.28G | 47.13M | 6.53G | 89.73% | 93.87% | 49.29% |
| BC2 | 49.20M | 7.43G | 48.15M | 6.69G | 90.07% | 93.99% | 49.52% |
| BC3 | 47.87M | 7.23G | 46.83M | 6.51G | 90.09% | 93.88% | 49.21% |
| L1 | 48.63M | 7.34G | 47.53M | 6.60G | 89.91% | 93.81% | 49.44% |
| L2 | 49.67M | 7.50G | 48.63M | 6.71G | 89.44% | 94.07% | 49.81% |
| L3 | 50.32M | 7.60G | 49.25M | 6.86G | 90.24% | 93.92% | 49.12% |
| L_PS1 | 49.03M | 7.40G | 47.75M | 6.49G | 87.59% | 93.39% | 49.02% |
| L_PS2 | 49.34M | 7.45G | 48.05M | 6.53G | 87.69% | 93.36% | 49.06% |
| L_PS3 | 49.59M | 7.49G | 48.26M | 6.55G | 87.42% | 93.23% | 49.30% |
| PS1 | 51.08M | 7.66G | 49.66M | 6.70G | 87.46% | 93.15% | 49.43% |
| PS2 | 48.48M | 7.32G | 47.13M | 6.43G | 87.82% | 93.64% | 49.41% |
| PS3 | 51.37M | 7.76G | 49.95M | 6.77G | 87.32% | 93.36% | 49.15% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Qian, J.; Ji, D.; Liu, X.; Dong, R. Transcriptome Analysis Reveals Effect of Dietary Probiotics on Immune Response Mechanism in Southern Catfish (Silurus meridionalis) in Response to Plesiomonas shigelloides. Animals 2023, 13, 449. https://doi.org/10.3390/ani13030449
Wang R, Qian J, Ji D, Liu X, Dong R. Transcriptome Analysis Reveals Effect of Dietary Probiotics on Immune Response Mechanism in Southern Catfish (Silurus meridionalis) in Response to Plesiomonas shigelloides. Animals. 2023; 13(3):449. https://doi.org/10.3390/ani13030449
Chicago/Turabian StyleWang, Rongrong, Jiaming Qian, Da Ji, Xingyu Liu, and Ranran Dong. 2023. "Transcriptome Analysis Reveals Effect of Dietary Probiotics on Immune Response Mechanism in Southern Catfish (Silurus meridionalis) in Response to Plesiomonas shigelloides" Animals 13, no. 3: 449. https://doi.org/10.3390/ani13030449
APA StyleWang, R., Qian, J., Ji, D., Liu, X., & Dong, R. (2023). Transcriptome Analysis Reveals Effect of Dietary Probiotics on Immune Response Mechanism in Southern Catfish (Silurus meridionalis) in Response to Plesiomonas shigelloides. Animals, 13(3), 449. https://doi.org/10.3390/ani13030449