
Clicker Training as an Applied Refinement Measure in Chickens

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Training
2.4. Training Procedure
2.4.1. Reward Food and Clicker–Reward Combination
2.4.2. Influence of Chicken Breed and Individual Character on Training
2.4.3. Group Training
2.4.4. Single Training
2.5. Sample Collection, Sample Preparation, and Description of the General Examination as the Induced Stress Situation
2.6. Corticosterone Measurement
2.6.1. Verification of Corticosterone ELISA
2.6.2. Extraction of Plasma Corticosterone
2.6.3. Dilution of Saliva Sample
2.7. New Area Test—Behavior and Fecal Matter
2.8. Statistical Evaluation
3. Results
3.1. Clicker Training
3.1.1. Group Training
3.1.2. Single Training
3.1.3. Character Types, Description of the Character, and Their Frequency in This Study
- Type 1—courageous and active (frequency: often (9 of 21 animals))
- Type 2—courageous and accepting (frequency: often (8 of 21 animals))
- Type 3—fearful and active (frequency: rare (3 of 21 animals))
- Type 4—fearful (frequency: very rare (1 of 21 animals))
3.2. Corticosterone ELISA
3.2.1. Verification of Corticosterone ELISA for Plasma
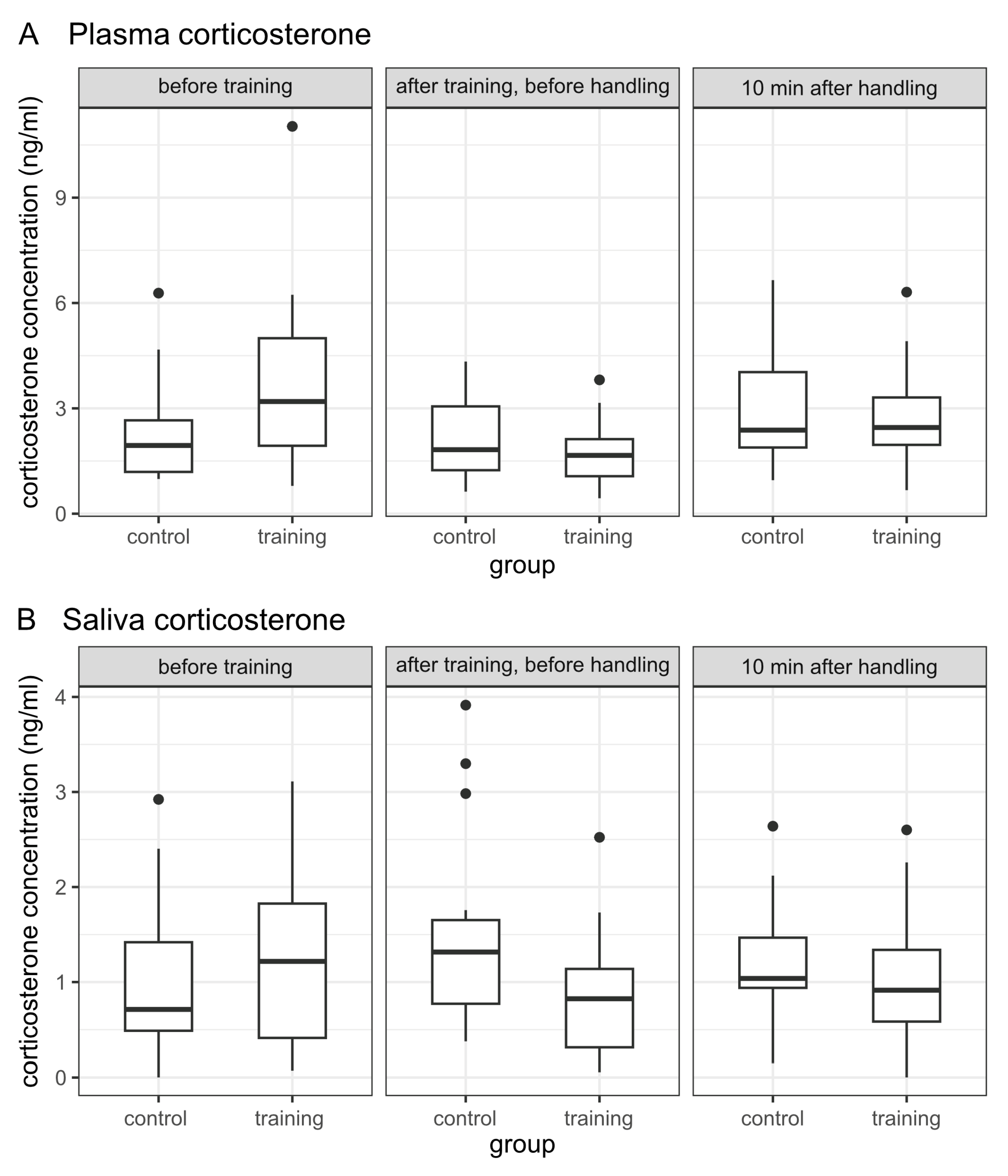
3.2.2. Corticosterone Concentrations in Plasma and Saliva
3.3. New Area Test Behavior
- -
- One animal: disturbance by crowing rooster in a neighboring stable.
- -
- Two animals: disturbance by crowing rooster in a neighboring stable.
- -
- Two animals: temporarily in unavoidable blind spot of camera.
3.4. New Area Test—Defecation
4. Discussion
4.1. Verification of Corticosterone ELISA
4.2. Stress Measurement Using Plasma and Salivary Corticosterone
4.3. New Area Test
4.3.1. Shaking as a Reordering Behavior
4.3.2. Locomotion
4.3.3. New Area Test—Defecation
4.4. Cannibalism and the Role of Active Cognitive Enrichment
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Angelis, I.; Ricceri, L.; Vitale, A. The 3R principle: 60 years taken well. Preface. Ann. Ist. Super. Sanita. 2019, 55, 398–399. [Google Scholar] [CrossRef]
- Schlüter, K.D. Tierversuche: Übertragbarkeit auf den Menschen und ihre Durchführbarkeit. Z. Für Herz-Thorax-Und Gefäßchirurgie 2019, 33, 67–71. [Google Scholar] [CrossRef]
- Leidinger, C.; Herrmann, F.; Thöne-Reineke, C.; Baumgart, N.; Baumgart, J. Introducing clicker training as a cognitive enrichment for laboratory mice. J. Vis. Exp. 2017, 6, 55415. [Google Scholar] [CrossRef]
- O’Connor, M.D. The 3R principle: Advancing clinical application of human pluripotent stem cells. Stem Cell Res. Ther. 2013, 4, 21. [Google Scholar] [CrossRef]
- Erkekoglu, P.; Giray, B.K.; Basran, N. 3R principle and alternative toxicity testing methods. Fabad J. Pharm. Sci. 2011, 36, 101–117. Available online: http://dergi.fabad.org.tr/pdf/volum36/issue2/101-117.pdf (accessed on 17 May 2020).
- Manciocco, A.; Chiarotti, F.; Vitale, A.; Calamandrei, G.; Laviola, G.; Alleva, E. The application of Russell and Burch 3R principle in rodent models of neurodegenerative disease: The case of Parkinson’s disease. Neurosci. Biobehav. Rev. 2009, 33, 18–32. [Google Scholar] [CrossRef]
- Spielmann, H. A chair on alternatives? ALTEX 2002, 19, 69–73. Available online: https://altex.org/index.php/altex/article/view/1105/1099 (accessed on 25 January 2023). [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes Text with EEA Relevance; ELI. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 15 October 2021).
- The German Animal Welfare Act; in the Version Published on 18 May 2006 (Federal Law Gazette I p. 1206, 1313), as Last Amended by Article 2(20) of the Act of 20 December 2022 (Federal Law Gazette I p. 2752). Available online: https://www.gesetze-im-internet.de/tierschg/BJNR012770972.html (accessed on 22 July 2023).
- The German Animal Protection Experimental Animal Ordinance of 1 August 2013 (Federal Law Gazette I p. 3125, 3126), as Last Amended by Article 1 of the Ordinance of 11 August 2021 (Federal Law Gazette I p. 3570). Available online: https://www.gesetze-im-internet.de/tierschversv/BJNR312600013.html (accessed on 7 May 2023).
- NC3Rs (National Centre for the Replacement, Refinement and Reduction of Animals in Research). Available online: https://nc3rs.org.uk/who-we-are/3rs (accessed on 14 September 2023).
- Flecknell, P. Replacement, Reduction, Refinement. ALTEX 2002, 19, 73–78. [Google Scholar] [PubMed]
- Macartney, E.L.; Lagisz, M.; Nakagawa, S. The relative benefits of environmental enrichment on learning and memory are greater when stressed: A meta-analysis of interactions in rodents. Neurosci. Biobehav. Rev. 2022, 135, 104554. [Google Scholar] [CrossRef]
- Baumans, V.; Van Loo, P.L.P. How to improve housing conditions of laboratory animals: The possibilities of environmental refinement. Vet. J. 2013, 195, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.H.; Reed, B.T.; Hawkins, P. Enrichment for laboratory zebrafish—A review of the evidence and the challenges. Animals 2021, 11, 698. [Google Scholar] [CrossRef]
- Gillis, T.E.; Janes, A.C.; Kaufman, M.J. Positive reinforcement training in squirrel monkeys using clicker training. Am. J. Primatol. 2012, 74, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Laule, G.E.; Bloomsmith, M.A.; Schapiro, S.J. The use of positive reinforcement training techniques to enhance the care, management, and welfare of primates in the laboratory. J. Appl. Anim. Welf. Sci. 2003, 6, 163–173. [Google Scholar] [CrossRef]
- Kogan, L.; Kolus, C.; Schoenfeld-Tacher, R. Assessment of clicker training for shelter cats. Animals 2017, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Ellis, S.; Greening, L. Positively reinforcing an operant task using tactile stimulation and food—A comparison in horses using clicker training. J. Vet. Behav. Clin. Appl. Res. 2016, 15, 78. [Google Scholar] [CrossRef]
- Strychalski, J.; Gugołek, A.; Konstantynowicz, M. Clicker training efficiency in shaping the desired behaviour in the following dog breeds: Boxer, chow chow and yorkshire terrier. Pol. J. Nat. Sc. 2015, 30, 235–243. Available online: https://wmbc.olsztyn.pl/Content/1663/Nat30_3.pdf#page=23 (accessed on 27 May 2021).
- Jønholt, L.; Bundgaard, C.J.; Carlsen, M.; Sørensen, D.B. A Case Study on the Behavioural Effect of Positive Reinforcement Training in a Novel Task Participation Test in Göttingen Mini Pigs. Animals 2021, 11, 1610. [Google Scholar] [CrossRef] [PubMed]
- Callealta, I.; Lueders, I.; Luther-Binoir, I.; Ganswindt, A. Positive reinforcement conditioning as a tool for frequent minimally invasive blood and vaginal swab sampling in African lions (Panthera leo). J. Appl. Anim. Welf. Sci. 2020, 23, 508–519. [Google Scholar] [CrossRef]
- Armbrecht, Y.; Wilkens, M. Stressfreies Handling und Medical Training. Team Konkret 2016, 12, 20–23. [Google Scholar] [CrossRef]
- Dadone, L.I.; Schilz, A.; Friedman, S.G.; Bredahl, J.; Foxworth, S.; Chastain, B. Training giraffe (Giraffa camelopardalis reticulata) for front foot radiographs and hoof care. Zoo Biol. 2016, 35, 228–236. [Google Scholar] [CrossRef]
- Weiss, E.; Wilson, S. The use of classical and operant conditioning in training Aldabra tortoises (Geochelone gigantea) for venipuncture and other husbandry issues. J. Appl. Anim. Welf. Sci. 2003, 6, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Dudde, A.; Krause, E.T.; Matthews, L.R.; Schrader, L. More than eggs–relationship between productivity and learning in laying hens. Front. Psychol. 2018, 9, 2000. [Google Scholar] [CrossRef]
- Chiandetti, C.; Avella, S.; Fongaro, E.; Cerri, F. Can clicker training facilitate conditioning in dogs? Appl. Anim. Behav. Sci. 2016, 184, 109–116. [Google Scholar] [CrossRef]
- Vandbakk, M.; Olaff, H.S.; Holth, P. Conditioned reinforcement: The effectiveness of stimulus—Stimulus pairing and operant discrimination procedures. Psychol. Rec. 2019, 69, 67–81. [Google Scholar] [CrossRef]
- Peng, R.C.; Yan, W.R. An automated animal training apparatus for operant conditioning studies in rodents. In Proceedings of the 2019 8th International Conference on Bioinformatics and Biomedical Science, Beijing, China, 23–25 October 2019; pp. 21–26. [Google Scholar] [CrossRef]
- Pierce, W.D.; Cheney, C.D. Behavior Analysis and Learning, 6th ed.; Routledge: New York, NY, USA, 2017; pp. 1–613. ISBN 978-1138898585. [Google Scholar]
- European Union Animal Research Statistics 2020. Available online: https://www.understandinganimalresearch.org.uk/news/eu-wide-animal-research-statistics-2020 (accessed on 14 November 2023).
- Annual Statistics of Animals Used in Great Britain. Available online: https://www.understandinganimalresearch.org.uk/what-is-animal-research/numbers-animals/annual-statistics-of-animals-used-in-great-britain#2022. (accessed on 14 November 2023).
- Animaltestinfo. The German Federal Institute for Risk Assessment. Available online: https://www.animaltestinfo.de/act_search_file.cfm?CFID=35551967&CFTOKEN=501e9b7e37caad25-F6B7F831-B846-42FC-C0EA7A8FA04B37D3; (accessed on 22 May 2023).
- Davison, T.F. The immunologists’ debt to the chicken. Br. Poult. Sci. 2003, 44, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, G.R.; Marchant, T.A.; Blas, J.; German, T. Corticosterone in feathers is a longterm, integrated measure of avian stress physiology. Funct. Ecol. 2008, 22, 494–500. [Google Scholar] [CrossRef]
- Lagadic, H.; Faure, J.M.; Mills, A.D.; Williams, J.B. Effects of blood sampling on plasma concentrations of corticosterone and glucose in laying hens caged in groups. Br. Poult. Sci. 1990, 31, 823–829. [Google Scholar] [CrossRef]
- Knowles, T.G.; Broom, D.M. Effect of catching method on the concentration of plasma corticosterone in end-of-lay battery hens. Vet. Rec. 1993, 133, 527–528. [Google Scholar] [CrossRef]
- Marino, L. Thinking chickens: A review of cognition, emotion, and behavior in the domestic chicken. Anim. Cogn. 2017, 20, 127–147. [Google Scholar] [CrossRef]
- Ericsson, M.; Fallahsharoudi, A.; Bergquist, J.; Kushnir, M.M.; Jensen, P. Domestication effects on behavioural and hormonal responses to acute stress in chickens. Physiol. Behav. 2014, 133, 161–169. [Google Scholar] [CrossRef]
- Dereli Fidan, E.; Türkyılmaz, M.K.; Nazlıgül, A.; Ünübol Aypak, S.; Karaarslan, S. Effect of Preslaughter Shackling on Stress, Meat Quality Traits, and Glycolytic Potential in Broilers. J. Agric. Sci. Tech. 2018, 17, 1141–1150. [Google Scholar]
- Krause, E.T.; Ruploh, T. Captive domesticated zebra finches (Taeniopygia guttata) have increased plasma corticosterone concentrations in the absence of bathing water. Appl. Anim. Behav. Sci. 2016, 182, 80–85. [Google Scholar] [CrossRef]
- Honarmand, M.; Goymann, W.; Naguib, M. Stressful dieting: Nutritional conditions but not compensatory growth elevate corticosterone levels in zebra finch nestlings and fledglings. PLoS ONE 2010, 5, e12930. [Google Scholar] [CrossRef]
- Cockrem, J.F.; Silverin, B. Variation within and between birds in corticosterone responses of great tits (Parus major). Gen. Comp. Endocrinol. 2002, 125, 197–206. [Google Scholar] [CrossRef]
- Chloupek, P.; Bedanova, I.; Chloupek, J.; Vecerek, V. Changes in selected biochemical indices resulting from various pre-sampling handling techniques in broilers. Acta Vet. Scand. 2011, 53, 31. [Google Scholar] [CrossRef]
- Badmus, K.A.; Idrus, Z.; Meng, G.Y.; Sazili, A.Q.; Mamat-Hamidi, K. Telomere Length and Regulatory Genes as Novel Stress Biomarkers and their Diversities in Broiler Chickens (Gallus gallus domesticus) Subjected to Corticosterone Feeding. Animals 2021, 11, 2759. [Google Scholar] [CrossRef]
- Al-Maksousi, S.K.; Al-Hayani, W.K.; Hussein, F.M. Effect of the level of corticosteron hormone in the blood of local iraqi chicken mothers on the sex ratio of the produced offsprings. Plant Arch. 2019, 19, 1411–1415. [Google Scholar]
- Weimer, S.L.; Wideman, R.F.; Scanes, C.G.; Mauromoustakos, A.; Christensen, K.D.; Vizzier-Thaxton, Y. An evaluation of methods for measuring stress in broiler chickens. Poult. Sci. 2018, 97, 3381–3389. [Google Scholar] [CrossRef]
- Abobaker, H.; Hu, Y.; Hou, Z.; Sun, Q.; Idriss, A.A.; Omer, N.A.; Zong, Y.; Zhao, R. Dietary betaine supplementation increases adrenal expression of steroidogenic acute regulatory protein and yolk deposition of corticosterone in laying hens. Poult. Sci. 2017, 96, 4389–4398. [Google Scholar] [CrossRef]
- Antonissen, G.; De Baere, S.; Devreese, M.; Van Immerseel, F.; Martel, A.; Croubels, S. Feed contamination with Fusarium mycotoxins induces a corticosterone stress response in broiler chickens. Poult. Sci. 2017, 96, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Huth, J.C.; Archer, G.S. Comparison of two LED light bulbs to a dimmable CFL and their effects on broiler chicken growth, stress, and fear. Poult. Sci. 2015, 94, 2027–2036. [Google Scholar] [CrossRef]
- Mirfendereski, E.; Jahanian, R. Effects of dietary organic chromium and vitamin C supplementation on performance, immune responses, blood metabolites, and stress status of laying hens subjected to high stocking density. Poult. Sci. 2015, 94, 281–288. [Google Scholar] [CrossRef]
- Xie, J.; Tang, L.; Lu, L.; Zhang, L.; Lin, X.; Liu, H.C.; Odle, J.; Luo, X. Effects of acute and chronic heat stress on plasma metabolites, hormones and oxidant status in restrictedly fed broiler breeders. Poult. Sci. 2015, 94, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, H.A.; Purswell, J.L.; Collier, S.D.; Branton, S.L. Effects of genetic strain and light intensity on blood physiological variables of broilers grown to heavy weights. Poult. Sci. 2014, 93, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Vosmerova, P.; Chloupek, J.; Bedanova, I.; Chloupek, P.; Kruzikova, K.; Blahova, J.; Vecerek, V. Changes in selected biochemical indices related to transport of broilers to slaughterhouse under different ambient temperatures. Poult. Sci. 2010, 89, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yue, H.Y.; Zhang, H.J.; Xu, L.; Wu, S.G.; Yan, H.J.; Gong, Y.S.; Qi, G.H. Transport stress in broilers: I. Blood metabolism, glycolytic potential, and meat quality. Poult. Sci. 2009, 88, 2033–2041. [Google Scholar] [CrossRef]
- Post, J.; Rebel, J.M.; Ter Huurne, A.A. Physiological effects of elevated plasma corticosterone concentrations in broiler chickens. An alternative means by which to assess the physiological effects of stress. Poult. Sci. 2003, 82, 1313–1318. [Google Scholar] [CrossRef]
- Samir, H.; Nyametease, P.; Elbadawy, M.; Fathi, M.; Mandour, A.S.; Radwan, F.; Nagaoka, K.; Sasaki, K.; Watanabe, G. Assessment of correlations and concentrations of salivary and plasma steroids, testicular morphometry, and semen quality in different climatic conditions in goats. Theriogenology 2020, 157, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Vincent, I.C.; Michell, A.R. Comparison of cortisol concentrations in saliva and plasma of dogs. Res. Vet. Sci. 1992, 53, 342–345. [Google Scholar] [CrossRef]
- Teruhisa, U.; Ryoji, H.; Taisuke, I.; Tatsuya, S.; Fumihiro, M.; Tatsuo, S. Use of saliva for monitoring unbound free cortisol levels in serum. Clin. Chim. Acta 1981, 110, 245–253. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Shutt, D.A. Salivary and plasma cortisol as an index of stress in goats. Aust. Vet. J. 1992, 69, 161–163. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Hellhammer, D.H. Salivary Cortisol. In Encyclopedia of Stress; Fink, G., Ed.; Academic Press: San Diego, CA, USA, 2000; pp. 379–383. ISBN 978-0122267383. [Google Scholar]
- Rød, A.M.K.; Harkestad, N.; Jellestad, F.K.; Murison, R. Comparison of commercial ELISA assays for quantification of corticosterone in serum. Sci. Rep. 2017, 7, 6748. [Google Scholar] [CrossRef]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Stanford, S.C. The open field test: Reinventing the wheel. J. Psychopharmacol. 2007, 21, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.S. Emotional behavior in the rat. I. Defecation and urination as measures of individual differences in emotionality. J. Com. Psychol. 1934, 18, 385. [Google Scholar] [CrossRef]
- Galli, S.J.; Borregaard, N.; Wynn, T.A. Phenotypic and functional plasticity of cells of innate immunity: Macrophages, mast cells and neutrophils. Nat. Immunol. 2011, 12, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Li, D.; Li, Z.; Dong, B.; Wang, F. Effects of glycinin on IgE-mediated increase of mast cell numbers and histamine release in the small intestine. J. Nutr. Biochem. 2008, 19, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Metz, M.; Piliponsky, A.M.; Chen, C.C.; Lammel, V.; Åbrink, M.; Pejler, G.; Tsai, M.; Galli, S.J. Mast cells can enhance resistance to snake and honeybee venoms. Science 2006, 313, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Tschaikowski, K.L.; Jorde, W. Allergische Krankheiten des Magen-Darm-Traktes: Ein Ratgeber für die Praxis; Springer-Verlag: Berlin/Heidelberg, Germany, 1989; ISBN 9783540513629. [Google Scholar]
- Gershon, M. Der kluge Bauch: Die Entdeckung des zweiten Gehirns; Goldmann: Munich, Germany, 2001; ISBN 978-3442151141. [Google Scholar]
- Moeser, A.J.; Ryan, K.A.; Nighot, P.K.; Blikslager, A.T. Gastrointestinal dysfunction induced by early weaning is attenuated by delayed weaning and mast cell blockade in pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Pees, M.; Pees, K. Papageien und Sittiche–Die Untersuchung der Faeces als wichtige diagnostische Möglichkeit. Kleintier Konkret 2008, 11, 18–23. [Google Scholar] [CrossRef]
- Filaretova, L.; Bagaeva, T. The Realization of the Brain-Gut Interactions with Corticotropin-Releasing Factor and Glucocorticoids. Curr. Neuropharmacol. 2016, 14, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Fraisse, F.; Cockrem, J.F. Corticosterone and fear behaviour in white and brown caged laying hens. Br. Poult. Sci. 2006, 47, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, M.; Pschernig, E.; Wallner, B.; Millesi, E. Non-invasive cortisol measurements as indicators of physiological stress responses in guinea pigs. PeerJ 2016, 4, 1590. [Google Scholar] [CrossRef] [PubMed]
- Freeman, N.E.; Newman, A.E. Quantifying corticosterone in feathers: Validations for an emerging technique. Conserv. Physiol. 2018, 6, 51. [Google Scholar] [CrossRef]
- Kobayashi, H.; Miyazaki, Y. Distribution characteristics of salivary cortisol measurements in a healthy young male population. J. Physiol. Anthr. 2015, 34, 30. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Hillsdale, N.Y., Ed.; Lawrence Earlbaum Associates: Mahwah, NJ, USA, 1988; ISBN 0-8058-0283-5. [Google Scholar]
- Carabajal, A. (Autonomous University of Barcelona, Barcelona, Spain). Personal communication. 11 November 2020. [Google Scholar]
- Hasan Siddiqui, S.; Kang, D.; Park, J.; Choi, H.W.; Shim, K. Acute heat stress induces the differential expression of heat shock proteins in different sections of the small intestine of chickens based on exposure duration. Animals 2020, 10, 1234. [Google Scholar] [CrossRef]
- Qureshi, S.; Khan, H.M.; Shafi, S.; Adil, S.; Mir, M.S. Effect of cold stress and its various ameliorating remedies on histomorphology of lymphoid glands (Bursa of Fabricius and Spleen) in broiler chicken. J. Entomol. Zool. Stud. 2020, 8, 866–871. [Google Scholar]
- Hartanto, S.; Ko, H.S.; Jee, S.H.; Kang, J.U.; Seo, J.S.; Kang, Y.H.; Kim, H.N.; Ohh, S.J. Effect of dietary nutmeg oil on heat stress tolerance related parameters in Korean native chicken reared under hot temperature. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1160–1167. [Google Scholar] [CrossRef]
- Zaboli, G.; Huang, X.; Feng, X.; Ahn, D.U. How can heat stress affect chicken meat quality?—A review. Poult. Sci. 2019, 98, 1551–1556. [Google Scholar] [CrossRef]
- Qureshi, S.; Khan, H.M.; Mir, M.S.; Raja, T.A.; Khan, A.A.; Ali, H.; Adil, S. Effect of cold stress and various suitable remedies on performance of broiler chicken. J. World Poult. Res. 2018, 8, 66–73. [Google Scholar]
- Nissa, S.S.; Sheikh, I.U.; Banday, M.T.; Zaffer, B.; Shafiq, S.; Chishti, A.; Qureshi, A.I. Estimation of stress level of broiler chicken reared under different sources of light. J. Entomol. Zool. Stud. 2018, 6, 363–365. [Google Scholar]
- Li, W.; Wei, F.; Xu, B.; Sun, Q.; Deng, W.; Ma, H.; Bai, J.; Li, S. Effect of stocking density and alpha-lipoic acid on the growth performance, physiological and oxidative stress and immune response of broilers. Asian-Australas. J. Anim. Sci. 2019, 32, 1914–1922. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, X.; Wang, W.; Zhang, H.; Xu, S. Application of transcriptome analysis: Oxidative stress, inflammation and microtubule activity disorder caused by ammonia exposure may be the primary factors of intestinal microvilli deficiency in chicken. Sci. Total Environ. 2019, 696, 134035. [Google Scholar] [CrossRef]
- Park, B.S.; Um, K.H.; Park, S.O.; Zammit, V.A. Effect of stocking density on behavioral traits, blood biochemical parameters and immune responses in meat ducks exposed to heat stress. Arch. Anim. Breed. 2018, 61, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Park, S.B.; Kim, S.H.; Kim, C.H. Effects of stock density on the laying performance, blood parameter, corticosterone, litter quality, gas emission and bone mineral density of laying hens in floor pens. Poult. Sci. 2016, 95, 2764–2770. [Google Scholar] [CrossRef]
- Yan, C.; Xiao, J.; Chen, D.; Turner, S.P.; Li, Z.; Liu, H.; Liu, W.; Liu, J.; Chen, S.; Zhao, X. Feed Restriction Induced Changes in Behavior, Corticosterone, and Microbial Programming in Slow-and Fast-Growing Chicken Breeds. Animals 2021, 11, 141. [Google Scholar] [CrossRef]
- Bowling, M.; Forder, R.; Hughes, R.J.; Weaver, S.; Hynd, P.I. Effect of restricted feed intake in broiler breeder hens on their stress levels and the growth and immunology of their offspring. Transl. Anim. Sci. 2018, 2, 263–271. [Google Scholar] [CrossRef]
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Goh, Y.M. Acute phase proteins response to feed deprivation in broiler chickens. Poult. Sci. 2016, 95, 760–763. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Lensen, C.M.; Moons, C.P.; Diederich, C. Saliva sampling in dogs: How to select the most appropriate procedure for your study. J. Vet. Behav. 2015, 10, 504–512. [Google Scholar] [CrossRef]
- Hernandez, C.E.; Thierfelder, T.; Svennersten-Sjaunja, K.; Berg, C.; Orihuela, A.; Lidfors, L. Time lag between peak concentrations of plasma and salivary cortisol following a stressful procedure in dairy cattle. Acta Vet. Scand. 2014, 56, 61. [Google Scholar] [CrossRef]
- Chacón, G.; Laita, S.G.B.; del Portal, J.C.I.; Liesa, J.P. Validation of an EIA technique for the determination of salivary cortisol in cattle. Span. J. Agric. Res. 2004, 2, 45–52. [Google Scholar] [CrossRef]
- Kobelt, A.J.; Hemsworth, P.H.; Barnett, J.L.; Butler, K.L. Sources of sampling variation in saliva cortisol in dogs. Res. Vet. Sci. 2003, 75, 157–161. [Google Scholar] [CrossRef]
- Korbel, R.; Lierz, M. Klinische Labordiagnostik beim Vogel. Spezielle Untersuchungen und Labordiagnostik ausgewählter Tierspezies. In Klinische Labordiagnostik in der Tiermedizin, 7th ed.; Kraft, W., Dürr, U.M., Möritz, A., Eds.; Schattauer: Stuttgart, Germany, 2014; ISBN 9783794527373. [Google Scholar]
- Schmidt, K. Würde oder Integrität–Verlangt die gentechnische Veränderung von Tieren neue tierethische Konzepte. Altex 2008, 25, 313–320. [Google Scholar] [CrossRef]
- Pastore, C.; Pirrone, F.; Balzarotti, F.; Faustini, M.; Pierantoni, L.; Albertini, M. Evaluation of physiological and behavioral stress-dependent parameters in agility dogs. J. Vet. Behav. 2011, 6, 188–194. [Google Scholar] [CrossRef]
- Fölsch, D.W.; Vestergaard, K. Das Verhalten von Hühnern. / The behavior of Fowl. In Das Normalverhalten und die Auswirkung Verschiedener Haltungssysteme und Aufzuchtmethoden, 12th ed.; Birkhäuser, Ed.; Springer: Basel, Switzerland, 1981; ISBN 978-3-7643-1240-4. [Google Scholar]
- Calder, C.; Albright, J. Chicken behavior. Backyard Poultry Medicine and Surgery: A Guide for Veterinary Practitioners, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2021; pp. 434–454. ISBN 978-1-119-51175-5. [Google Scholar]
- Lages, Y.V.; Maisonnette, S.S.; Marinho, B.; Rosseti, F.P.; Krahe, T.E.; Landeira-Fernandez, J. Behavioral effects of chronic stress in Carioca high-and low-conditioned freezing rats. Stress 2021, 24, 602–611. [Google Scholar] [CrossRef]
- Campbell, D.L.; Taylor, P.S.; Hernandez, C.E.; Stewart, M.; Belson, S.; Lee, C. An attention bias test to assess anxiety states in laying hens. PeerJ 2019, 7, 7303. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.H. Open-field freezing as a stable parameter of wildness in Mus musculus. Behav. Biol. 1978, 23, 67–74. [Google Scholar] [CrossRef]
- Jones, R.B.; Blokhuis, H.J.; Beuving, G. Open field and tonic immobility responses in domestic chicks of two genetic lines differing in their propensity to feather peck. Br. Poult. Sci. 1995, 36, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.D.; Gallup, G.G., Jr. Open-field behaviour in chickens: A replication revisited. Behav Process. 1985, 10, 333–340. [Google Scholar] [CrossRef]
- Suarez, S.D.; Gallup, G.G. Predatory overtones of open-field testing in chickens. Anim. Learn. Behav. 1981, 9, 153–163. [Google Scholar] [CrossRef]
- Suarez, S.D.; Gallup, G.G. Social reinstatement and open-field testing in chickens. Anim. Learn. Behav. 1983, 11, 119–126. [Google Scholar] [CrossRef]
- McKeegan, D.E.; Smith, F.S.; Demmers, T.G.; Wathes, C.M.; Jones, R.B. Behavioral correlates of olfactory and trigeminal gaseous stimulation in chickens, Gallus domesticus. Physiol. Behav. 2005, 84, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Burne, T.H.J.; Rogers, L.J. Odors, volatiles and approach-avoidance behavior of the domestic chick (Gallus gallus domesticus). Int. J. Com. Psychol. 1995, 8, 99–114. [Google Scholar] [CrossRef]
- Vallortigara, G.; Cailotto, M.; Zanforlin, M. Sex differences in social reinstatement motivation of the domestic chick (Gallus gallus) revealed by runway tests with social and nonsocial reinforcement. J. Comp. Psychol. 1990, 104, 361. [Google Scholar] [CrossRef] [PubMed]
- Gallup, G.G., Jr.; Suarez, S.D. An ethological analysis of open-field behaviour in chickens. Anim. Behav. 1980, 28, 368–378. [Google Scholar] [CrossRef]
- Gallup, G.G., Jr.; Rager, D.R.; Scheuerman, K.W. Predatory aspects of open field testing: Towards a functional analysis of ongoing behaviour in chickens. Bird Behav. 1990, 8, 67–72. [Google Scholar] [CrossRef]
- Sethy, K.; Dash, R.; Vaidantika, V.; Priyadarshinee, P.; Dhaigude, V.; Dwibedy, P.; Adhikary, P.D. Cannibalism: A threat to poultry industry. INDiAN FARmER 2018, 5, 357–370. [Google Scholar]
- Gvaryahu, G.; Ararat, E.; Asaf, E.; Lev, M.; Weller, J.I.; Robinzon, B.; Snapir, N. An enrichment object that reduces aggressiveness and mortality in caged laying hens. Physiol. Behav. 1994, 55, 313–316. [Google Scholar] [CrossRef]


{kind=link}
| Behavior Exhibited | Behavior Is Equal |
|---|---|
| Axial body shaking | → to an axial shaking movement out and back to the starting position |
| Locomotion | → to moving of the chicken by one body length |
| Fecal Parameters | Type of Assessment |
|---|---|
| Number of fecal droppings | Ordinal: 0–3 |
| Fecal consistency | Nominal: 1 = well formed, with clearly separated uric acid and fecal components; 2 = pasty, no clear separation of components; 3 = mushy, clearly increased to separate liquid part, pungent odor |
| Plasma Corticosterone | ||||||||
|---|---|---|---|---|---|---|---|---|
| Sample | Number of Individuals (n) | Geometric Mean (ng/mL) | t-Test for Independent Samples | |||||
| C | T | C | T | df | t-Value | p-Value | Cohen’s d | |
| 1 | 21 | 21 | 1.998 | 2.948 | 40 | 2.106 | 0.042 | 0.650 |
| 2 | 21 | 21 | 1.832 | 1.524 | 40 | −1.057 | 0.297 | −0.326 |
| 3 | 21 | 21 | 2.581 | 2.450 | 40 | −0.323 | 0.748 | −0.100 |
| Saliva Corticosterone | ||||||||
|---|---|---|---|---|---|---|---|---|
| Sample | Number of Individuals (n) | Median (ng/mL) | Mann–Whitney U Test | |||||
| C | T | C | T | U | Z | p-Value | Effect Size | |
| 1 | 20 | 20 | 0.713 | 1.218 | 167.5 | −0.879 | 0.379 | 0.139 |
| 2 | 21 | 20 | 1.316 | 0.825 | 123.0 | −2.269 | 0.023 | 0.354 |
| 3 | 21 | 20 | 1.038 | 0.915 | 169.0 | −1.069 | 0.285 | 0.167 |
| Axial Body Shaking | |||||||
| Number of Individuals * (n) | Median | Mann–Whitney U Test | |||||
| C | T | C | T | U | Z | p-Value | Effect Size |
| 20 | 17 | 2.5 | 1.5 | 91.5 | −2.413 | 0.016 | 0.400 |
| Locomotion | |||||||
| Number of Individuals * (n) | Mean | t-Test for Independent Samples | |||||
| C | T | C | T | df | t-Value | p-Value | Cohen’s d |
| 20 | 17 | 20.05 | 18.18 | 35 | −0.938 | 0.345 | −0.310 |
| Defecation in the New Area Test | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Number of Individuals (n) | Median | Mann–Whitney U Test | |||||
| C | T | C | T | U | Z | p-Value | Effect Size | |
| Number of droppings deposited (0–3) | 21 | 21 | 2 | 2 | 170.5 | −1.378 | 0.168 | 0.213 |
| Consistency of droppings (Scoring: 1–3) | 21 | 20 | 1.67 | 1 | 54.0 | −4.584 | 0.000 | 0.716 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mählis, G.; Kleine, A.; Lüschow, D.; Bartel, A.; Wiegard, M.; Thoene-Reineke, C. Clicker Training as an Applied Refinement Measure in Chickens. Animals 2023, 13, 3836. https://doi.org/10.3390/ani13243836
Mählis G, Kleine A, Lüschow D, Bartel A, Wiegard M, Thoene-Reineke C. Clicker Training as an Applied Refinement Measure in Chickens. Animals. 2023; 13(24):3836. https://doi.org/10.3390/ani13243836
Chicago/Turabian StyleMählis, Gordon, Anne Kleine, Dörte Lüschow, Alexander Bartel, Mechthild Wiegard, and Christa Thoene-Reineke. 2023. "Clicker Training as an Applied Refinement Measure in Chickens" Animals 13, no. 24: 3836. https://doi.org/10.3390/ani13243836
APA StyleMählis, G., Kleine, A., Lüschow, D., Bartel, A., Wiegard, M., & Thoene-Reineke, C. (2023). Clicker Training as an Applied Refinement Measure in Chickens. Animals, 13(24), 3836. https://doi.org/10.3390/ani13243836