

Mechanisms of Digestive Enzyme Response to Acute Salinity Stress in Juvenile Yellowfin Tuna (Thunnus albacares)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
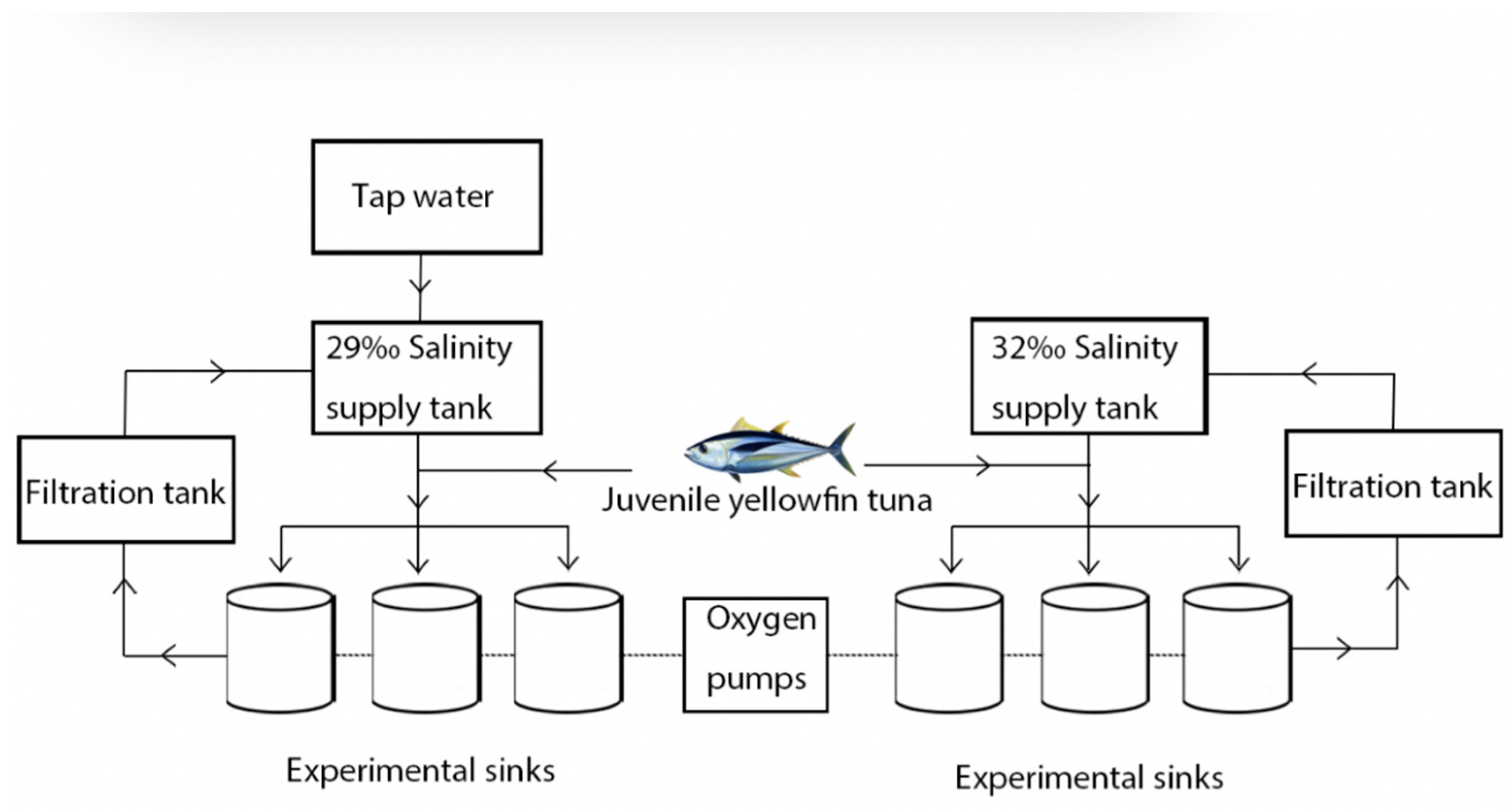
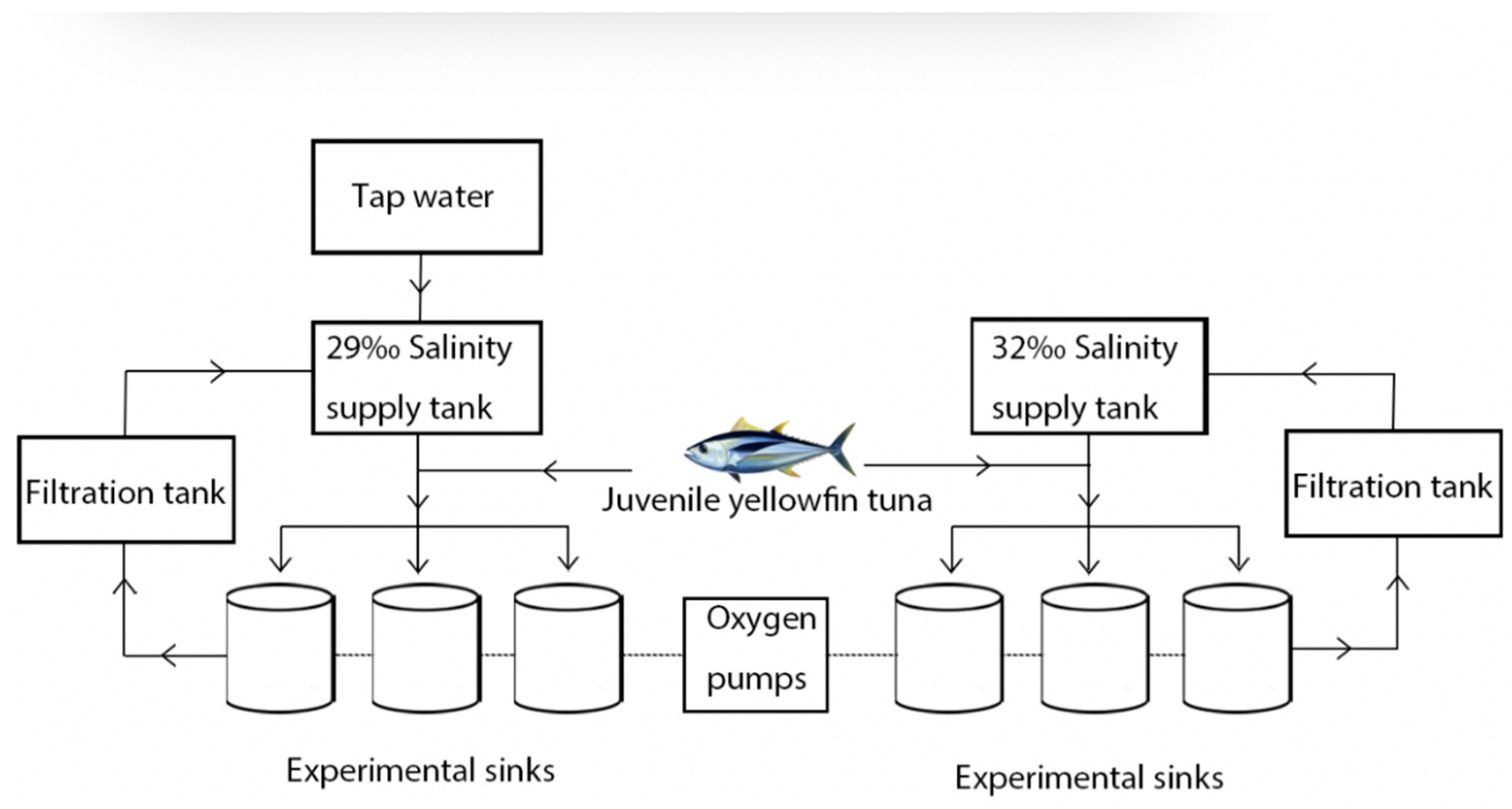
2.1. Experimental Methods and Design
2.2. Analytical Method
2.3. Statistical Analysis
3. Results
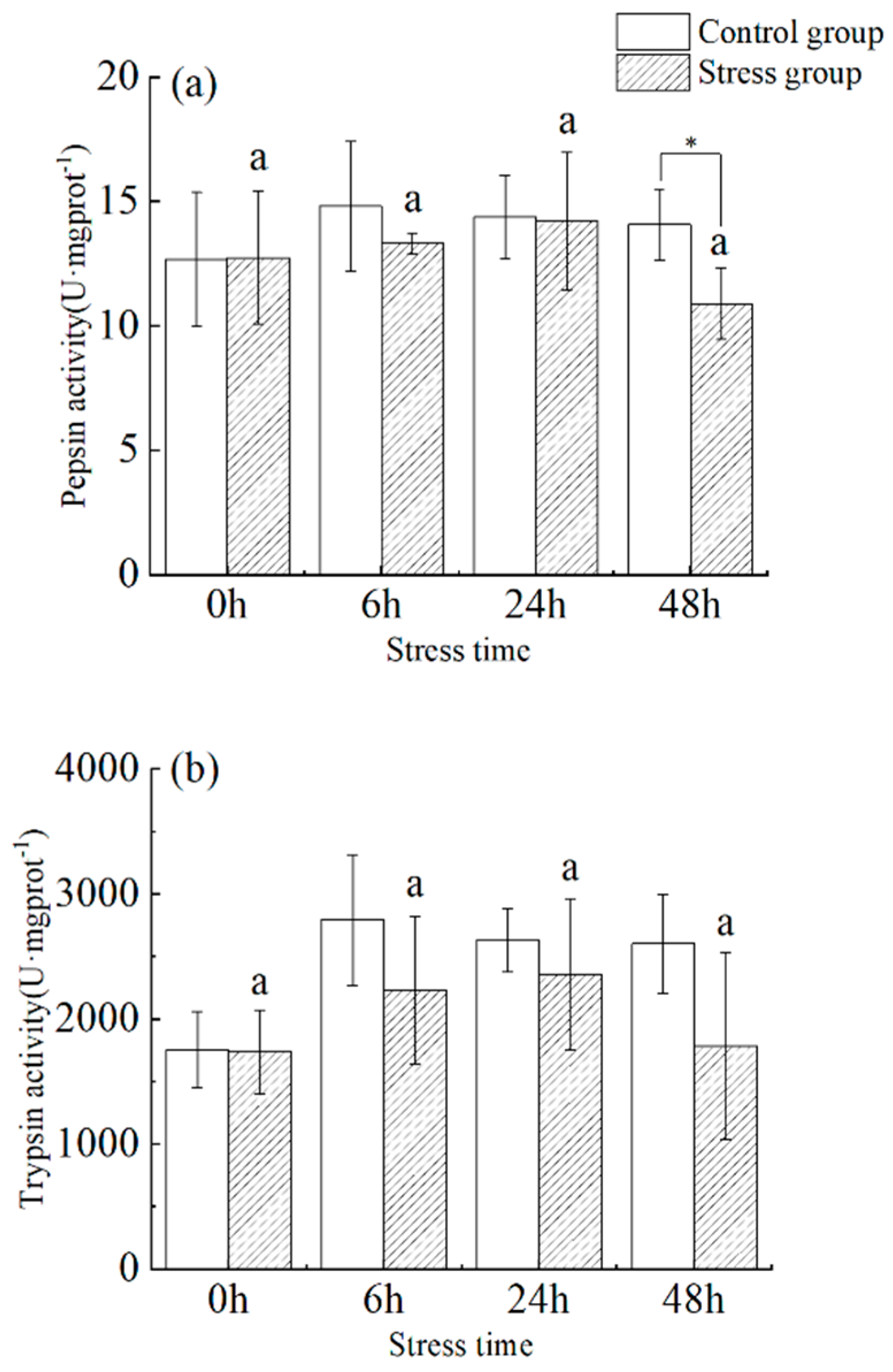
3.1. Effect of Salinity Changes on Pepsin and Trypsin Activity
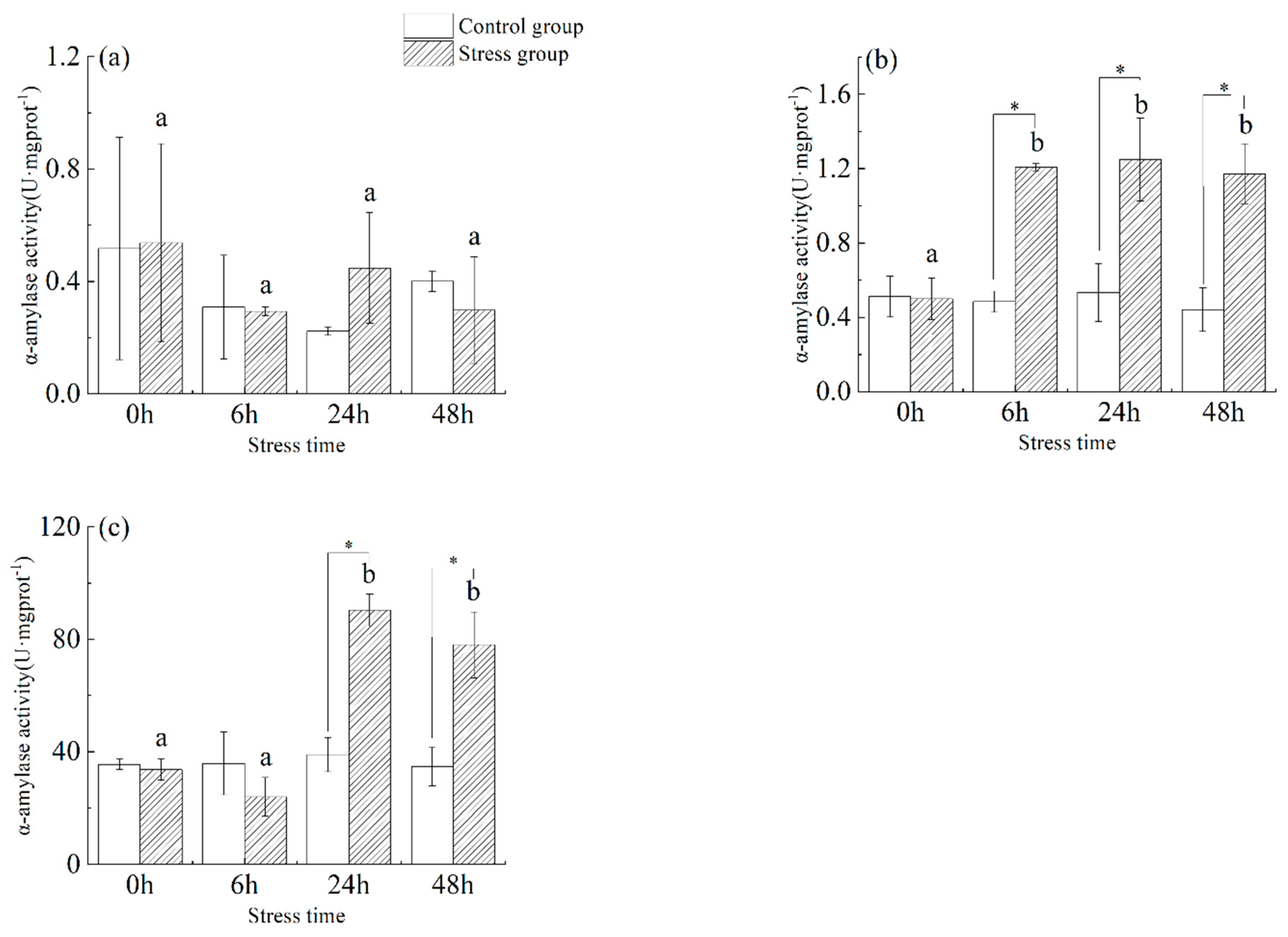
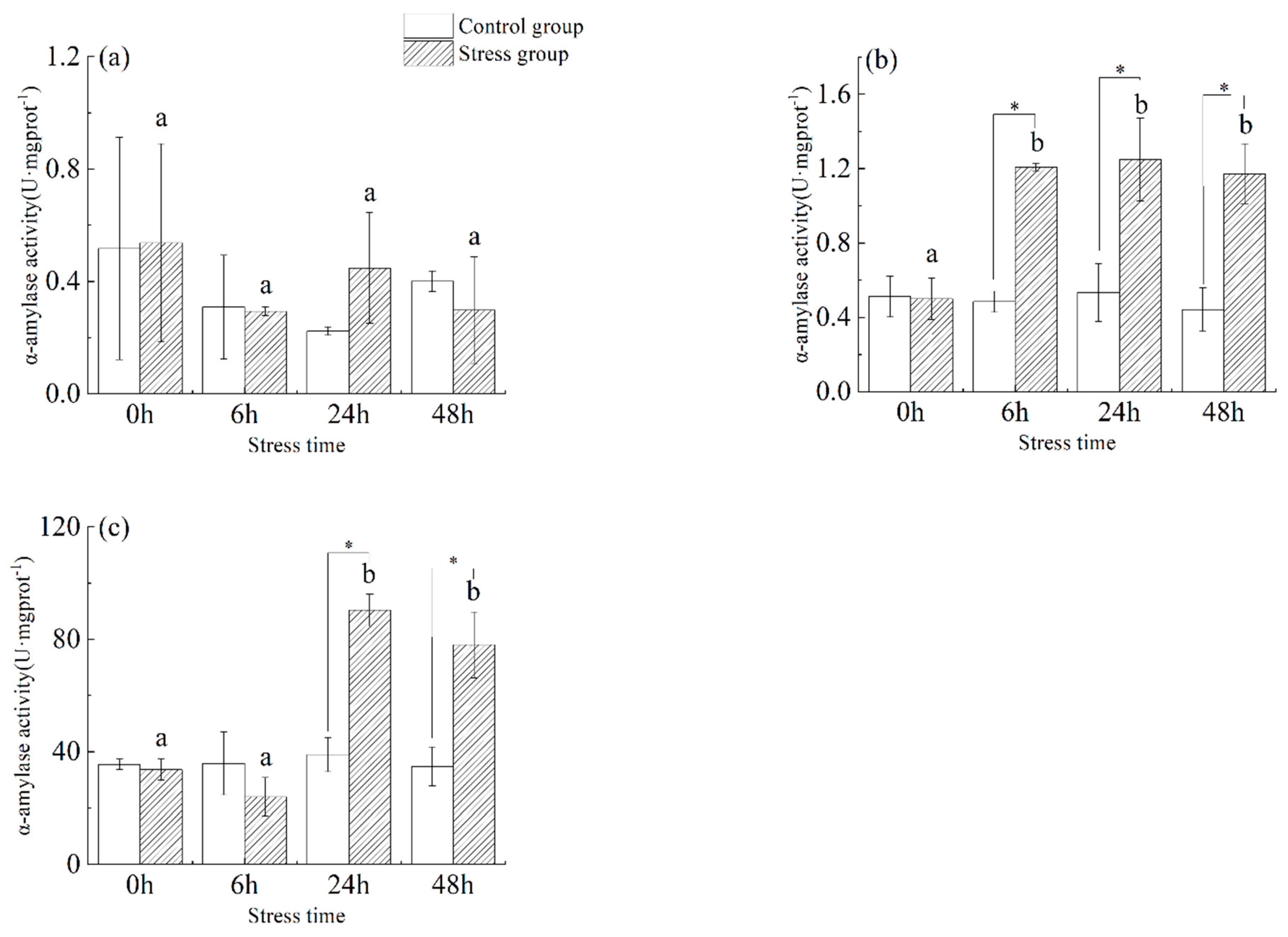
3.2. Effect of Salinity Changes on α-Amylase Activity
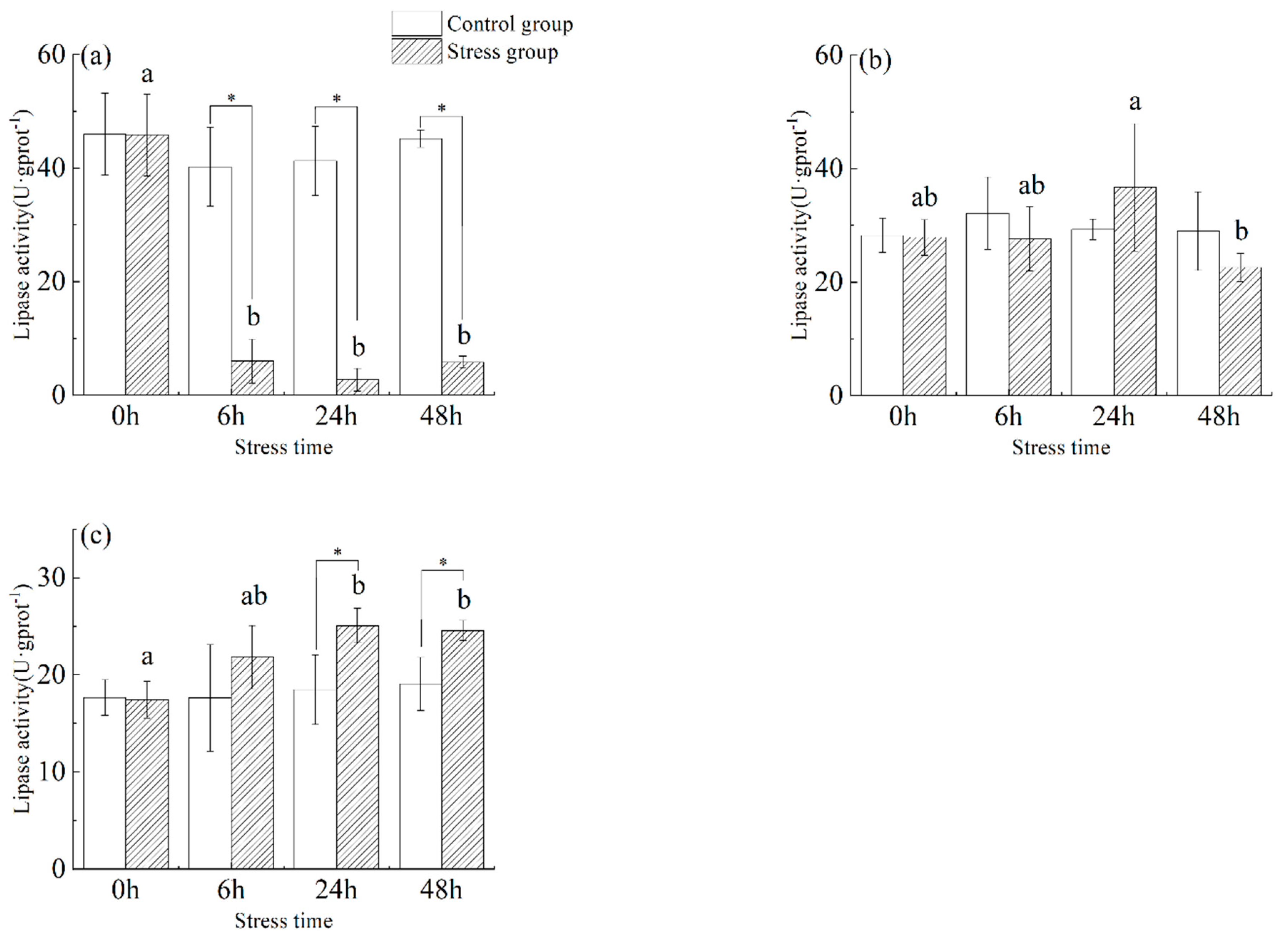
3.3. Effect of Salinity Changes on Lipase Activity
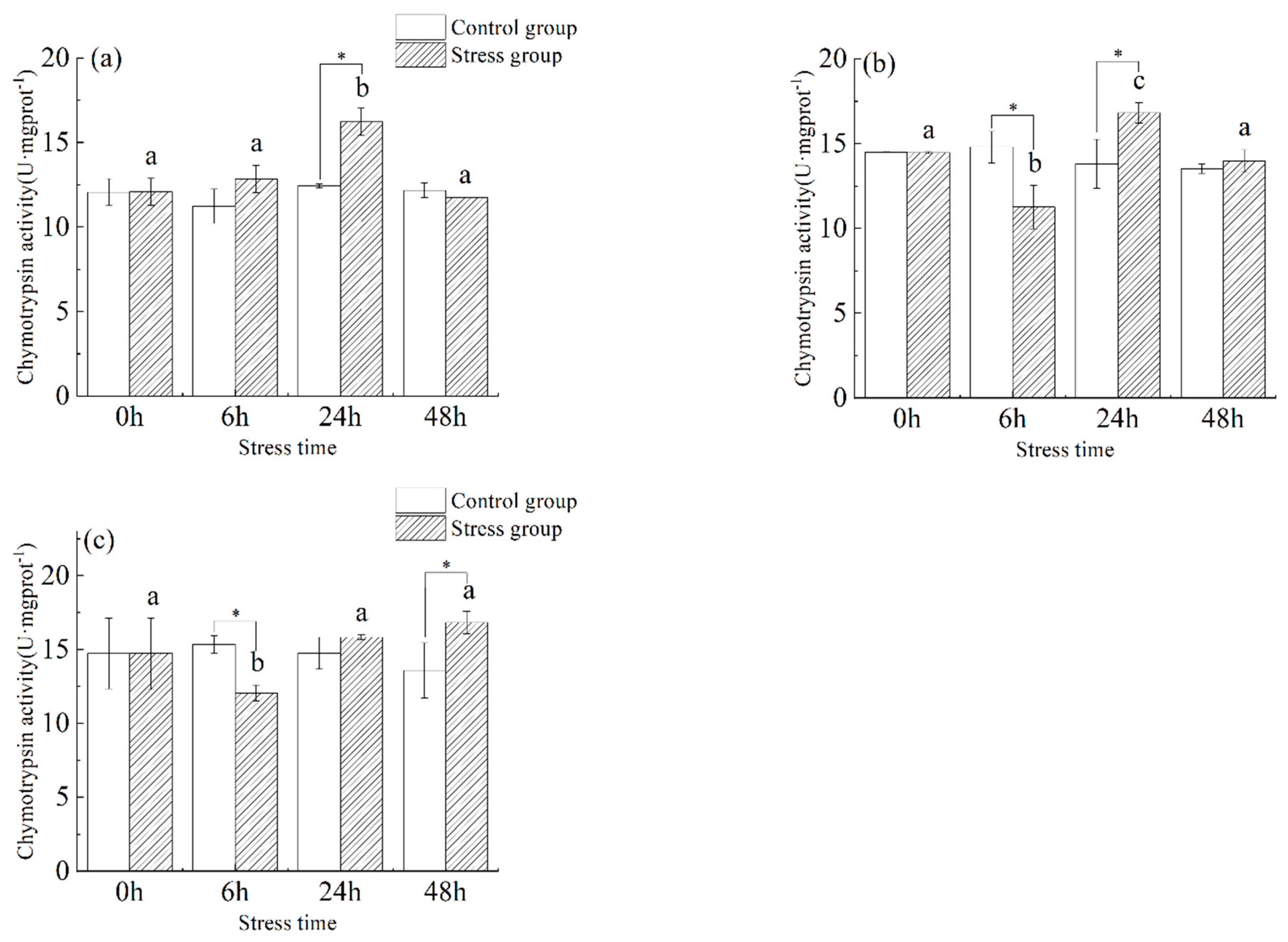
3.4. Effect of Salinity Changes on Chymotrypsin Activity
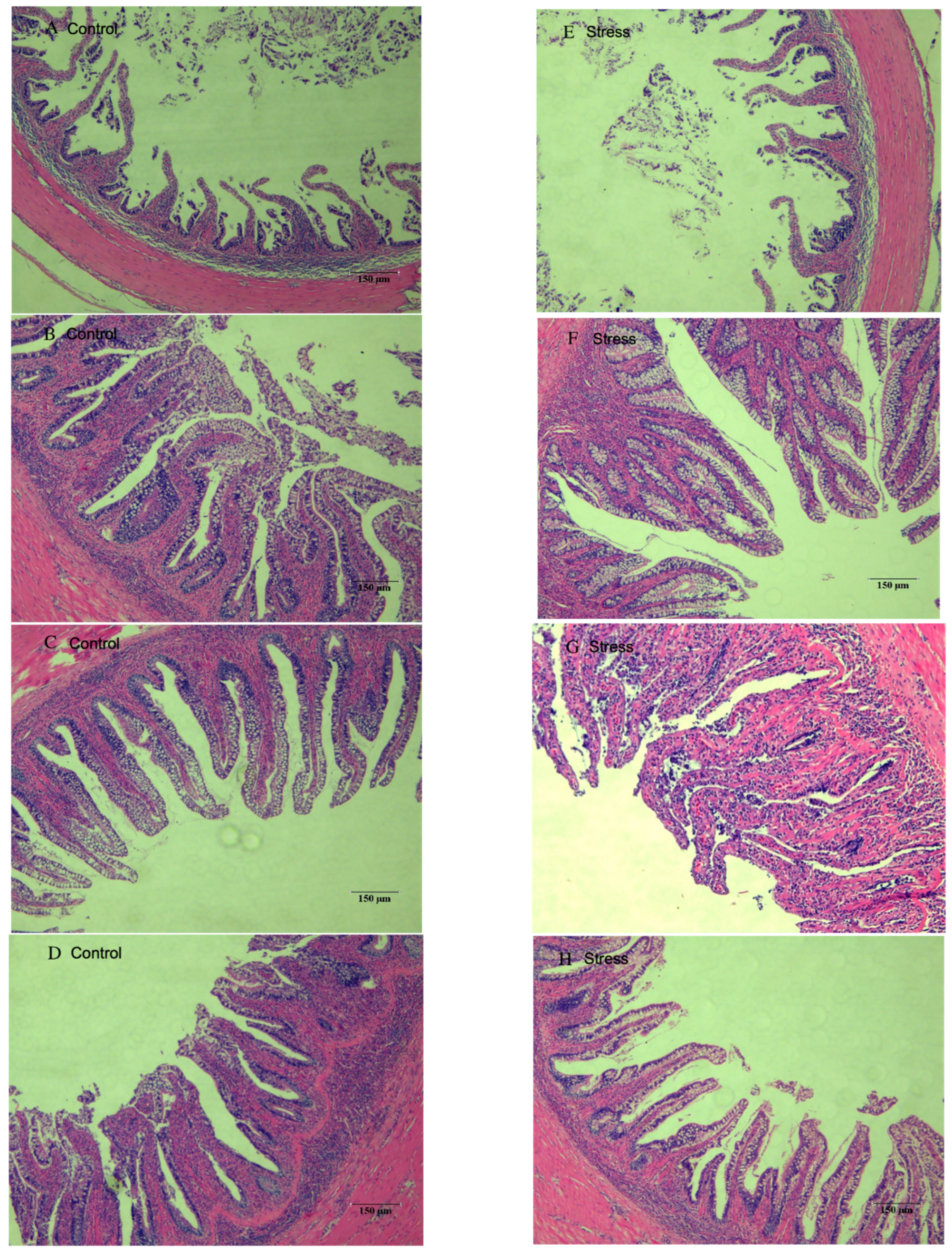
3.5. Effect of Salinity Changes on Foregut Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Surhone, L.M.; Tennoe, M.T.; Henssonow, S.F. Yellowfin Tuna; Betascript Publishing: New York, NY, USA, 2010. [Google Scholar]
- Sun, R.-X.; Sun, Y.; Xie, X.-D.; Yang, B.-Z.; Cao, L.-Y.; Luo, S.; Wang, Y.-Y.; Mai, B.-X. Bioaccumulation and human health risk assessment of DDT and its metabolites (DDTs) in yellowfin tuna (Thunnus albacares) and their prey from the South China Sea. Mar. Pollut. Bull. 2020, 158, 111396. [Google Scholar] [CrossRef]
- FAO (Fisheries and Aquaculture Department). The State of World Fisheries and Aquaculture; FAO Fisheries Report; FAO: Rome, Italy, 2020; Volume 2020, pp. 6–8. [Google Scholar]
- Murua, H.; Rodriguez-Marin, E.; Neilson, J.D.; Farley, J.H.; Juan-Jordá, M.J. Fast versus slow growing tuna species: Age, growth, and implications for population dynamics and fisheries management. Rev. Fish Biol. Fish. 2017, 27, 733–773. [Google Scholar] [CrossRef]
- Tian, Z.P.; Wang, F.; Tian, S.Q.; Ma, Q.Y. Stock assessment for Atlantic yellowfin tuna based on extended surplus production model considering life history. Acta Oceanol. Sin. 2022, 41, 41–51. [Google Scholar] [CrossRef]
- Zhou, W.F.; Hu, H.J.; Wei, F.; Jin, S.F. Impact of Abnormal Climatic Events on the CPUE of Yellowfin Tuna Fishing in the Central and Western Pacific. Sustainability 2022, 14, 1217. [Google Scholar] [CrossRef]
- Pacicco, A.E.; Brown-Peterson, N.J.; Murie, D.J.; Allman, R.J.; Snodgrass, D.; Franks, J.S. Reproductive biology of yellowfin tuna (Thunnus albacares) in the northcentral U.S. Gulf of Mexico. Fish. Res. 2023, 261, 106620. [Google Scholar] [CrossRef]
- Schaefer, K.M.; Fuller, D.W. Spatiotemporal variability in the reproductive biology of yellowfin tuna (Thunnus albacares) in the eastern Pacific Ocean. Fish. Res. 2022, 248, 106225. [Google Scholar] [CrossRef]
- Hoyle, S.D.; Williams, A.J.; Minte-Vera, C.V.; Maunder, M.N. Approaches for estimating natural mortality in tuna stock assessments: Application to global yellowfin tuna stocks. Fish. Res. 2023, 257, 106498. [Google Scholar] [CrossRef]
- Filous, A.; Friedlander, A.M.; Griffin, L.; Lennox, R.J.; Danylchuk, A.J.; Mereb, G.; Golbuu, Y. Movements of juvenile yellowfin tuna (Thunnus albacares) within the coastal FAD network adjacent to the Palau National Marine Sanctuary: Implications for local fisheries development. Fish. Res. 2020, 230, 105688. [Google Scholar] [CrossRef]
- Sánchez-Parra, M.; Lopez, A.; Muñoz-Redondo, J.M.; Montenegro-Gómez, J.C.; Pérez-Aparicio, J.; Pereira-Caro, G.; Rodríguez-Solana, R.; Moreno-Rojas, J.M.; Ordóñez-Díaz, J.L. Study of the influence of the fishing season and the storage temperature in the fishing vessel on the biogenic amine and volatile profiles in fresh yellowfin tuna (Thunnus albacares) and dry-cured mojama. J. Food Compos. Anal. 2022, 114, 104845. [Google Scholar] [CrossRef]
- Alexi, N.; Hvam, J.; Lund, B.W.; Nsubuga, L.; Hansen, R.M.d.O.; Thamsborg, K.; Lofink, F.; Byrne, D.V.; Leisner, J.J. Potential of novel cadaverine biosensor technology to predict shelf life of chilled yellowfin tuna (Thunnus albacares). Food Control. 2020, 119, 107458. [Google Scholar] [CrossRef]
- Huang, C.-H.; Hsieh, C.-Y.; Lee, Y.-C.; Ou, T.-Y.; Chang, T.-H.; Lee, S.-H.; Tseng, C.-H.; Tsai, Y.-H. Inhibitory Effects of High-Hydrostatic-Pressure Processing on Growth and Histamine Formation of Histamine-Forming Bacteria in Yellowfin Tuna Meat during Storage. Biology 2022, 11, 702. [Google Scholar] [CrossRef] [PubMed]
- Rahmah, A.; Mardhatillah, I.; Damora, A.; Muhammad, M.; Nurfadillah, N. Application of Surplus Production Model to the Yellowfin Tuna Thunnus albacares in the northern and western parts of Aceh waters. IOP Conf. Ser. Earth Environ. Sci. 2021, 869, 012072. [Google Scholar] [CrossRef]
- Nguyen, K.Q.; Phan, H.T.; Tran, P.D.; Van Nguyen, B.; Van Do, T.; Nguyen, L.T.; Van To, P.; Vu, N.K. Length-length, Length–weight, and Weight-weight Relationships of Yellowfin (Thunnus albacares) and Bigeye (Thunnus obesus) Tuna Collected From the Commercial Handlines Fisheries in the South China Sea. Int. J. Mar. Sci. 2022, 38, 911–917. [Google Scholar] [CrossRef]
- Liu, H.Y.; Fu, Z.Y.; Zhou, S.J.; Hu, J.; Yang, R.; Yu, G.; Ma, Z.H. The Complete Mitochondrial Genome of Pennella sp. Parasitizing Thunnus albacares. Front. Cells Infect. Microbiol. 2022, 12, 945152. [Google Scholar] [CrossRef]
- Sloman, K.A.; Wilson, R.W.; Balshine, S. Fish physiology. In Behaviour and Physiology of Fish; Elsevier: Amsterdam, The Netherlands, 2005; Volume 24, pp. 1–480. [Google Scholar]
- You, H.Z.; Sun, Z.J.; Zhang, Q.; Zhang, Z.K. Effects of salinity on feeding growth and body composition of juvenile leopard gill perch. J. Dalian Ocean. Univ. 2013, 28, 89–93. [Google Scholar]
- Liu, W.; Zhi, B.-J.; Zhan, P.-R.; Guan, H.-H.; Qin, D.-L. Effects of salinity on haematological biochemistrical indices and liver tissue in juvenile Oncorhynchus keta. J. Appl. Ecol. 2010, 21, 2411–2417. [Google Scholar]
- Jiang, Z.; Huang, X.; Zhang, J. Dynamics of nonstructural carbohydrates in seagrass Thalassia hemprichii and its response to shading. Acta Oceanol. Sin. 2013, 32, 61–67. [Google Scholar] [CrossRef]
- Artetxe-Arratea, I.; Fraile, I.; Marsac, F.; Farley, J.H.; Murua, H. A review of the fisheries, life history and stock structure of tropical tuna (skipjack Katsuwonus pelamis, yellowfin Thunnus albacares and bigeye Thunnus obesus) in the Indian Ocean. Adv. Mar. Biol. 2020, 88, 39–89. [Google Scholar]
- Ashouri, G.; Soofiani, N.M.; Hoseinifar, S.H.; Jalali, S.A.H.; Morshedi, V.; Valinassab, T.; Bagheri, D.; Van Doan, H.; Mozanzadeh, M.T.; Carnevali, O. Influence of dietary sodium alginate and Pediococcus acidilactici on liver antioxidant status, intestinal lysozyme gene expression, histomorphology, microbiota, and digestive enzymes activity, in Asian sea bass (Lates calcarifer) juveniles. Aquaculture 2020, 518, 734638. [Google Scholar] [CrossRef]
- Saleh, N.E.; Helal, M.; Ali, N.G.; Abbas, E.; Abdel-Tawwab, M. Effects of using vital wheat gluten in practical diets on growth, intestinal histopathology, proinflammation-related gene expression, and resistance of white seabream (Diplodus sargus) to Staphylococcus epidermidis infection. Aquaculture 2021, 537, 736508. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Hayasaka, O.; Matsui, H.; Waqalevu, V.; Honda, A.; Nakajima, K.; Yamashita, H.; Ishikawa, M.; Shiozaki, K.; Kotani, T. Changes in early digestive tract morphology, enzyme expression and activity of Kawakawa tuna (Euthynnus affinis). Aquaculture 2021, 530, 735935. [Google Scholar] [CrossRef]
- Fruton, J.S. A History of Pepsin and Related Enzymes. Q. Rev. Biol. 2002, 77, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, E.A.; Zaichik, B.T.; Eremin, S.A.; Zherdev, A.V.; Dzantiev, B.B. Enzyme immunoassay for detection of Sudan I dye and its application to the control of foodstuffs. J. Anal. Chem. 2016, 71, 944–948. [Google Scholar] [CrossRef]
- Psochiou, E.; Sarropoulou, E.; Mamuris, Z.; Moutou, K.A. Sequence analysis and tissue expression pattern of Sparus aurata chymotrypsinogens and trypsinogen. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 147, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Darias, M.J.; Murray, H.M.; Gallant, J.W.; Douglas, S.E.; Yúfera, M.; Martínez-Rodríguez, G. The spatiotemporal expression pattern of trypsinogen and bile salt-activated lipase during the larval development of red porgy (Pagrus pagrus, Pisces, Sparidae). Mar. Biol. 2007, 152, 109–118. [Google Scholar] [CrossRef]
- Ohshima, Y.; Suzuki, Y.; Nakatani, A.; Nohara, D. Refolding of Fully Reduced Bovine Pancreatic Trypsin. J. Biosci. Bioeng. 2008, 106, 345–349. [Google Scholar] [CrossRef]
- Hidalgo, M.C.; Urea, E.; Sanz, A. Comparative study of digestive enzymes in fish with different nutritional habits. Proteolytic and amylase activities. Aquaculture 1999, 170, 267–283. [Google Scholar] [CrossRef]
- Kumar, V. Enzymes in Human and Animal Nutrition Principles and Perspectives; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Anthonsen, H.W.; Baptista, A.; Drabls, F.; Martel, P.; Petersen, S.B.; Sebastio, M.; Vaz, L. Lipases and esterases: A review of their sequences, structure and evolution—Sciencedirect. Biotechnol. Annu. Rev. 1995, 1, 315–371. [Google Scholar] [CrossRef]
- Sandoval, G. Chemoen-zymatic Synthesis of Nitrogen Polymers with Biomedical Applications Catalyzed by Lipases: An overview. In Lipases and Phospholipases (Methods and Protocols); Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1835, pp. 359–376. [Google Scholar] [CrossRef]
- Guo, J.P.; Wang, J.; Li, H.J.; Ma, T.; Qi, Z.G.; Hao, Z.L.; Chen, J.Y. Effect of different weaning ages on the main digestive enzyme activities in the small intestinal contents of lamb. Chin. Anim. Husb. Vet. Med. Intest. Contents Lamb 2017, 44, 2603–2612. [Google Scholar]
- Gheisvandi, N.; Hajimoradloo, A.; Ghorbani, R.; Hoseinifar, S.H. The effects of gradual or abrupt changes in salinity on digestive enzymes activity of Caspian kutum, Rutilus kutum (Kamensky, 1901) larvae. J. Appl. Ichthyol. 2015, 31, 1107–1112. [Google Scholar] [CrossRef]
- Navarro-Guillén, C.; Yúfera, M.; Perera, E. Biochemical features and modulation of digestive enzymes by environmental temperature in the greater amberjack, Seriola dumerili. Front. Mar. Sci. 2022, 9, 960746. [Google Scholar] [CrossRef]
- Hieu, D.Q.; Hang, B.T.B.; Huong, D.T.T.; El Kertaoui, N.; Farnir, F.; Phuong, N.T.; Kestemont, P. Salinity affects growth performance, physiology, immune responses and temperature resistance in striped catfish (Pangasianodon hypophthalmus) during its early life stages. Fish Physiol. Biochem. 2021, 47, 1995–2013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-F.; Gao, X.-Q.; Yu, J.-X.; Qian, X.-M.; Xue, G.-P.; Zhang, Q.-Y.; Liu, B.-L.; Hong, L. Effects of different salinities on growth performance, survival, digestive enzyme activity, immune response, and muscle fatty acid composition in juvenile American shad (Alosa sapidissima). Fish Physiol. Biochem. 2016, 43, 761–773. [Google Scholar] [CrossRef]
- Hamed, S.S.; Jiddawi, N.S.; Poj, B. Effect of salinity levels on growth, feed utilization, body composition and digestive enzymes activities of juvenile silver pompano Trachinotus blochii. AkiNik Publ. 2016, 4, 279–283. [Google Scholar]
- Squires, E.J.; Haard, N.F.; Feltham, L.A.W. Gastric proteases of the Greenland cod Gadus ogac. I. Isolation and kinetic properties. Biochem. Cell Biol. 1986, 64, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Lin, H. Fish Physiology; Guangdong Higher Education Press: Guangzhou, China, 2011; pp. 213–219. [Google Scholar]
- Lee-Shing, F.; Shu-Fen, C. Effect of salinity on the activities of digestive proteases from the tilapia fish, Oreochromis niloticus in different culture environments. Comp. Biochem. Physiol. Part A Physiol. 1989, 93, 439–443. [Google Scholar] [CrossRef]
- Noda, M.; Murakami, K. Studies on proteinases from the digestive organs of sardine. II. Purification and characterization of two acid proteinases from the stomach. Biochim. Biophys. Acta (BBA) Enzym. 1981, 658, 27–34. [Google Scholar] [CrossRef]
- Buentello, J.A.; Pohlenz, C.; Margulies, D.; Scholey, V.P.; Wexler, J.B.; Tovar-Ramírez, D.; Neill, W.H.; Hinojosa-Baltazar, P.; Gatlin, D.M. A preliminary study of digestive enzyme activities and amino acid composition of early juvenile yellowfin tuna (Thunnus albacares). Aquaculture 2011, 312, 205–211. [Google Scholar] [CrossRef]
- Kihara, M. Pepsin-like protease activity and the gastric digestion within ex vivo Pacific bluefin tuna Thunnus orientalis stomachs, as a gastric digestion model. Anim. Feed. Sci. Technol. 2015, 206, 87–99. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Sharma, J.G. Digestive physiology of fish larvae during ontogenic development: A brief overview. Indian J. Anim. Sci. 2005, 75, 1337–1347. [Google Scholar] [CrossRef]
- Kolkovski, S. Digestive enzymes in fish larvae and juveniles—Implications and applications to formulated diets. Aquaculture 2001, 200, 181–201. [Google Scholar] [CrossRef]
- Zhou, S.J.; Zhang, N.L.; Fu, Z.Y.; Yu, G.; Ma, Z.H.; Zhao, L. Impact of Salinity Changes on the Antioxidation of Juvenile Yellowfin Tuna (Thunnus albacares). J. Mar. Sci. Eng. 2023, 11, 132. [Google Scholar] [CrossRef]
- Li, X.J.; Shen, Y.D.; Bao, Y.G.; Wu, Z.X.; Yang, B.Q.; Jiao, L.F.; Zhang, C.D.; Tocher, D.R.; Zhou, Q.C.; Jin, M. Physiological responses and adaptive strategies to acute low-salinity environmental stress of the euryhaline marine fish black seabream (Acanthopagrus schlegelii). Aquaculture 2022, 554, 738117. [Google Scholar] [CrossRef]
- Mozanzadeh, M.T.; Safari, O.; Oosooli, R.; Mehrjooyan, S.; Najafabadi, M.Z.; Hoseini, S.J.; Saghavi, H.; Monem, J. The effect of salinity on growth performance, digestive and antioxidant enzymes, humoral immunity and stress indices in two euryhaline fish species: Yellowfin seabream (Acanthopagrus latus) and Asian seabass (Lates calcarifer). Aquaculture 2020, 534, 736329. [Google Scholar] [CrossRef]
- Kawai, S.-I.; Ikeda, S. Studies on digestive enzymes of fishes Ⅱ. Effect of dietary change on activities of digestive enzymes in carp intestine. Bull. Jpn. Soc. Sci. Fish. 1972, 38, 265–270. [Google Scholar] [CrossRef]
- Sánchez-Chiang, L.; Cisternas, E.; Ponce, O. Partial purification of pepsins from adult and juvenile salmon fish Oncorhynchus keta. Effect of NaCl on proteolytic activities. Comp. Biochem. Physiol. Part B Comp. Biochem. 1987, 87, 793–797. [Google Scholar] [CrossRef]
- Luo, M.Z.; Guan, R.Z.; Jin, H. Effects of the salinity on the growth performance and digestive enzyme activities of Anguilla marmorata elver and A. bicolor pacifica elver. Acta Hydrobiol. Sin. 2015, 39, 653–660. [Google Scholar]
- Chiu, Y.N.; Benitez, L.V. Studies on the carbohydrases in the digestive tract of the milkfish chanos chanos. Mar. Biol. 1981, 61, 247–254. [Google Scholar] [CrossRef]
- Soltan, N.M.; Soaudy, M.R.; Abdella, M.M.; Hassaan, M.S. Partial dietary fishmeal replacement with mixture of plant protein sources supplemented with exogenous enzymes modify growth performance, digestibility, intestinal morphology, haemato-biochemical and immune responses for Nile tilapia, Oreochromis niloticus. Anim. Feed. Sci. Technol. 2023, 299, 115642. [Google Scholar] [CrossRef]
- Zhi, B.J.; Liu, W.; Zhao, C.G.; Duan, Y.Y. Effects of salinity on digestive enzyme and alkaline phosphatase activity of young chum salmon (Oncorhynchus keta Walbaum). J. Shanghai Ocean. Univ. 2009, 31, 627–632. [Google Scholar]
- Yang, J.W.; Zhou, Y.G.; Huang, M.; Xiong, Y.H.; Wang, F.; Gao, Q.F.; Sun, D.J.; Dong, S.L. Comparative studies on the digestive and antioxidant enzyme activities between juvenile rainbow (Oncorhynchus mykiss) and steelhead troutof rainbow trout (O. mykiss). J. Ocean. Univ. China 2019, 49, 119–128. [Google Scholar]
- Zhang, L.G.; An, L.; Sun, D.; Fu, P.S. Effects of salinity on digestive enzyme activities of juvenile Jade Perch Scortum barcoo. Chin. J. Fish. 2021, 24, 21–24. [Google Scholar]
- Rivera-Ingraham, G.A.; Lignot, J.-H. Osmoregulation, bioenergetics and oxidative stress in coastal marine invertebrates: Raising the questions for future research. J. Exp. Biol. 2017, 220, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.S.; Faleiro, F.; Diniz, M.; Machado, J.; Pousão-Ferreira, P.; Peck, M.A.; Pörtner, H.O.; Rosa, R. Oxidative Stress and Digestive Enzyme Activity of Flatfish Larvae in a Changing Ocean. PLoS ONE 2015, 10, e0134082. [Google Scholar] [CrossRef]
- Pujante, I.M.; Moyano, F.J.; Martos-Sitcha, J.A.; Mancera, J.M.; Martínez-Rodríguez, G. Effect of different salinities on gene expression and activity of digestive enzymes in the thick-lipped grey mullet (Chelon labrosus). Fish Physiol. Biochem. 2017, 44, 349–373. [Google Scholar] [CrossRef]
- Yu, L.J.; Wen, H.; Jiang, M.; Wu, F.; Tian, J.; Lu, X.; Xiao, J.R.; Liu, W. Effects of ferulic acid on intestinal enzyme activities, morphology, microbiome composition of genetically improved farmed tilapia (Oreochromis niloticus) fed oxidized fish oil. Aquaculture 2020, 528, 735543. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, S.; Dong, X.H.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y.; Tan, B.P.; Xie, S.W. Effects of fishmeal replacement by black soldier fly on growth performance, digestive enzyme activity, intestine morphology, intestinal flora and immune response of pearl gentian grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Fish Shellfish. Immunol. 2022, 120, 497–506. [Google Scholar] [CrossRef]
- Pe Tan, P.; Zhu, W.L.; Zhang, P.; Wang, L.G.; Chen, R.Y.; Xu, D.D. Dietary soybean lecithin inclusion promotes growth, development, and intestinal morphology of yellow drum (Nibea albiflora) larvae. Aquaculture 2022, 559, 738446. [Google Scholar] [CrossRef]
- Chen, J.; Xu, P.; Wen, H.; Xue, M.; Wang, Q.; He, J.; He, C.; Su, S.; Li, J.; Yu, F.; et al. Hypothermia-mediated oxidative stress induces immunosuppression, morphological impairment and cell fate disorder in the intestine of freshwater drum, Aplodinotus grunniens. Aquaculture 2023, 575, 739805. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Yang, R.; Fu, Z.; Yu, G.; Ma, Z. Mechanisms of Digestive Enzyme Response to Acute Salinity Stress in Juvenile Yellowfin Tuna (Thunnus albacares). Animals 2023, 13, 3454. https://doi.org/10.3390/ani13223454
Zhang N, Yang R, Fu Z, Yu G, Ma Z. Mechanisms of Digestive Enzyme Response to Acute Salinity Stress in Juvenile Yellowfin Tuna (Thunnus albacares). Animals. 2023; 13(22):3454. https://doi.org/10.3390/ani13223454
Chicago/Turabian StyleZhang, Ninglu, Rui Yang, Zhengyi Fu, Gang Yu, and Zhenhua Ma. 2023. "Mechanisms of Digestive Enzyme Response to Acute Salinity Stress in Juvenile Yellowfin Tuna (Thunnus albacares)" Animals 13, no. 22: 3454. https://doi.org/10.3390/ani13223454
APA StyleZhang, N., Yang, R., Fu, Z., Yu, G., & Ma, Z. (2023). Mechanisms of Digestive Enzyme Response to Acute Salinity Stress in Juvenile Yellowfin Tuna (Thunnus albacares). Animals, 13(22), 3454. https://doi.org/10.3390/ani13223454