
Identification of Lipids and Cytokines in Plasma and Follicular Fluid before and after Follicle-Stimulating Hormone Stimulation as Potential Markers for Follicular Maturation in Cattle
 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Estrous Synchronization and Stimulation
2.3. Blood Collections
2.4. Ovariectomies
2.5. Steroid Hormone Assays
2.6. Lipidomic Analysis
2.7. Cytokine Analysis
2.8. Statistical Analysis
3. Results
3.1. Steroid Profile before and after FSH Stimulation
3.2. Plasma Lipidomic Profiles before and after FSH Stimulation
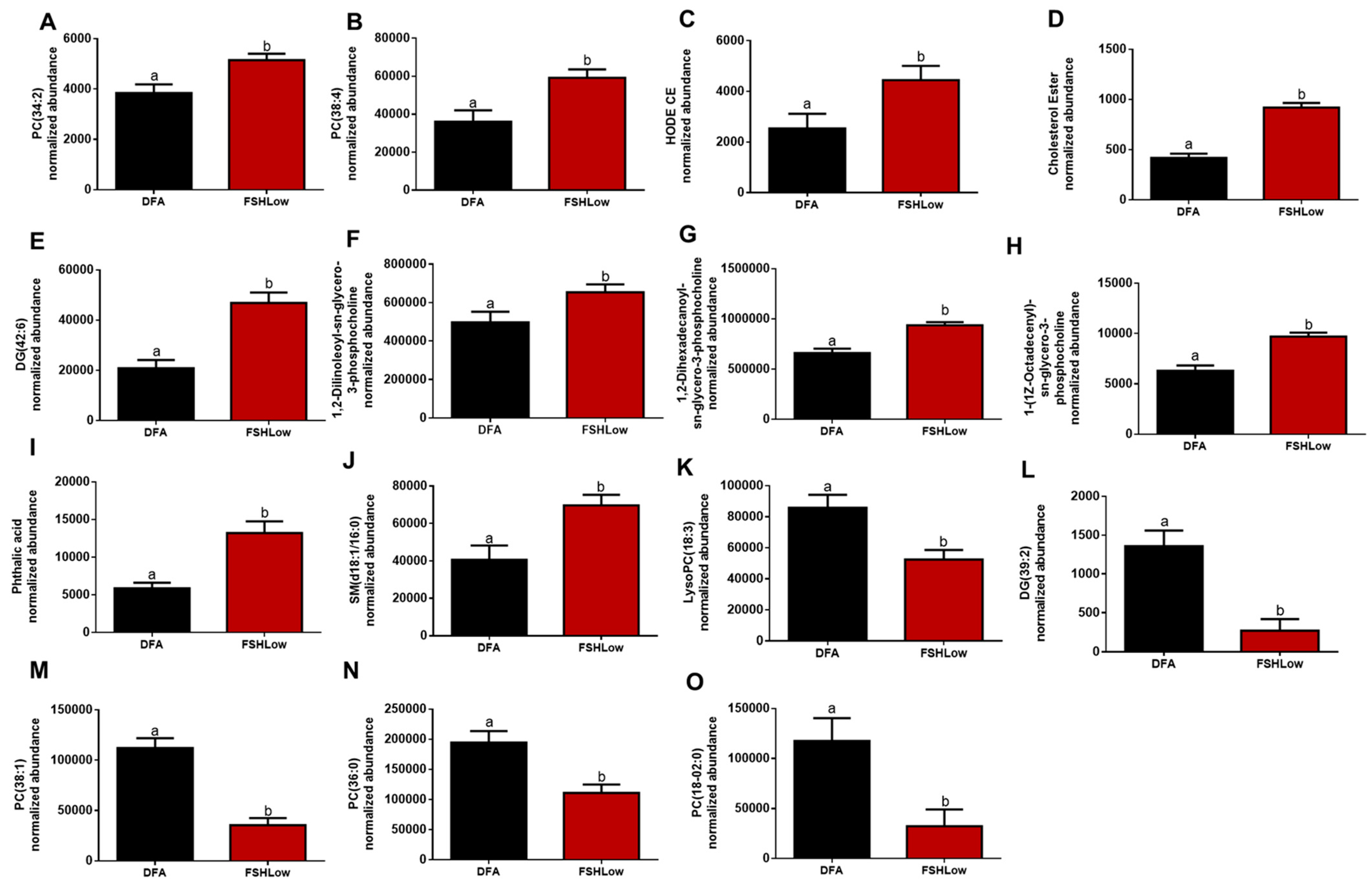
3.3. Follicular Fluid Lipidomic Profiles before and after FSH Stimulation
3.4. Plasma and Follicular Fluid Correlations of Lipids before and after FSH Stimulation
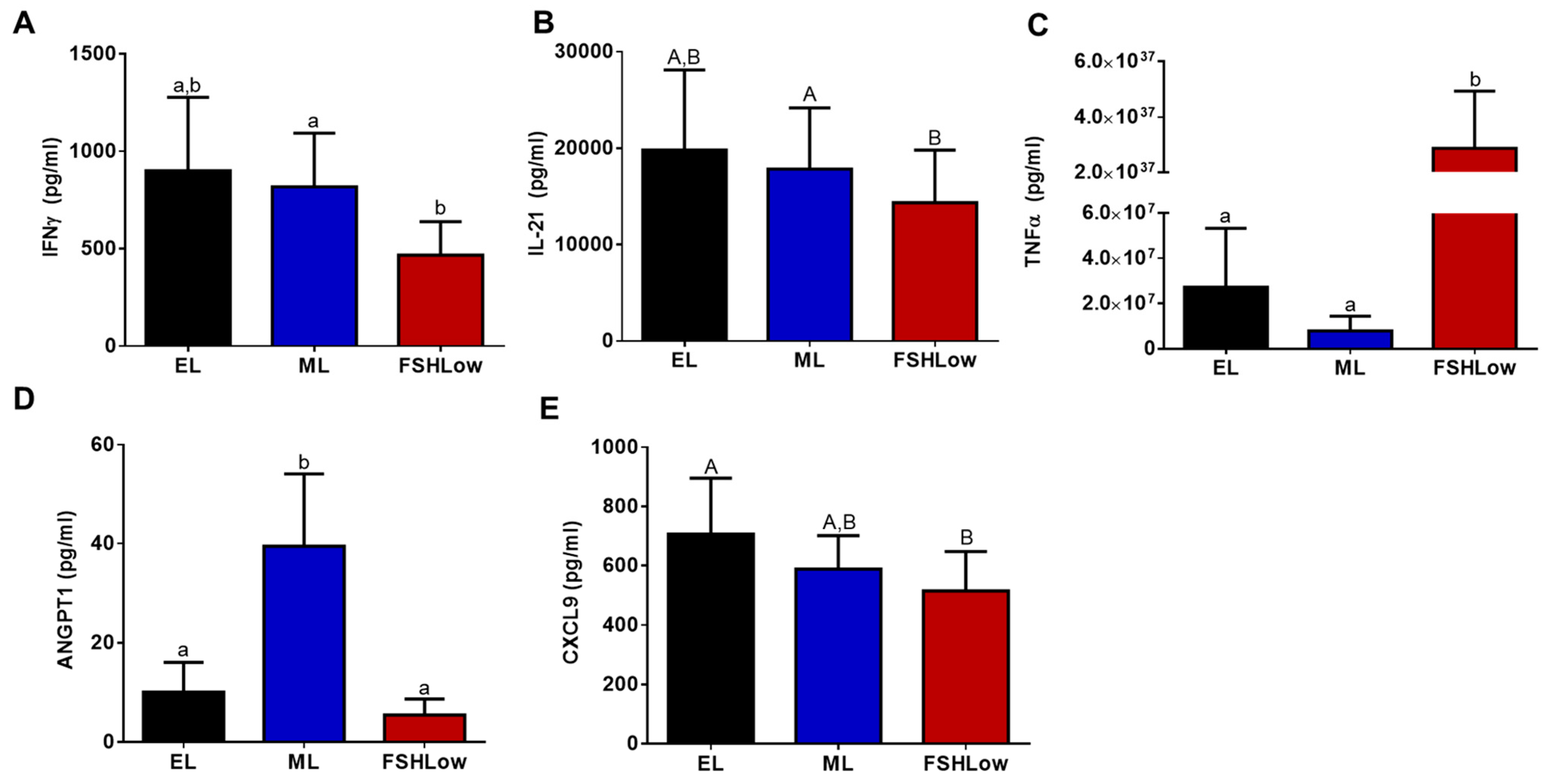
3.5. Plasma and Follicular Fluid Cytokine Profiles before and after FSH Stimulation
3.6. Plasma and Follicular Fluid Correlations of Cytokines and Circulating Steroids before and after FSH Stimulation
4. Discussion
4.1. Lipid and Cytokine Profiles during Early to Mid-Luteal Phase
4.2. Lipid and Cytokine Profiles after FSHLow
4.3. Follicular Fluid Lipid and Cytokine Profiles after FSHLow
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hambridge, H.L.; Mumford, S.L.; Mattison, D.R.; Ye, A.; Pollack, A.Z.; Bloom, M.S.; Mendola, P.; Lynch, K.L.; Wactawski-Wende, J.; Schisterman, E.F. The influence of sporadic anovulation on hormone levels in ovulatory cycles. Hum. Reprod. 2013, 28, 1687–1694. [Google Scholar] [CrossRef]
- Veiga-Lopez, A.; Gonzalez-Bulnes, A.; Tresguerres, J.A.; Dominguez, V.; Ariznavarreta, C.; Cocero, M.J. Causes, characteristics and consequences of anovulatory follicles in superovulated sheep. Domest. Anim. Endocrinol. 2006, 30, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Wiltbank, M.C.; Gümen, A.; Sartori, R. Physiological classification of anovulatory conditions in cattle. Theriogenology 2002, 57, 21–52. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.R.; van den Hurk, R.; Figueiredo, J.R. Ovarian follicle development in vitro and oocyte competence: Advances and challenges for farm animals. Domest. Anim. Endocrinol. 2016, 55, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.R.V.; Lima, F.E.O.; Souza, A.L.P.; Silva, A.W.B. Interleukin-1beta and TNF-alpha systems in ovarian follicles and their roles during follicular development, oocyte maturation and ovulation. Zygote 2020, 28, 270–277. [Google Scholar] [CrossRef]
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E. Ovulation: Parallels with Inflammatory Processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef]
- Gareis, N.C.; Angeli, E.; Huber, E.; Salvetti, N.R.; Rodríguez, F.M.; Ortega, H.H.; Hein, G.J.; Rey, F. Alterations in key metabolic sensors involved in bovine cystic ovarian disease. Theriogenology 2018, 120, 138–146. [Google Scholar] [CrossRef]
- Ribeiro, E.S. Symposium review: Lipids as regulators of conceptus development: Implications for metabolic regulation of reproduction in dairy cattle1. J. Dairy Sci. 2018, 101, 3630–3641. [Google Scholar] [CrossRef]
- Santos, J.E.P.; Bilby, T.R.; Thatcher, W.W.; Staples, C.R.; Silvestre, F.T. Long Chain Fatty Acids of Diet as Factors Influencing Reproduction in Cattle. Reprod. Domest. Anim. 2008, 43, 23–30. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Z.; Shen, W.J.; Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. 2010, 7, 47. [Google Scholar] [CrossRef]
- Talbott, H.A.; Plewes, M.R.; Krause, C.; Hou, X.; Zhang, P.; Rizzo, W.B.; Wood, J.R.; Cupp, A.S.; Davis, J.S. Formation and characterization of lipid droplets of the bovine corpus luteum. Sci. Rep. 2020, 10, 11287. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Spicer, L.J.; Echternkamp, S.E. Ovarian Follicular Growth, Function and Turnover in Cattle: A Review. J. Anim. Sci. 1986, 62, 428–451. [Google Scholar] [CrossRef]
- Mavangira, V.; Sordillo, L.M. Role of lipid mediators in the regulation of oxidative stress and inflammatory responses in dairy cattle. Res. Vet. Sci. 2018, 116, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Denis, U.; Lecomte, M.; Paget, C.; Ruggiero, D.; Wiernsperger, N.; Lagarde, M. Advanced glycation end-products induce apoptosis of bovine retinal pericytes in culture: Involvement of diacylglycerol/ceramide production and oxidative stress induction. Free Radic. Biol. Med. 2002, 33, 236–247. [Google Scholar] [CrossRef]
- Glass, C.K.; Olefsky, J.M. Inflammation and lipid signaling in the etiology of insulin resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An updated review of lysophosphatidylcholine metabolism in human diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed]
- Sadik, C.D.; Luster, A.D. Lipid-cytokine-chemokine cascades orchestrate leukocyte recruitment in inflammation. J. Leukoc. Biol. 2012, 91, 207–215. [Google Scholar] [CrossRef]
- Treede, I.; Braun, A.; Sparla, R.; Kühnel, M.; Giese, T.; Turner, J.R.; Anes, E.; Kulaksiz, H.; Füllekrug, J.; Stremmel, W.; et al. Anti-inflammatory effects of phosphatidylcholine. J. Biol. Chem. 2007, 282, 27155–27164. [Google Scholar] [CrossRef] [PubMed]
- Welty, F.K. How do elevated triglycerides and low HDL-cholesterol affect inflammation and atherothrombosis? Curr. Cardiol. Rep. 2013, 15, 400. [Google Scholar] [CrossRef]
- Kelly, P.; Duffy, P.; Roche, J.F.; Boland, M.P. Superovulation in cattle: Effect of FSH type and method of administration on follicular growth, ovulatory response and endocrine patterns. Anim. Reprod. Sci. 1997, 46, 1–14. [Google Scholar] [CrossRef]
- McFee, R.M.; Romereim, S.M.; Snider, A.P.; Summers, A.F.; Pohlmeier, W.E.; Kurz, S.G.; Cushman, R.A.; Davis, J.S.; Wood, J.R.; Cupp, A.S. A high-androgen microenvironment inhibits granulosa cell proliferation and alters cell identity. Mol. Cell. Endocrinol. 2021, 531, 111288. [Google Scholar] [CrossRef]
- Summers, A.F.; Pohlmeier, W.E.; Sargent, K.M.; Cole, B.D.; Vinton, R.J.; Kurz, S.G.; McFee, R.M.; Cushman, R.A.; Cupp, A.S.; Wood, J.R. Altered Theca and Cumulus Oocyte Complex Gene Expression, Follicular Arrest and Reduced Fertility in Cows with Dominant Follicle Follicular Fluid Androgen Excess. PLoS ONE 2014, 9, e110683. [Google Scholar] [CrossRef]
- Youngquist, R.; Garverick, H.; Keisler, D. Use of umbilical cord clamps for ovariectomy in cows. J. Am. Vet. Med. Assoc. 1995, 207, 474–475. [Google Scholar] [PubMed]
- Kojima, N.; Stumpf, T.T.; Cupp, A.S.; Werth, L.A.; Roberson, M.S.; Wolfe, M.W.; Kittok, R.J.; Kinder, J.E. Exogenous progesterone and progestins as used in estrous synchrony regimens do not mimic the corpus luteum in regulation of luteinizing hormone and 17β-estradiol in circulation of cows. Biol. Reprod. 1992, 47, 1009–1017. [Google Scholar] [CrossRef]
- Walsh, S.C.; Miles, J.R.; Yao, L.; Broeckling, C.D.; Rempel, L.A.; Wright-Johnson, E.C.; Pannier, A.K. Metabolic compounds within the porcine uterine environment are unique to the type of conceptus present during the early stages of blastocyst elongation. Mol. Reprod. Dev. 2020, 87, 174–190. [Google Scholar] [CrossRef]
- Poole, R.K.; Brown, A.R.; Poore, M.H.; Pickworth, C.L.; Poole, D.H. Effects of endophyte- infected tall fescue seed and protein supplementation on stocker steers: II. Adaptive and innate immune function. J. Anim. Sci. 2019, 97, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fang, S.; Wang, Y.; Chi, X.; Ma, X.; Zhang, T.; Hu, S. Receptor and signaling pathway involved in bovine lymphocyte activation by Atractylodis macrocephalae polysaccharides. Carbohydr. Polym. 2020, 234, 115906. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D.; Afsar, F.A.; Neumann, S.; Ben-Hur, A.; Prenni, J.E. RAMClust: A novel feature clustering method enables spectral-matching-based annotation for metabolomics data. Anal. Chem. 2014, 86, 6812–6817. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B. (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Shelton, K.; Gayerie De Abreu, M.F.; Hunter, M.G.; Parkinson, T.J.; Lamming, G.E. Luteal inadequacy during the early luteal phase of subfertile cows. J. Reprod. Fertil. 1990, 90, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Welte, M.A.; Gould, A.P. Lipid droplet functions beyond energy storage. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1260–1272. [Google Scholar] [CrossRef]
- Divecha, N.; Banfic, H.; Irvine, R.F. The polyphosphoinositide cycle exists in the nuclei of Swiss 3T3 cells under the control of a receptor (for IGF-I) in the plasma membrane, and stimulation of the cycle increases nuclear diacylglycerol and apparently induces translocation of protein kinase C to the nucleus. EMBO J. 1991, 10, 3207–3214. [Google Scholar] [PubMed]
- Massart, J.; Zierath, J.R. Role of Diacylglycerol Kinases in Glucose and Energy Homeostasis. Trends Endocrinol. Metab. 2019, 30, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.A. Changes in lipid contents and fatty acid compositions in ovine corpora lutea during the estrous cycle and early pregnancy. Biol. Reprod. 1988, 38, 605–615. [Google Scholar] [CrossRef]
- Ye, W.; Zhang, L. Heme controls the expression of cell cycle regulators and cell growth in HeLa cells. Biochem. Biophys. Res. Commun. 2004, 315, 546–554. [Google Scholar] [CrossRef]
- Rivett, E.D.; Heo, L.; Feig, M.; Hegg, E.L. Biosynthesis and trafficking of heme o and heme a: New structural insights and their implications for reaction mechanisms and prenylated heme transfer. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 640–668. [Google Scholar] [CrossRef]
- Legrand-Poels, S.; Esser, N.; L’Homme, L.; Scheen, A.; Paquot, N.; Piette, J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem. Pharacol. 2014, 92, 131–141. [Google Scholar] [CrossRef]
- Brindle, N.P.; Saharinen, P.; Alitalo, K. Signaling and functions of angiopoietin-1 in vascular protection. Circ. Res. 2006, 98, 1014–1023. [Google Scholar] [CrossRef]
- Fiedler, U.; Augustin, H.G. Angiopoietins: A link between angiogenesis and inflammation. Trends Immunol. 2006, 27, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Bowolaksono, A.; Fauzi, M.; Sundari, A.M.; Pustimbara, A.; Lestari, R.; Abinawanto; Dwiranti, A.; Fadhillah. The effects of luteinizing hormone as a suppression factor for apoptosis in bovine luteal cells in vitro. Reprod. Domest. Anim. 2021, 56, 744–753. [Google Scholar] [CrossRef]
- Mumford, S.L.; Browne, R.W.; Schliep, K.C.; Schmelzer, J.; Plowden, T.C.; Michels, K.A.; Sjaarda, L.A.; Zarek, S.M.; Perkins, N.J.; Messer, L.C.; et al. Serum Antioxidants Are Associated with Serum Reproductive Hormones and Ovulation among Healthy Women. J. Nutr. 2016, 146, 98–106. [Google Scholar] [CrossRef]
- Abedal-Majed, M.A.; Springman, S.A.; Sutton, C.M.; Snider, A.P.; Bell, B.E.; Hart, M.; Kurz, S.G.; Bergman, J.; Summers, A.F.; McFee, R.M.; et al. VEGFA165 can rescue excess steroid secretion, inflammatory markers, and follicle arrest in the ovarian cortex of High A4 cowsdagger. Biol. Reprod. 2022, 106, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Rekawiecki, R.; Kowalik, M.K.; Slonina, D.; Kotwica, J. Regulation of progesterone synthesis and action in bovine corpus luteum. J. Physiol. Pharmacol. 2008, 59 (Suppl. S9), 75–89. [Google Scholar]
- Fields, M.J.; Fields, P.A. Morphological characteristics of the bovine corpus luteum during the estrous cycle and pregnancy. Theriogenology 1996, 45, 1295–1325. [Google Scholar] [CrossRef] [PubMed]
- Gwynne, J.T.; Strauss, J.F. The Role of Lipoproteins in Steroidogenesis and Cholesterol Metabolism in Steroidogenic Glands. Endocr. Rev. 1982, 3, 299–329. [Google Scholar] [CrossRef] [PubMed]
- Eichmann, T.O.; Lass, A. DAG tales: The multiple faces of diacylglycerol--stereochemistry, metabolism, and signaling. Cell. Mol. Life Sci. 2015, 72, 3931–3952. [Google Scholar] [CrossRef]
- Bradford, B.J.; Mamedova, L.K.; Minton, J.E.; Drouillard, J.S.; Johnson, B.J. Daily injection of tumor necrosis factor-{alpha} increases hepatic triglycerides and alters transcript abundance of metabolic genes in lactating dairy cattle. J. Nutr. 2009, 139, 1451–1456. [Google Scholar] [CrossRef]
- Lalanne, F.; Pruneta, V.; Bernard, S.; Ponsin, G. Distribution of diacylglycerols among plasma lipoproteins in control subjects and in patients with non-insulin-dependent diabetes. Eur. J. Clin. Investig. 1999, 29, 139–144. [Google Scholar] [CrossRef]
- Macrae, K.; Stretton, C.; Lipina, C.; Blachnio-Zabielska, A.; Baranowski, M.; Gorski, J.; Marley, A.; Hundal, H.S. Defining the role of DAG, mitochondrial function, and lipid deposition in palmitate-induced proinflammatory signaling and its counter-modulation by palmitoleate. J. Lipid Res. 2013, 54, 2366–2378. [Google Scholar] [CrossRef]
- Toya, M.; Hozumi, Y.; Ito, T.; Takeda, M.; Sakane, F.; Kanoh, H.; Saito, H.; Hiroi, M.; Kurachi, H.; Kondo, H.; et al. Gene expression, cellular localization, and enzymatic activity of diacylglycerol kinase isozymes in rat ovary and placenta. Cell Tissue Res. 2005, 320, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Klausen, C.; Severson, D.L.; Chang, J.P.; Habibi, H.R. Role of PKC in the regulation of gonadotropin subunit mRNA levels: Interaction with two native forms of gonadotropin-releasing hormone. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1634–R1643. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hirota, K.; Hirota, T.; Aguilera, G.; Catt, K.J. Hormone-induced redistribution of calcium-activated phospholipid-dependent protein kinase in pituitary gonadotrophs. J. Biol. Chem. 1985, 260, 3243–3246. [Google Scholar] [CrossRef] [PubMed]
- Azhar, S.; Revan, E. Scavenger receptor class BI and selective cholesteryl ester uptake: Partners in the regulation of steroidogenesis. Mol. Cell. Endocrinol. 2002, 195, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L.; Bose, H.S. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res. 2011, 52, 2111–2135. [Google Scholar] [CrossRef]
- Hiley, C.R.; Hoi, P.M. Oleamide: A fatty acid amide signaling molecule in the cardiovascular system? Cardiovasc. Drug Rev. 2007, 25, 46–60. [Google Scholar] [CrossRef]
- Abedel-Majed, M.A.; Romereim, S.M.; Davis, J.S.; Cupp, A.S. Perturbations in Lineage Specification of Granulosa and Theca Cells May Alter Corpus Luteum Formation and Function. Front. Endocrinol. 2019, 10, 832. [Google Scholar] [CrossRef]
- Girard, A.; Dufort, I.; Douville, G.; Sirard, M.A. Global gene expression in granulosa cells of growing, plateau and atretic dominant follicles in cattle. Reprod. Biol. Endocrinol. 2015, 13, 17. [Google Scholar] [CrossRef]
- Dutra, F.F.; Bozza, M.T. Heme on innate immunity and inflammation. Front. Pharmacol. 2014, 5, 115. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Mühl, H.; Pfeilschifter, J. Anti-inflammatory properties of pro-inflammatory interferon-γ. Int. Immunopharmacol. 2003, 3, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte environment: Follicular fluid and cumulus cells are critical for oocyte health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Ferrazza, R.A.; Garcia, H.D.M.; Schmidt, E.; Mihm Carmichael, M.; Souza, F.F.; Burchmore, R.; Sartori, R.; Eckersall, P.D.; Ferreira, J.C.P. Quantitative proteomic profiling of bovine follicular fluid during follicle development. Biol. Reprod. 2017, 97, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pena, L.A.; Fuksf, Z.; Kolesnick, R. Stress-Induced Apoptosis and the Sphingomyelin Pathway. Biochem. Pharacol. 1997, 53, 615–621. [Google Scholar] [CrossRef]
- Subbaiah, P.V.; Sargis, R.M. Sphingomyelin: A natural modulator of membrane homeostasis and inflammation. Med. Hypothesis 2001, 57, 135–138. [Google Scholar] [CrossRef]
- Cutler, R.G.; Mattson, M.P. Sphingomyelin and ceramide as regulators of development and lifespan. Mech. Ageing Dev. 2001, 122, 895–908. [Google Scholar] [CrossRef]
- Milhas, D.; Clarke, C.J.; Hannun, Y.A. Sphingomyelin metabolism at the plasma membrane: Implications for bioactive sphingolipids. FEBS Lett. 2010, 584, 1887–1894. [Google Scholar] [CrossRef]
- Abdulrahman Alrabiah, N.; Simintiras, C.A.; Evans, A.C.O.; Lonergan, P.; Fair, T. Biochemical alterations in the follicular fluid of bovine peri-ovulatory follicles and association with final oocyte maturation. Reprod. Fertil. 2022, 4, 220090. [Google Scholar] [CrossRef]
- Santos, P.H.; Fontes, P.K.; Franchi, F.F.; Nogueira, M.F.; Belaz, K.R.; Tata, A.; Everlin, M.N.; Sudano, M.J.; Barros, C.M.; Castilho, A.C. Lipid profiles of follicular fluid from cows submitted to ovarian superstimulation. Theriogenology 2017, 94, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Bertevello, P.S.; Teixeira-Gomes, A.P.; Labas, V.; Cordeiro, L.; Blache, M.C.; Papillier, P.; Singina, G.; Uzbekov, R.; Maillard, V.; Uzbekova, S. MALDI-TOF mass spectrometry revealed significant lipid variations in follicular fluid and somatic follicular cells but not in enclosed oocytes between the large dominant and small subordinate follicles in bovine ovary. Int. J. Mol. Sci. 2020, 21, 6661. [Google Scholar] [CrossRef]
- da Silveria, J.C.; Andrade, G.M.; Simas, R.C.; Martins-Junior, H.A.; Eberlin, M.S.; Smith, L.C.; Perecin, F.; Meirelles, F.V. Lipid Profile of extracellular vesicles and their relationship with bovine oocyte developmental competence: New players in intra follicular cell communication. Theriogenology 2021, 174, 1–8. [Google Scholar] [CrossRef]
- Melo, E.O.; Cordeiro, D.M.; Pellegrino, R.; Wei, Z.; Daye, Z.J.; Nishimura, R.C.; Dode, M.A. Identification of molecular markers for oocyte competence in bovine cumulus cells. Anim. Genet. 2017, 48, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Snider, A.P.; Wood, J.R. Obesity induces ovarian inflammation and reduces oocyte quality. Reproduction 2019, 158, R79–R90. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma | |||||
|---|---|---|---|---|---|
| DFA | FSHLow | ||||
| Lipid Name | R2 | p-Value | R2 | p-Value | |
| E2 | TG 54:6 | −0.83 | 0.02 | ||
| TG 58:6 | −0.80 | 0.03 | |||
| HemeA | −0.80 | 0.04 | |||
| A4 | Cholesteryl-11-hydroperoxy-eicosateraenoate | 0.87 | 0.01 | ||
| DG 38:0 | 0.79 | 0.04 | |||
| TG 49:1 | 0.79 | 0.04 | |||
| TG 50:0 | 0.78 | 0.05 | |||
| TG 54:6 | 0.80 | 0.05 | |||
| TG 58:6 | 0.85 | 0.01 | |||
| P4 | PC 38:2 | 0.78 | 0.05 | ||
| 3-Deoxyvitamin | 0.82 | 0.02 | |||
| Oleamide | 0.87 | 0.01 | |||
| sn-glycero-3-phosphoserine | 0.82 | 0.02 | |||
| CE 16:0 | 0.88 | 0.02 | |||
| C.E. | 0.80 | 0.03 | |||
| HODE | 0.84 | 0.02 | |||
| DG 16:0–18:0 | 0.77 | 0.05 | |||
| DG 42:6 | 0.86 | 0.01 | |||
| TG 52:0 | 0.77 | 0.05 | |||
| 16:0 SM (d18:1/16:0) | 0.87 | 0.01 | |||
| Diolein | 0.83 | 0.02 | |||
| HemeA | 0.83 | 0.02 | |||
| Follicular Fluid | |||||
|---|---|---|---|---|---|
| DFA | FSHLow | ||||
| Lipid Name | R2 | p-Value | R2 | p-Value | |
| E2 | 1,2-dilinoleoyl sn-glycero-3-phosphocholine | −0.96 | 0.01 | ||
| Lyso-PC20:4 | −0.95 | 0.02 | |||
| Phthalic acid | −0.79 | 0.04 | |||
| HemeA | −0.87 | 0.01 | |||
| A4 | PC34:2 | −0.94 | 0.02 | ||
| PC36:0 | −0.97 | 0.004 | |||
| sodium glycochenodeoxycholate | −0.81 | 0.03 | |||
| DG42:6 | 0.86 | 0.01 | |||
| 1-oleoyl-2-hydroxy-sn-glycero-3-phosphocholine | 0.90 | 0.05 | |||
| P4 | Phthalic acid | 0.91 | 0.04 | ||
| 1,2-dilinoleoyl sn-glycero-3-phosphocholine | 0.83 | 0.02 | |||
| Plasma | |||||||
|---|---|---|---|---|---|---|---|
| EL | ML | FSHLow | |||||
| Cytokine | R2 | p-Value | R2 | p-Value | R2 | p-Value | |
| E2 | Decorin | 0.81 | 0.03 | ||||
| IFNα | 0.77 | 0.05 | |||||
| IL-10 | 0.81 | 0.03 | |||||
| IL-18 | 0.79 | 0.04 | |||||
| RANTES | 0.79 | 0.04 | |||||
| A4 | IL-1αA | 0.85 | 0.01 | ||||
| IL1-F5 | 0.79 | 0.04 | |||||
| RANTES | −0.84 | 0.02 | |||||
| TNFα | 0.83 | 0.02 | |||||
| IL-1β | 0.82 | 0.02 | |||||
| IL-13 | 0.85 | 0.01 | |||||
| IL-17αA | 0.82 | 0.01 | |||||
| IL-21 | 0.82 | 0.02 | 0.81 | 0.03 | |||
| MIG | 0.81 | 0.03 | 0.80 | 0.03 | |||
| P4 | CD40L | 0.83 | 0.02 | 0.82 | 0.02 | ||
| IL-13 | −0.81 | 0.03 | |||||
| IL17αA | −0.80 | 0.04 | |||||
| MIG | −0.82 | 0.03 | |||||
| Follicular Fluid | |||||
|---|---|---|---|---|---|
| DFA | FSHLow | ||||
| Cytokine | R2 | p-Value | R2 | p-Value | |
| E2 | MIG | −0.82 | 0.02 | ||
| P4 | ANGPT1 | 0.79 | 0.05 | ||
| RANTES | 0.84 | 0.02 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snider, A.P.; Gomes, R.S.; Summers, A.F.; Tenley, S.C.; Abedal-Majed, M.A.; McFee, R.M.; Wood, J.R.; Davis, J.S.; Cupp, A.S. Identification of Lipids and Cytokines in Plasma and Follicular Fluid before and after Follicle-Stimulating Hormone Stimulation as Potential Markers for Follicular Maturation in Cattle. Animals 2023, 13, 3289. https://doi.org/10.3390/ani13203289
Snider AP, Gomes RS, Summers AF, Tenley SC, Abedal-Majed MA, McFee RM, Wood JR, Davis JS, Cupp AS. Identification of Lipids and Cytokines in Plasma and Follicular Fluid before and after Follicle-Stimulating Hormone Stimulation as Potential Markers for Follicular Maturation in Cattle. Animals. 2023; 13(20):3289. https://doi.org/10.3390/ani13203289
Chicago/Turabian StyleSnider, Alexandria P., Renata S. Gomes, Adam F. Summers, Sarah C. Tenley, Mohamed A. Abedal-Majed, Renee M. McFee, Jennifer R. Wood, John S. Davis, and Andrea S. Cupp. 2023. "Identification of Lipids and Cytokines in Plasma and Follicular Fluid before and after Follicle-Stimulating Hormone Stimulation as Potential Markers for Follicular Maturation in Cattle" Animals 13, no. 20: 3289. https://doi.org/10.3390/ani13203289
APA StyleSnider, A. P., Gomes, R. S., Summers, A. F., Tenley, S. C., Abedal-Majed, M. A., McFee, R. M., Wood, J. R., Davis, J. S., & Cupp, A. S. (2023). Identification of Lipids and Cytokines in Plasma and Follicular Fluid before and after Follicle-Stimulating Hormone Stimulation as Potential Markers for Follicular Maturation in Cattle. Animals, 13(20), 3289. https://doi.org/10.3390/ani13203289