
Seasonal Prey Abundance and Food Plasticity of the Vulnerable Snow Leopard (Panthera uncia) in the Lapchi Valley, Nepal Himalayas

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
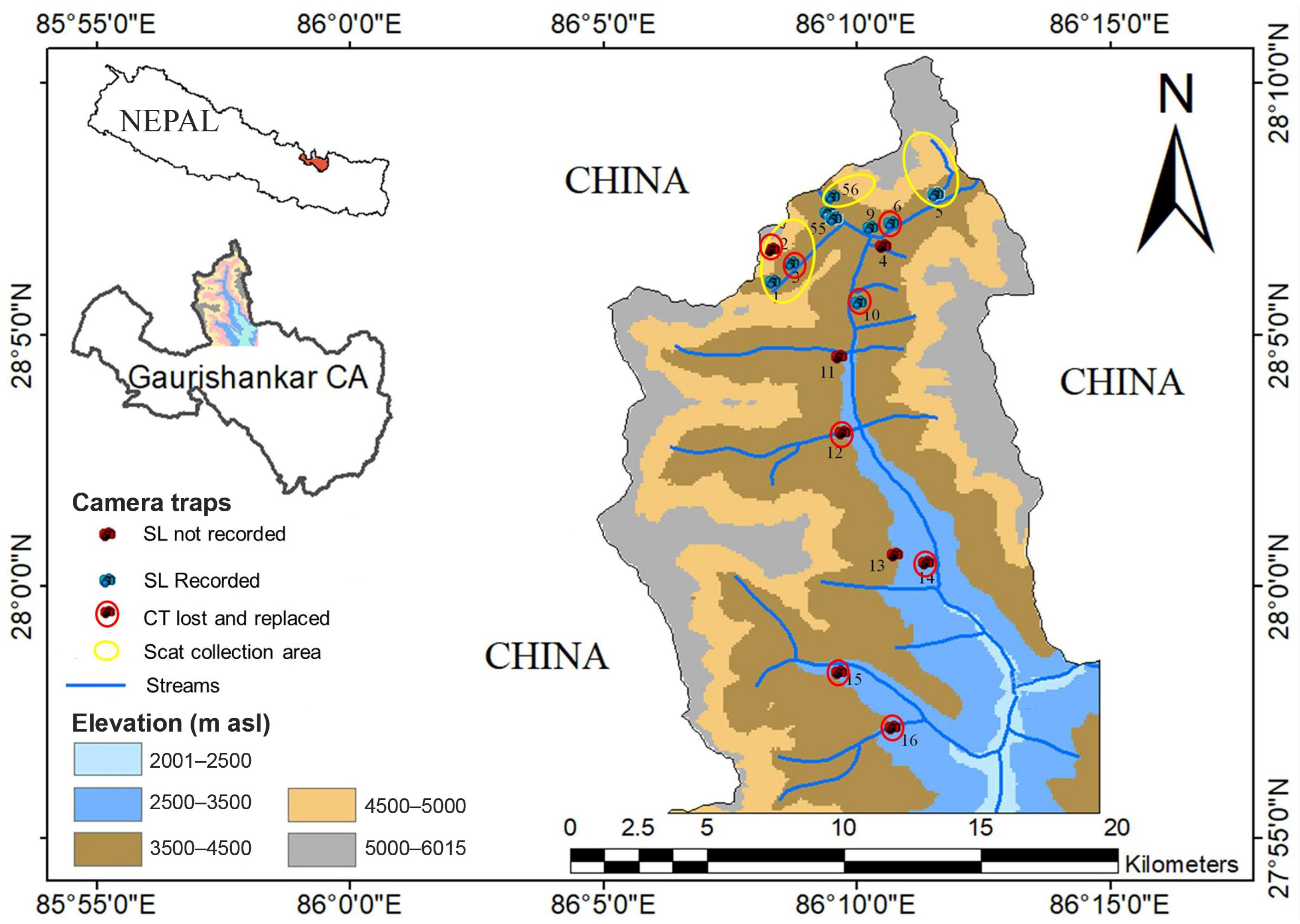
2.1. Study Area
2.2. Camera Trap Surveys
2.3. Scat Collection and Laboratory Analysis
2.4. Data Analysis
3. Results
3.1. Preys of Snow Leopard in Lapchi Valley
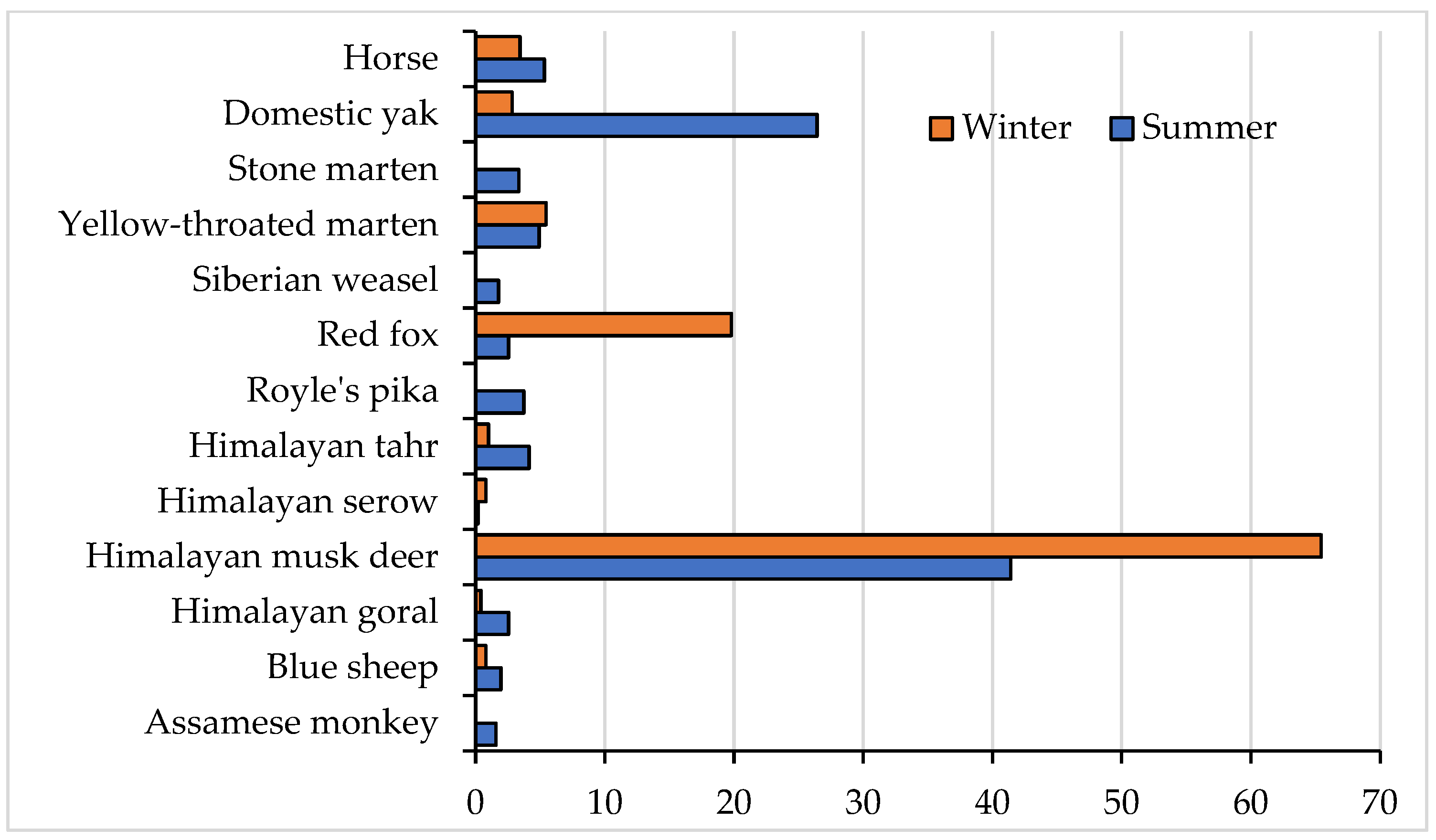
3.2. Seasonal Prey Diversity and Abundance in Lapchi Valley
3.3. Diet Composition of Snow Leopard
3.4. Biomass of Prey Consumption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCarthy, T.; Mallon, D.; Jackson, R.; Zahler, P.; McCarthy, K. Panthera uncia. The IUCN Red List of Threatened Species 2017:e.T22732A50664030. Available online: https://www.iucnredlist.org/species/22732/50664030 (accessed on 22 May 2023).
- Jackson, R. Snow leopards in Nepal. Oryx 1979, 15, 191–195. [Google Scholar] [CrossRef]
- McCarthy, T.M.; Chapron, G. Snow Leopard Survival Strategy; International Snow Leopard Trust and Snow Leopard Network: Seattle, WA, USA, 2003; Volume 105. [Google Scholar]
- WWF. Available online: http://www.wwfnepal.org/what_we_do/wildlife/snow_leopard/ (accessed on 21 May 2023).
- Green, M.J. Protected areas and snow leopards: Their distribution and status. In Proceedings of the Fifth International Snow Leopard Symposium, Srinagar, India, 13–15 October 1986; International Snow Leopard Trust and Wildlife Institute of India: Seattle, WA, USA, 1988; pp. 3–19. [Google Scholar]
- Jackson, R. Aboriginal hunting in West Nepal with reference to musk deer Moschus moschiferus moschiferus and snow leopard Panthera uncia. Biol. Conserv. 1979, 16, 63–72. [Google Scholar] [CrossRef]
- Kyes, R.C.; Chalise, M.K. Assessing the Status of the Snow Leopard Population in Langtang National Park, Nepal; Final Project Report Submitted to International Snow Leopard Trust: Seattle, WA, USA, 2005; 11p. [Google Scholar]
- Khanal, G.; Poudyal, L.P.; Devkota, B.P.; Ranabhat, R.; Wegge, P. Status and conservation of the snow leopard Panthera uncia in Api Nampa Conservation Area, Nepal. Oryx 2018, 54, 421–428. [Google Scholar] [CrossRef]
- WWF. Altai-Sayan Ecoregion Conservation Strategy; WWF: Gland, Switzerland, 2012. [Google Scholar]
- Koju, N.P.; Bashyal, B.; Pandey, B.P.; Shah, S.N.; Thami, S.; Bleisch, W.V. First camera-trap record of the snow leopard Panthera uncia in Gaurishankar Conservation Area, Nepal. Oryx 2021, 55, 173–176. [Google Scholar] [CrossRef]
- MoFSC. Snow Leopard and Ecosystem Management Plan (2017–2026); Ministry of Forest and Soil Conservation: Kathmandu, Nepal, 2017.
- Lahkar, D.; Ahmed, M.F.; Begum, R.H.; Das, S.K.; Lahkar, B.P.; Sarma, H.K.; Harihar, A. Camera-trapping survey to assess diversity, distribution and photographic capture rate of terrestrial mammals in the aftermath of the ethnopolitical conflict in Manas National Park, Assam, India. J. Threat. Taxa 2018, 10, 12008–12017. [Google Scholar] [CrossRef]
- Lovari, S.; Boesi, R.; Minder, I.; Mucci, N.; Randi, E.; Dematteis, A.; Ale, S. Restoring a keystone predator may endanger a prey species in a human-altered ecosystem: The return of the snow leopard to Sagarmatha National Park. Anim. Conserv. 2009, 12, 559–570. [Google Scholar] [CrossRef]
- Jackson, R.; Mishra, C.; McCarthy, T.; Ale, S. Snow leopards: Conflict and conservation. In Biology and Conservation of Wild Felids; Macdonald, D.W., Loveridge, A.J., Eds.; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Ahmad, S.; Nabi, G.; Hacker, C.E.; Strelnikov, I.I.; Luan, X. Increasing threats to snow leopard survival in Pakistan. Front. Ecol. Evol. 2022, 10, 818798. [Google Scholar] [CrossRef]
- Li, J.; Weckworth, B.V.; McCarthy, T.M.; Liang, X.; Liu, Y.; Xing, R.; Li, D.; Zhang, Y.; Xue, Y.; Jackson, R. Defining priorities for global snow leopard conservation landscapes. Biol. Conserv. 2020, 241, 108387. [Google Scholar] [CrossRef]
- Fox, J.L. A Review of the Status and Ecology of the Snow Leopard (Panthera uncia); International Snow Leopard Trust: Seattle, WA, USA, 1989. [Google Scholar]
- Oli, M.K.; Taylor, I.R.; Rogers, D.M. Diet of the snow leopard (Panthera uncia) in the Annapurna Conservation Area, Nepal. J. Zool. 1993, 231, 365–370. [Google Scholar] [CrossRef]
- Shehzad, W.; McCarthy, T.M.; Pompanon, F.; Purevjav, L.; Coissac, E.; Riaz, T.; Taberlet, P. Prey Preference of Snow Leopard (Panthera uncia) in South Gobi, Mongolia. PloS ONE 2012, 7, e32104. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Aihartza, J.; Kindlmann, P. Diet and prey selection by snow leopards in the Nepalese Himalayas. PLoS ONE 2018, 13, e0206310. [Google Scholar] [CrossRef] [PubMed]
- Chetri, M.; Odden, M.; Wegge, P. Snow leopard and Himalayan wolf: Food habits and prey selection in the Central Himalayas, Nepal. PLoS ONE 2017, 12, e0170549. [Google Scholar] [CrossRef]
- Sindorf, N. Eastern Nepal Landscape Water Resources and Climate Change Sensitivity Analysis: High Asia Mountain Initiative; WWF-US: Washington DC, USA, 2017. [Google Scholar]
- Koju, N.P.; Bashyal, B.; Pandey, B.P.; Thami, S.; Dhamala, M.K.; Shah, S.N. New record on Asiatic golden cat Catopuma temminckii Vigors & Horsfield, 1827 (Mammalia: Carnivora: Felidae): Photographic evidence of its westernmost distribution in Gaurishankar Conservation Area, Nepal. J. Threat. Taxa 2020, 12, 15256–15261. [Google Scholar]
- Pandey, B.P.; Thami, S.M.; Shrestha, R.; Chalise, M.K. On the occurrence of the Himalayan Wolf Canis lupus, L. 1758 (Mammalia: Carnivora: Canidae) in the Gaurishankar Conservation Area, Nepal; its existence confirmed through sign and visual evidence in Rolwaling Valley. J. Threat. Taxa 2021, 13, 18967–18974. [Google Scholar] [CrossRef]
- Chetri, M.; Regmi, P.R.; Dahal, T.P.; Thami, S. A checklist of mammals of Gaurishankar Conservation Area, Nepal. Nepal. J. Zool. 2022, 6, 56–62. [Google Scholar] [CrossRef]
- Jnawali, S.; Baral, H.; Lee, S.; Acharya, K.; Upadhyay, G.; Pandey, M.; Shrestha, R.; Joshi, D.; Lamichhane, B.; Griffiths, J. The Status of Nepal’s Mammals: The National Red List Series-IUCN; Department of National Parks and Wildlife Conservation: Kathmandu, Nepal, 2011; p. 276.
- Koju, N.P.; Bashyal, B.; Shah, S.N. Habitat preference of Himalayan musk deer (Moschus leucogaster Hodgson, 1839) at Lapchi of Bigu Rural Municipality, Gaurishankar Conservation Area. Nepal J. Environ. Sci. 2021, 9, 21–28. [Google Scholar] [CrossRef]
- Wangdi, T.; Tobgay, S.; Dorjee, K.; Dorji, K.; Wangyel, S. The distribution, status and conservation of the Himalayan Musk Deer Moschus chrysogaster in Sakteng Wildlife Sanctuary. Glob. Ecol. Conserv. 2019, 17, e00466. [Google Scholar] [CrossRef]
- Li, X.; Bleisch, W.V.; Jiang, X. Unveiling a wildlife haven: Occupancy and activity patterns of mammals at a Tibetan sacred mountain. Eur. J. Wildl. Res. 2018, 64, 53. [Google Scholar] [CrossRef]
- Rovero, F.; Tobler, M.; Sanderson, J. Camera trapping for inventorying terrestrial vertebrates. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories and Monitoring; Belgian Development Cooperation: Brussels, Belgium, 2010; pp. 100–128. [Google Scholar]
- Khatoon, R.; Hussain, I.; Anwar, M.; Nawaz, M.A. Diet selection of snow leopard (Panthera uncia) in Chitral, Pakistan. Turk. J. Zool. 2017, 41, 914–923. [Google Scholar] [CrossRef]
- Alexander, J.S.; Johansson, O.; Sharma, K.; Durbach, I.; Borchers, D. Snow Leopard Identification Good Practices. 2020. Available online: https://globalsnowleopard.org/wp-content/uploads/2020/10/Best-practices-for-Individual-ID.pdf (accessed on 5 July 2023).
- Oli, M.K. A key for the identification of the hair of mammals of a snow leopard (Panthera uncia) habitat in Nepal. J. Zool. 1993, 231, 71–93. [Google Scholar] [CrossRef]
- Mukherjee, S.; Goyal, S.; Chellam, R. Standardisation of scat analysis techniques for leopard (Panthera pardus) in Gir National Park, Western India. Mammalia 1994, 139–143. [Google Scholar] [CrossRef]
- Bahuguna, A. Species Identification from Guard Hair of Selected Indian Mammals; Wildlife Institute of India: Dehradun, India, 2010. [Google Scholar]
- Tremori, T.; Monteiro Garcia, F.; Montoya Flórez, L.M.; Picado Gonçalves, B.; Ferraz de Camargo, B.; Gwinnett, C.; Teixeira, C.; Sousa Rocha, N. Hair analysis of mammals of Brazilian wildlife for forensic purposes. Open J. Anim. Sci. 2018, 8, 335–345. [Google Scholar] [CrossRef]
- Kshirsagar, S.; Singh, B.; Fulari, S. Comparative study of human and animal hair in relation with diameter and medullary index. Indian J. Forensic Med. Pathol. 2009, 2, 105–108. [Google Scholar]
- Devkota, B.P.; Silwal, T.; Kolejka, J. Prey density and diet of snow leopard (Uncia uncia) in Shey Phoksundo National Park, Nepal. Appl. Ecol. Environ. Sci. 2013, 1, 55–60. [Google Scholar]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Carbone, C.; Christie, S.; Conforti, K.; Coulson, T.; Franklin, N.; Ginsberg, J.; Griffiths, M.; Holden, J.; Kawanishi, K.; Kinnaird, M. The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim. Conserv. Forum 2001, 4, 75–79. [Google Scholar] [CrossRef]
- Ackerman, B.B.; Lindzey, F.G.; Hemker, T.P. Cougar food habits in southern Utah. J. Wildl. Manag. 1984, 48, 147–155. [Google Scholar] [CrossRef]
- Harihar, A.; Pandav, B.; Goyal, S. Responses of leopard Panthera pardus to the recovery of a tiger Panthera tigris population. J. Appl. Ecol. 2011, 48, 806–814. [Google Scholar] [CrossRef]
- Palei, H.S.; Pradhan, T.; Sahu, H.K.; Nayak, A.K. Diet and activity pattern of leopard in relation to prey in tropical forest ecosystem. Mammalia 2022, 86, 1–12. [Google Scholar] [CrossRef]
- Shrestha, B.; Vařachová, S.; Kindlmann, P. A Key for Identifying the prey of snow leopard in Nepal using features of the structure of the hair of their prey present in their faeces. In Snow Leopards in Nepal: Predator-Prey System on the Top of the World; Springer: Berlin/Heidelberg, Germany, 2022; pp. 75–94. [Google Scholar]
- Amin, R.; Baral, H.S.; Lamichhane, B.R.; Poudyal, L.P.; Lee, S.; Jnawali, S.R.; Acharya, K.P.; Upadhyaya, G.P.; Pandey, M.B.; Shrestha, R. The status of Nepal’s mammals. J. Threat. Taxa 2018, 10, 11361–11378. [Google Scholar] [CrossRef]
- Karanth, K.U.; Sunquist, M.E. Prey selection by tiger, leopard and dhole in tropical forests. J. Anim. Ecol. 1995, 64, 439–450. [Google Scholar] [CrossRef]
- Pianka, E. The structure of lizard communities. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 53–74. [Google Scholar] [CrossRef]
- Gotelli, N.; Hart, E.; Ellison, A.; Hart, M.E. Package ‘EcoSimR’. 2015. Available online: https://zenodo.org/record/16522 (accessed on 21 July 2023). [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 21 July 2023).
- Wegge, P.; Shrestha, R.; Flagstad, Ø. Snow leopard Panthera uncia predation on livestock and wild prey in a mountain valley in northern Nepal: Implications for conservation management. Wildl. Biol. 2012, 18, 131–141. [Google Scholar] [CrossRef]
- Ale, S.B.; Shrestha, B.; Jackson, R. On the status of snow leopard Panthera uncia (Schreber, 1775) in Annapurna, Nepal. J. Threat. Taxa 2014, 6, 5534–5543. [Google Scholar] [CrossRef]
- Jackson, R.; Ahlborn, G. Preliminary habitat suitability model for the snow leopard Panthera uncia in west Nepal. In International Pedigree Book of Snow Leopards; Helsinki Zoo: Helsinki, Finland, 1984; Volume 4, pp. 43–52. [Google Scholar]
- Lyngdoh, S.; Shrotriya, S.; Goyal, S.P.; Clements, H.; Hayward, M.W.; Habib, B. Prey preferences of the snow leopard (Panthera uncia): Regional diet specificity holds global significance for conservation. PLoS ONE 2014, 9, e88349. [Google Scholar] [CrossRef]
- Hacker, C.E.; Jevit, M.; Hussain, S.; Muhammad, G.; Munkhtsog, B.; Munkhtsog, B.; Zhang, Y.; Li, D.; Liu, Y.; Farrington, J.D. Regional comparison of snow leopard (Panthera uncia) diet using DNA metabarcoding. Biodivers. Conserv. 2021, 30, 797–817. [Google Scholar] [CrossRef]
- Lu, Q.; Xiao, L.; Cheng, C.; Lu, Z.; Zhao, J.; Yao, M. Snow leopard dietary preferences and livestock predation revealed by fecal DNA Metabarcoding: No evidence for apparent competition between wild and domestic prey. Front. Ecol. Evol. 2021, 9, 783546. [Google Scholar] [CrossRef]
- Oli, M.K.; Taylor, I.R.; Rogers, M.E. Snow leopard Panthera uncia predation of livestock: An assessment of local perceptions in the Annapurna Conservation Area, Nepal. Biol. Conserv. 1994, 68, 63–68. [Google Scholar] [CrossRef]
- Mallon, D.; Harris, R.B.; Wegge, P. Snow leopard prey and diet. In Snow Leopards; Elsevier: Amsterdam, The Netherlands, 2016; pp. 43–55. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Wildlife Recorded in Cameras | Total Photos | CT Days to Respective CTs | No of Recorded CTs | PCRI Per Recorded CT Base |
|---|---|---|---|---|
| 8 | 333 | 1 | 2.4 |
| 14 | 590 | 3 | 2.37 |
| 15 | 333 | 1 | 4.5 |
| 534 | 2188 | 8 | 24.4 |
| 3 | 393 | 2 | 0.76 |
| 26 | 333 | 1 | 7.8 |
| 19 | 382 | 2 | 4.97 |
| 53 | 1780 | 7 | 2.97 |
| 29 | 1271 | 5 | 2.28 |
| 46 | 1272 | 5 | 3.61 |
| 57 | 1628 | 7 | 3.5 |
| 101 | 2690 | 11 | 3.75 |
| 9 | 901 | 3 | 0.99 |
| 37 | 2063 | 8 | 1.79 |
| 51 | 1930 | 7 | 2.64 |
| 17 | 394 | 2 | 4.31 |
| 1 | 334 | 1 | 0.29 |
| 148 | 2451 | 9 | 6.03 |
| 44 | 1621 | 5 | 2.71 |
| Total events | 1212 | 82.16 | ||
| CT days | 3023 |
| SN | Prey Type | Summer | Winter | χ2 | df | p |
|---|---|---|---|---|---|---|
| Livestock | 31.75 | 6.26 | 4.27 | 1 | 0.03 | |
| All wild ungulates | 50.29 | 68.48 | 7.12 | 4 | 0.12 | |
| Wild ungulates, consumed (present in scat) | 47.53 | 67.27 | 4.53 | 2 | 0.001 | |
| Wild ungulate, not consumed | 2.76 | 1.21 | 1.57 | 1 | 0.208 | |
| Meso-mammals, all | 9.07 | 25.25 | 6.96 | 2 | 0.03 | |
| Meso-mammals, not consumed | 9.86 | 5.45 | 4.02 | 2 | 0.133 | |
| Small mammals | 8.87 | 0 | NA | NA | NA | |
| Wild prey consumed only * | 88.08 | 53.84 | 15.28 | 4 | 0.0041 |
| Prey Species Common Name | Winter | Summer | ||
|---|---|---|---|---|
| Number of Occurrences | % of Prey Species Contribution | Number of Occurrences | % of Prey Species Contribution | |
| 3 | 1.5 | 2 | 1 |
| 12 | 6 | 5 | 2.5 |
| 0 | 0 | 2 | 1 |
| 9 | 4.5 | 3 | 1.5 |
| 41 | 20.5 | 11 | 5.5 |
| 6 | 3 | 2 | 1 |
| 9 | 4.5 | 2 | 1 |
| 88 | 44 | 31 | 15.5 |
| 13 | 6.5 | 23 | 11.5 |
| 0 | 0 | 65 | 32.5 |
| 18 | 9 | 51 | 25.5 |
| 1 | 0.5 | 0 | 0 |
| 0 | 0 | 3 | 1.5 |
| Prey Type | Winter | Summer | χ2 | df | p |
|---|---|---|---|---|---|
| Livestock | 31 | 139 | 25.06 | 2 | 0.0001 |
| Wild ungulates | 135 | 44 | 0.46 | 2 | 0.791 |
| Meso-mammals | 12 | 6 | 5.23 | 2 | 0.0001 |
| Small mammals | 21 | 8 | 0.06 | 1 | 0.793 |
| Prey | Average Weight | No of Scat with Respective Prey | Biomass Per scat | Biomass Consumed | Estimated Percentage of Biomass Consumed (E) | ||
|---|---|---|---|---|---|---|---|
| A | C | B | D | Summer | Winter | Total | |
| 0.50 | 3.00 | 2.00 | 5.99 | 2.70 | 6.34 | 4.37 |
| 0.20 | 6.00 | 1.99 | 11.92 | 5.37 | 12.61 | 8.70 |
| 6.00 | 1.00 | 2.19 | 2.19 | 2.96 | 0.00 | 1.60 |
| 0.18 | 5.00 | 1.99 | 9.93 | 2.68 | 12.61 | 7.24 |
| 12.00 | 5.00 | 2.40 | 12.00 | 6.48 | 11.42 | 8.75 |
| 36.00 | 2.00 | 3.24 | 6.48 | 4.37 | 5.14 | 4.73 |
| 4.50 | 4.00 | 2.14 | 8.55 | 2.89 | 10.17 | 6.24 |
| 30.00 | 7.00 | 3.03 | 21.21 | 8.18 | 24.04 | 15.47 |
| 80.00 | 4.00 | 4.78 | 19.12 | 19.36 | 7.58 | 13.95 |
| 25.00 | 5.00 | 2.86 | 14.28 | 19.27 | 0.00 | 10.41 |
| 125.00 | 4.00 | 6.36 | 25.42 | 25.74 | 10.08 | 18.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koju, N.P.; Gosai, K.R.; Bashyal, B.; Byanju, R.; Shrestha, A.; Buzzard, P.; Beisch, W.B.; Khanal, L. Seasonal Prey Abundance and Food Plasticity of the Vulnerable Snow Leopard (Panthera uncia) in the Lapchi Valley, Nepal Himalayas. Animals 2023, 13, 3182. https://doi.org/10.3390/ani13203182
Koju NP, Gosai KR, Bashyal B, Byanju R, Shrestha A, Buzzard P, Beisch WB, Khanal L. Seasonal Prey Abundance and Food Plasticity of the Vulnerable Snow Leopard (Panthera uncia) in the Lapchi Valley, Nepal Himalayas. Animals. 2023; 13(20):3182. https://doi.org/10.3390/ani13203182
Chicago/Turabian StyleKoju, Narayan Prasad, Kamal Raj Gosai, Bijay Bashyal, Reena Byanju, Arati Shrestha, Paul Buzzard, Willian Bill Beisch, and Laxman Khanal. 2023. "Seasonal Prey Abundance and Food Plasticity of the Vulnerable Snow Leopard (Panthera uncia) in the Lapchi Valley, Nepal Himalayas" Animals 13, no. 20: 3182. https://doi.org/10.3390/ani13203182
APA StyleKoju, N. P., Gosai, K. R., Bashyal, B., Byanju, R., Shrestha, A., Buzzard, P., Beisch, W. B., & Khanal, L. (2023). Seasonal Prey Abundance and Food Plasticity of the Vulnerable Snow Leopard (Panthera uncia) in the Lapchi Valley, Nepal Himalayas. Animals, 13(20), 3182. https://doi.org/10.3390/ani13203182