

Effects of Glyceryl Monolaurate on Production Performance, Egg Quality, Oviduct Cytokines and Intestinal Microflora of 66 Weeks Old Laying Hens
 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Treatment
2.2. Sample Collection
2.3. Production Performance
2.4. Egg Quality
2.5. Inflammatory Cytokines
2.6. Histological Morphology
2.7. Real-Time Quantitative PCR
2.8. Volatile Fatty Acid Analysis
2.9. Intestinal Microbiota Community
2.10. Statistical Analysis
3. Results
3.1. Production Performance
3.2. Ileum Morphology
3.3. Tissue Inflammatory Factors
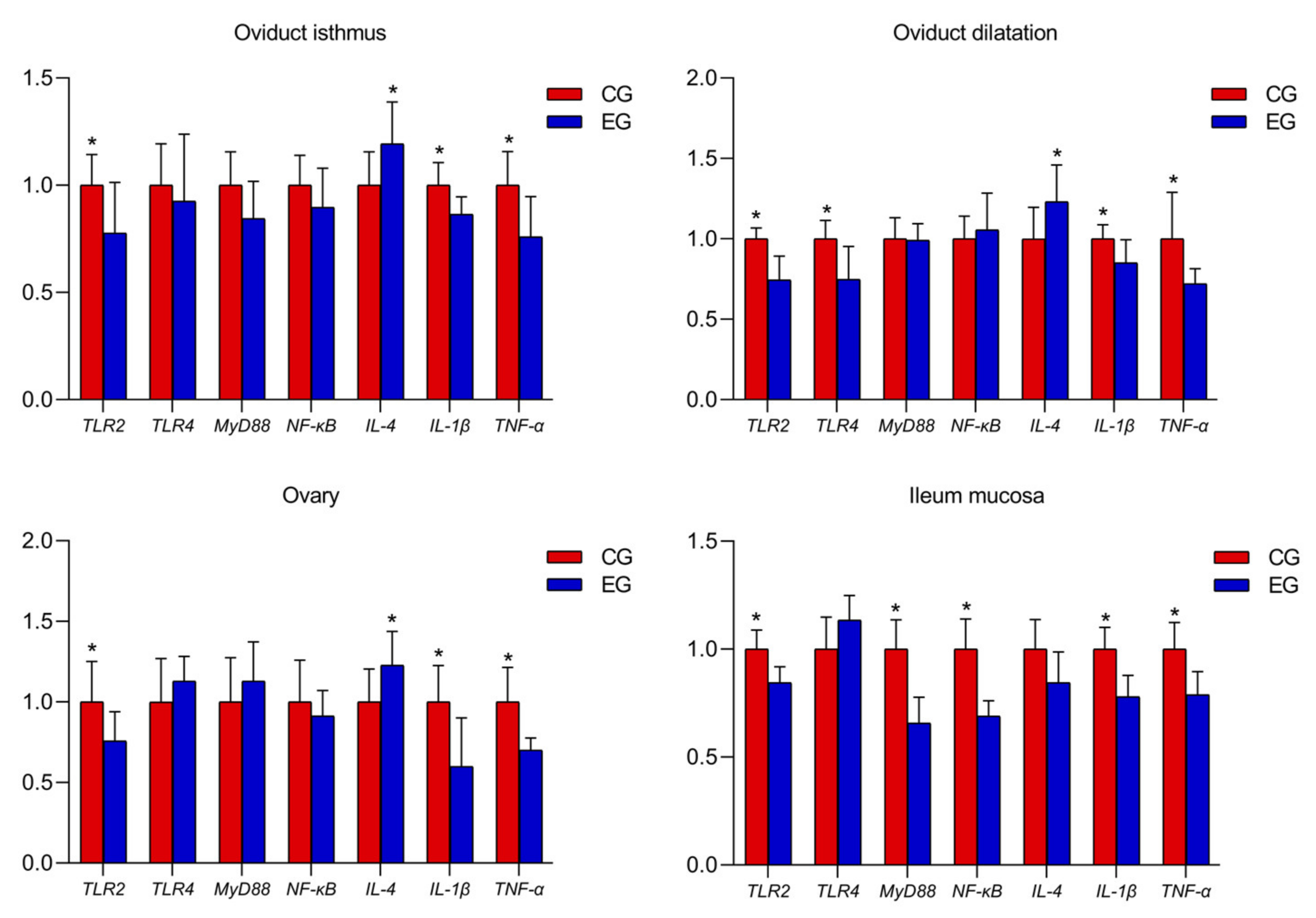
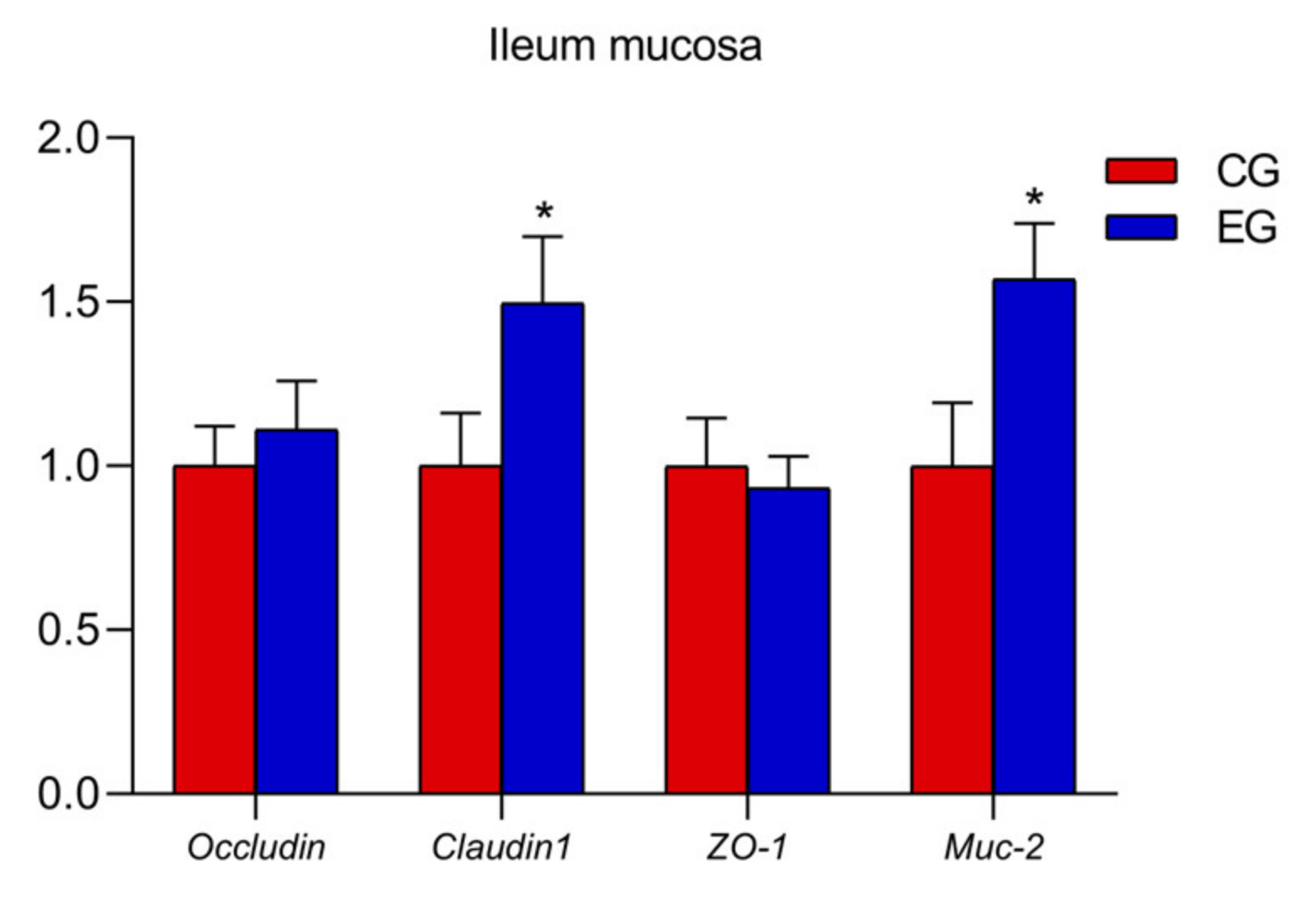
3.4. Relative mRNA Expression of Oviduct, Ovary and Ileum Mucosa Genes
3.5. Volatile Fatty Acid Levels in Cecal Contents
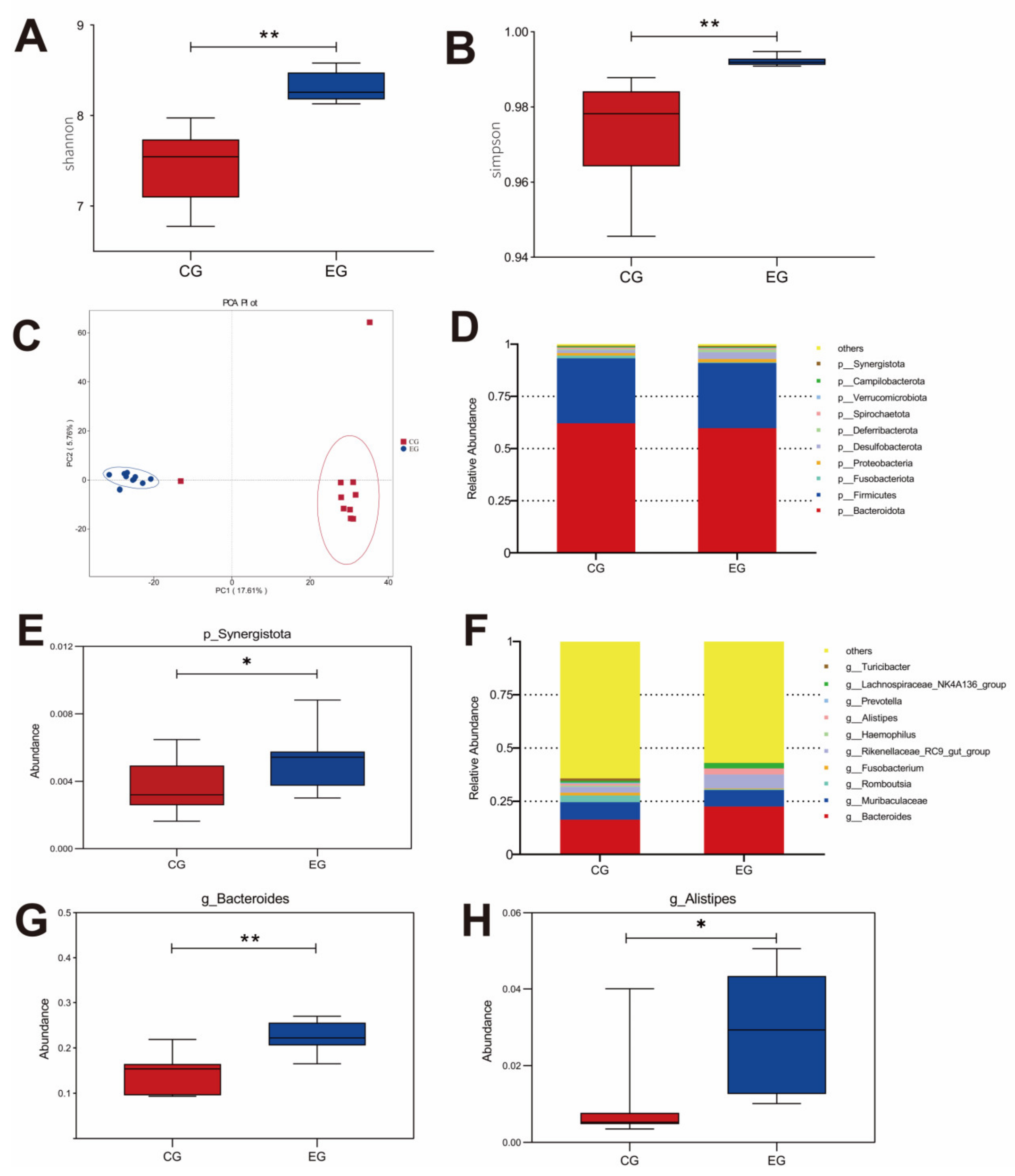
3.6. Cecal Microbial Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dai, H.; Lv, Z.; Huang, Z.; Ye, N.; Li, S.; Jiang, J.; Cheng, Y.; Shi, F. Dietary hawthorn-leaves flavonoids improves ovarian function and liver lipid metabolism in aged breeder hens. Poult. Sci. 2021, 100, 101499. [Google Scholar] [CrossRef]
- Guo, S.; Niu, J.; Xv, J.; Fang, B.; Zhang, Z.; Wang, L.; Ding, B. Interactive effects of vitamins A and K3 on laying performance, egg quality, tibia attributes and antioxidative status of aged Roman Pink laying hens. Animal 2021, 15, 100242. [Google Scholar] [CrossRef] [PubMed]
- Sirri, F.; Zampiga, M.; Berardinelli, A. Effects of genotype and age on eggshell cuticle coverage and color profile in modern laying hen strains. Poult. Sci. 2021, 101, 101691. [Google Scholar] [CrossRef] [PubMed]
- Nannoni, E.; Buonaiuto, G.; Martelli, G.; Lizzi, G.; Trevisani, G.; Garavini, G.; Sardi, L. Influence of Increased Freedom of Movement on Welfare and Egg Laying Pattern of Hens Kept in Aviaries. Animals 2022, 12, 2307. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zheng, J.; Duan, Z.; Yang, N.; Xu, G. The influences of SE infection on layers’ production performance, egg quality and blood biochemical indicators. J. Anim. Sci. Biotechnol. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Shokrollahi, B.; Yavari, Z.; Kordestani, A. Effects of dietary medium-chain fatty acids on performance, carcass characteristics, and some serum parameters of broiler chickens. Br. Poult. Sci. 2014, 55, 662–667. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhao, M.; Zhang, H.; Li, Y.; Liu, M.; Feng, F. Antimicrobial Emulsifier-Glycerol Monolaurate Induces Metabolic Syndrome, Gut Microbiota Dysbiosis, and Systemic Low-Grade Inflammation in Low-Fat Diet Fed Mice. Mol. Nutr. Food Res. 2018, 62, 1700547. [Google Scholar] [CrossRef]
- Marta, A.P.L.; Melissa, E.D.A.; Michelle, J.C.R.; Geraldo, L.S. Enzymatic synthesis of medium chain monoglycerides in a solvent-free system. Appl. Biochem. Biotechnol. 2002, 98–100, 987–996. [Google Scholar]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African swine fever virus in liquid and feed by medium-chain fatty acids and glycerol monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Fortuoso, B.F.; Dos Reis, J.H.; Gebert, R.R.; Barreta, M.; Griss, L.G.; Casagrande, R.A.; De, C.T.; Santiani, F.; Campigotto, G.; Rampazzo, L.; et al. Glycerol monolaurate in the diet of broiler chickens replacing conventional antimicrobials: Impact on health, performance and meat quality. Microb. Pathog. 2019, 129, 161–167. [Google Scholar] [CrossRef]
- Lan, J.; Chen, G.; Cao, G.; Tang, J.; Li, Q.; Zhang, B.; Yang, C. Effects of α-glyceryl monolaurate on growth, immune function, volatile fatty acids, and gut microbiota in broiler chickens. Poult. Sci. 2020, 100, 100875. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.T.; Loh, T.C.; Foo, H.L.; Hair-Bejo, M.; Azhar, B.K. Effects of feeding metabolite combinations produced by Lactobacillus plantarum on growth performance, faecal microbial population, small intestine villus height and faecal volatile fatty acids in broilers. Br. Poult. Sci. 2009, 50, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhang, L.; Cao, G.; Feng, J.; Yue, M.; Xu, Y.; Dai, B.; Han, Q.; Guo, X. Effects of dietary supplementation with essential oils and organic acids on the growth performance, immune system, fecal volatile fatty acids, and microflora community in weaned piglets. J. Anim. Sci. 2019, 97, 133–143. [Google Scholar] [CrossRef]
- Liu, M.; Wang, J.; Feng, F. Effects of glycerol monolaurate on production performance, egg quality, serum parameters, immune organ index and abdominal fat morphology of layers. China Poult. 2017, 39, 24–30. [Google Scholar]
- Shimaa, A.A.; Afaf, A.N.; Hanan, S.A.-K.; Dina, M.M.A.; Doaa, M.A.; Elshimaa, M.R.; Wafaa, R.I.A.S.; Mohamed, F.M.F.; Dalia, E.A.; Ahmed, A.A. A-W.; et al. Effect of Dietary Medium-Chain α-Monoglycerides on the Growth Performance, Intestinal Histomorphology, Amino Acid Digestibility, and Broiler Chickens’ Blood Biochemical Parameters. Animals 2020, 11, 57. [Google Scholar]
- Mo, Q.; Liu, T.; Fu, A.; Ruan, S.; Zhong, H.; Tang, J.; Zhao, M.; Li, Y.; Zhu, S.; Cai, H.; et al. Novel Gut Microbiota Patterns Involved in the Attenuation of Dextran Sodium Sulfate-Induced Mouse Colitis Mediated by Glycerol Monolaurate via Inducing Anti-inflammatory Responses. mBio 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Zhang, Y.; Li, X.; Jiang, X.; Ding, H. Effects of Dietary Supplementation with Glycerol Monolaurate (GML) or the Combination of GML and Tributyrin on Growth Performance and Rumen Microbiome of Weaned Lambs. Animals 2022, 12, 1309. [Google Scholar] [CrossRef]
- Liu, T.; Li, C.; Li, Y.; Feng, F. Glycerol Monolaurate Enhances Reproductive Performance, Egg Quality and Albumen Amino Acids Composition in Aged Hens with Gut Microbiota Alternation. Agriculture 2020, 10, 250. [Google Scholar] [CrossRef]
- Michaela, E.; Ivana, B.; Milos, S. Carotenoids and egg quality. Acta Fytotech. Et Zootech. 2014, 17, 55–57. [Google Scholar]
- Zhao, M.; Cai, H.; Liu, M.; Deng, L.; Li, Y.; Zhang, H.; Feng, F. Dietary glycerol monolaurate supplementation for the modification of functional properties of egg white protein. J. Sci. Food Agric. 2019, 99, 3852–3859. [Google Scholar] [CrossRef]
- Wen, C.; Xu, X.; Zhou, D.; Yu, Q.; Wang, T.; Zhou, Y. The effects of canthaxanthin microencapsulation on yolk color and canthaxanthin deposition in egg yolk of laying hens. Poult. Sci. 2022, 101, 101889. [Google Scholar] [CrossRef] [PubMed]
- Citi, S. Intestinal barriers protect against disease. Science 2018, 359, 1097–1098. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Chen, Z. The protective effect of γ-aminobutyric acid on the development of immune function in chickens under heat stress. J. Animal Physiol. Animal Nutrition 2015, 100, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, L.; Lv, Z.; Jin, S.; Wei, X.; Shi, F. Dietary Stevioside Supplementation Alleviates Lipopolysaccharide-Induced Intestinal Mucosal Damage through Anti-Inflammatory and Antioxidant Effects in Broiler Chickens. Antioxidants 2019, 8, 575. [Google Scholar] [CrossRef]
- Amer, S.A.; Tolba, S.A.; Alsadek, D.M.M.; Fattah, D.M.A.; Hassan, A.M.; Metwally, A.E. Effect of supplemental glycerol monolaurate and oregano essential oil blend on the growth performance, intestinal morphology, and amino acid digestibility of broiler chickens. BMC Vet. Res. 2021, 17, 1–12. [Google Scholar]
- Isaacs, C.E.; Litov, R.E.; Thormar, H. Antimicrobial activity of lipids added to human milk, infant formula, and bovine milk. J. Nutr. Biochem. 1995, 6, 362–366. [Google Scholar] [CrossRef]
- Papadopoulos, G.A.; Poutahidis, T.; Chalvatzi, S.; Kroustallas, F.; Karavanis, E.; Fortomaris, P. Effects of a tributyrin and monolaurin blend compared to high ZnO levels on growth performance, faecal microbial counts, intestinal histomorphometry and immunohistochemistry in weaned piglets: A field study in two pig herds. Res. Vet. Sci. 2022, 144, 54–65. [Google Scholar] [CrossRef]
- Jiang, Z.; Applegate, T.J.; Lossie, A.C. Cloning, annotation and developmental expression of the chicken intestinal MUC2 gene. PLoS ONE 2013, 8, 53781. [Google Scholar] [CrossRef]
- Heiskala, M.; Peterson, P.A.; Yang, Y. The roles of claudin superfamily proteins in paracellular transport. Traffic 2001, 2, 93–98. [Google Scholar] [CrossRef]
- Phua, D.C.Y.; Xu, J.; Ali, S.M.; Boey, A.; Gounko, N.V.; Hunziker, W. ZO-1 and ZO-2 are required for extra-embryonic endoderm integrity, primitive ectoderm survival and normal cavitation in embryoid bodies derived from mouse embryonic stem cells. PLoS ONE 2014, 9, 99532. [Google Scholar] [CrossRef]
- Mandon, M.; Cyr, D.G. Tricellulin and Its Role in the Epididymal Epithelium of the Rat1. Biol. Reprod. 2015, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mo, Q.; Fu, A.; Deng, L.; Zhao, M.; Li, Y.; Zhang, H.; Feng, F. High-dose Glycerol Monolaurate Up-Regulated Beneficial Indigenous Microbiota without Inducing Metabolic Dysfunction and Systemic Inflammation: New Insights into Its Antimicrobial Potential. Nutrients 2019, 11, 1981. [Google Scholar] [CrossRef] [PubMed]
- Michael, S.Z.; Aline, S.; Jon, C.D.H. Glycerol Monolaurate (GML) inhibits human T cell signaling and function by disrupting lipid dynamics. Sci. Rep. 2016, 6, 1–13. [Google Scholar]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Mo, Q.; Deng, L.; Zhong, H.; Liu, T.; Zhang, H.; Kang, J.; et al. Modulation of the Gut Microbiota during High-Dose Glycerol Monolaurate-Mediated Amelioration of Obesity in Mice Fed a High-Fat Diet. mBio 2020, 11, 20. [Google Scholar] [CrossRef]
- Matthew, A.O.; Jerrold, R.T. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2016, 14, 9–21. [Google Scholar]
- Yue, S.; Alemu, R.; Ji Hyuk, K.; Woo Kyun, K. The effect of dietary fructooligosaccharide supplementation on growth performance, intestinal morphology, and immune responses in broiler chickens challenged with Salmonella Enteritidis lipopolysaccharides. Poult. Sci. 2015, 94, 2887–2897. [Google Scholar]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol monolaurate ameliorated intestinal barrier and immunity in broilers by regulating intestinal inflammation, antioxidant balance, and intestinal microbiota. Biorxiv Zool. 2021, 12, 713485. [Google Scholar] [CrossRef]
- Marnie, L.P.; Patrick, M.S. Glycerol monolaurate inhibits the effects of Gram-positive select agents on eukaryotic cells. Biochemistry 2006, 45, 2387–2397. [Google Scholar]
- Sivinski, S.E.; Mamedova, L.K.; Rusk, R.A.; Elrod, C.C.; Swartz, T.H.; Mcgill, J.M.; Bradford, B.J. Development of an in vitro macrophage screening system on the immunomodulating effects of feed components. J. Anim. Sci. Biotechnol. 2020, 11, 89. [Google Scholar] [CrossRef]
- Kany, S.; Vollrathjan, T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Wang, W.; Li, Z.; Han, Q.; Guo, Y.; Zhang, B.; Dinca, R. Dietary live yeast and mannan-oligosaccharide supplementation attenuate intestinal inflammation and barrier dysfunction induced by Escherichia coli in broilers. Br. J. Nutr. 2016, 116, 1878–1888. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.H.; Hayley, E.G.; Jagadish, P.; Chris, S.M.; Scott, A.D.; Max, S. The efficacy of a cultured Synergistes jonesii inoculum to control hydroxypyridone toxicity in Bos indicus steers fed leucaena/grass diets. Anim. Prod. Sci. 2019, 59, 696–708. [Google Scholar]
- Lora, V.H.; Tore, M.; Jeffrey, I.G. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar]
- Zhu, C.; Huang, K.; Bai, Y.; Feng, X.; Gong, L.; Wei, C.; Huang, H.; Zhang, H. Dietary supplementation with berberine improves growth performance and modulates the composition and function of cecal microbiota in yellow-feathered broilers. Poult. Sci. 2020, 100, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Sam, Q.H.; Ling, H.; Yew, W.S.; Tan, Z.; Ravikumar, S.; Chang, M.; Chai, L. The Divergent Immunomodulatory Effects of Short Chain Fatty Acids and Medium Chain Fatty Acids. Int. J. Mol. Sci. 2021, 22, 6453. [Google Scholar] [CrossRef]
- Bianca, J.P.; Pamela, A.W.; Alida, C.M.V.; Alex, R.-P. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content, % | Nutrient Level 2 | Content |
|---|---|---|---|
| Corn | 63.50 | ME, MJ/kg 2 | 11.41 |
| Soybean meal | 22.00 | CP, % 3 | 15.91 |
| Soybean oil | 0.30 | Ca, % 3 | 4.06 |
| Bran | 5.00 | TP, % 3 | 0.48 |
| Limestone | 7.90 | Met, % 3 | 0.32 |
| NaCl | 0.30 | Lys, % 3 | 0.60 |
| Premixe 1 | 1.00 | Thr, % 3 | 0.64 |
| Total | 100.00 |
| Name | Position | Primer Sequence, 5′→3′ | Product Size (bp) | Access No. |
|---|---|---|---|---|
| β-Actin | Forward | TGCTGTGTTCCCATCTATCG | 150 | NM_205518 |
| Reverse | TTGGTGACAATACCGTGTTCA | |||
| TLR2 | Forward | CATTCACCATGAGGCAGGGATAG | 157 | NM_001396826.1 |
| Reverse | GGTGCAGATCAAGGACACTAGGA | |||
| TLR4 | Forward | TGACCTACCCATCGGACACT | 171 | NM_001030693.2 |
| Reverse | CTCAGGGCATCAAGGTCTCC | |||
| MyD88 | Forward | GATGATCCGTATGGGCATGGA | 170 | NM_001030962.4 |
| Reverse | ATGGACCACACACACGTTCC | |||
| NF-κB | Forward | TGCCTTTTGCTTGAGGGTGATG | 100 | XM_015285418.2 |
| Reverse | CTGCCAGTTTTGTGAAGCCC | |||
| IL-1β | Forward | CGACATCAACCAGAAGTGCTT | 298 | XM_015297469.1 |
| Reverse | GTCCAGGCGGTAGAAGATGA | |||
| IL-4 | Forward | AGCACTGCCACAAGAACCT | 160 | NM_001398460.1 |
| Reverse | GCTAGTTGGTGGAAGAAGGTAC | |||
| TNF-α | Forward | CCGTAGTGCTGTTCTATGACCG | 125 | NM_204267.2 |
| Reverse | GTTCCACATCTTTCAGAGCATCAA | |||
| Muc-2 | Forward | ATTGTGGTAACACCAACATTCATC | 135 | XM_0012334581.3 |
| Reverse | CTTTATAATGTCAGCACCAACTTCTC | |||
| ZO-1 | Forward | GGATGTTTATTTGGGCGGCT | 153 | XM_015278980.2 |
| Reverse | CCATTGTTGCACTCTTGCCG | |||
| Claudin-1 | Forward | CACACCCGTTAACACCAGATTT | 159 | NM_001013611.2 |
| Reverse | GAGGGGGCATTTTTGGGGTA | |||
| Occludin | Forward | AGCCCTCAATACCAGGATGTG | 125 | NM_205128.1 |
| Reverse | CGCTTGATGTGGAAGAGCTTG |
| Items | Control Group | Glyceryl Monolaurate Group | p-Value |
|---|---|---|---|
| Average egg weight, g | 58.66 ± 0.75 | 60.97 ± 0.54 | 0.024 |
| Laying rate, % | 83.38 ± 0.59 | 85.40 ± 0.51 | 0.019 |
| Feed intake, g | 129.27 ± 1.79 | 130.44 ± 1.20 | 0.595 |
| Feed conversion ratio, g/g | 2.20 ± 0.02 | 2.14 ± 0.02 | 0.038 |
| Defective eggs, % | 8.75 ± 0.17 | 8.02 ± 0.19 | 0.011 |
| Broken eggs, % | 0.78 ± 0.04 | 0.83 ± 0.03 | 0.358 |
| Pimpled eggs, % | 3.27 ± 0.05 | 3.12 ± 0.09 | 0.161 |
| Items | Control Group | Glyceryl Monolaurate Group | p-Value |
|---|---|---|---|
| Shell strength, kg·f/m2 | 5.27 ± 0.25 | 5.39 ± 0.15 | 0.676 |
| Shell thickness, mm | 0.38 ± 0.01 | 0.39 ± 0.01 | 0.365 |
| Albumen height, mm | 6.09 ± 0.04 | 6.28 ± 0.07 | 0.040 |
| Yolk color | 9.70 ± 0.15 | 10.30 ± 0.21 | 0.036 |
| Haugh unit | 78.55 ± 0.54 | 80.32 ± 0.57 | 0.037 |
| Items | Control Group | Glyceryl Monolaurate Group | p-Value |
|---|---|---|---|
| Villus height, μm | 627.79 ± 21.49 | 703.82 ± 24.33 | 0.031 |
| Crypt depth, μm | 144.91 ± 8.60 | 147.47 ± 7.55 | 0.826 |
| VCR, μm/μm | 4.39 ± 0.12 | 4.81 ± 0.12 | 0.023 |
| Items | Control Group | Glyceryl Monolaurate Group | p-Value |
|---|---|---|---|
| Oviduct isthmus | |||
| IL-2, pg/mg protein | 5.65 ± 0.30 | 5.17 ± 0.35 | 0.016 |
| IL-4, pg/mg protein | 140.31 ± 13.64 | 155.80 ± 16.78 | 0.083 |
| IL-6, pg/mg protein | 255.51 ± 15.52 | 237.13 ± 20.98 | 0.087 |
| IL-10, ng/g protein | 25.91 ± 1.52 | 28.99 ± 2.69 | 0.022 |
| IL-1β, pg/mg protein | 94.72 ± 3.78 | 88.12 ± 5.12 | 0.018 |
| TNF-α, ng/g protein | 34.15 ± 3.23 | 30.77 ± 2.01 | 0.036 |
| IFN-γ, pg/mg protein | 377.73 ± 11.66 | 347.53 ± 20.17 | 0.007 |
| Oviduct dilatation | |||
| IL-2, pg/mg protein | 5.89 ± 0.37 | 5.33 ± 0.35 | 0.012 |
| IL-4, pg/mg protein | 131.31 ± 7.41 | 147.61 ± 10.77 | 0.006 |
| IL-6, pg/mg protein | 265.78 ± 18.43 | 251.45 ± 25.48 | 0.251 |
| IL-10, ng/g protein | 25.01 ± 2.43 | 28.27 ± 2.43 | 0.027 |
| IL-1β, pg/mg protein | 100.08 ± 1.78 | 94.25 ± 5.37 | 0.028 |
| TNF-α, ng/g protein | 37.82 ± 2.99 | 34.44 ± 2.40 | 0.039 |
| IFN-γ, pg/mg protein | 385.31 ± 11.67 | 361.85 ± 22.09 | 0.034 |
| Ovary | |||
| IL-2, pg/mg protein | 4.15 ± 0.12 | 3.65 ± 0.44 | 0.023 |
| IL-4, pg/mg protein | 171.81 ± 11.81 | 187.04 ± 11.24 | 0.029 |
| IL-6, pg/mg protein | 213.61 ± 13.72 | 191.09 ± 9.58 | 0.004 |
| IL-10, ng/g protein | 35.46 ± 1.55 | 37.80 ± 1.28 | 0.010 |
| IL-1β, pg/mg protein | 67.87 ± 3.57 | 63.33 ± 3.94 | 0.043 |
| TNF-α, ng/g protein | 26.58 ± 1.52 | 24.40 ± 2.01 | 0.041 |
| IFN-γ, pg/mg protein | 323.79 ± 12.06 | 308.60 ± 10.21 | 0.026 |
| Items | Control Group | Glyceryl Monolaurate Group | p-Value |
|---|---|---|---|
| Acetic acid, μmol/g | 47.70 ± 10.44 | 59.08 ± 10.48 | 0.026 |
| Propionic acid, μmol/g | 12.75 ± 3.07 | 15.75 ± 2.97 | 0.031 |
| Isobutyric acid, μmol/g | 1.51 ± 0.28 | 1.45 ± 0.47 | 0.691 |
| Butyric acid, μmol/g | 5.65 ± 1.82 | 6.04 ± 2.28 | 0.678 |
| Isovaleric acid, μmol/g | 1.18 ± 0.27 | 1.23 ± 0.46 | 0.795 |
| Valeric acid, μmol/g | 1.22 ± 0.29 | 1.16 ± 0.23 | 0.596 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Zhang, R.; Dai, B.; Fu, C.; Zhao, G.; Xu, Y.; Yang, C. Effects of Glyceryl Monolaurate on Production Performance, Egg Quality, Oviduct Cytokines and Intestinal Microflora of 66 Weeks Old Laying Hens. Animals 2023, 13, 215. https://doi.org/10.3390/ani13020215
Cui Z, Zhang R, Dai B, Fu C, Zhao G, Xu Y, Yang C. Effects of Glyceryl Monolaurate on Production Performance, Egg Quality, Oviduct Cytokines and Intestinal Microflora of 66 Weeks Old Laying Hens. Animals. 2023; 13(2):215. https://doi.org/10.3390/ani13020215
Chicago/Turabian StyleCui, Zhenchuan, Ruiqiang Zhang, Bing Dai, Chunsheng Fu, Guiling Zhao, Yinglei Xu, and Caimei Yang. 2023. "Effects of Glyceryl Monolaurate on Production Performance, Egg Quality, Oviduct Cytokines and Intestinal Microflora of 66 Weeks Old Laying Hens" Animals 13, no. 2: 215. https://doi.org/10.3390/ani13020215
APA StyleCui, Z., Zhang, R., Dai, B., Fu, C., Zhao, G., Xu, Y., & Yang, C. (2023). Effects of Glyceryl Monolaurate on Production Performance, Egg Quality, Oviduct Cytokines and Intestinal Microflora of 66 Weeks Old Laying Hens. Animals, 13(2), 215. https://doi.org/10.3390/ani13020215