
The Effect of Prebiotic Supplements on the Gastrointestinal Microbiota and Associated Health Parameters in Pigs


Abstract
:Simple Summary
Abstract
1. Introduction
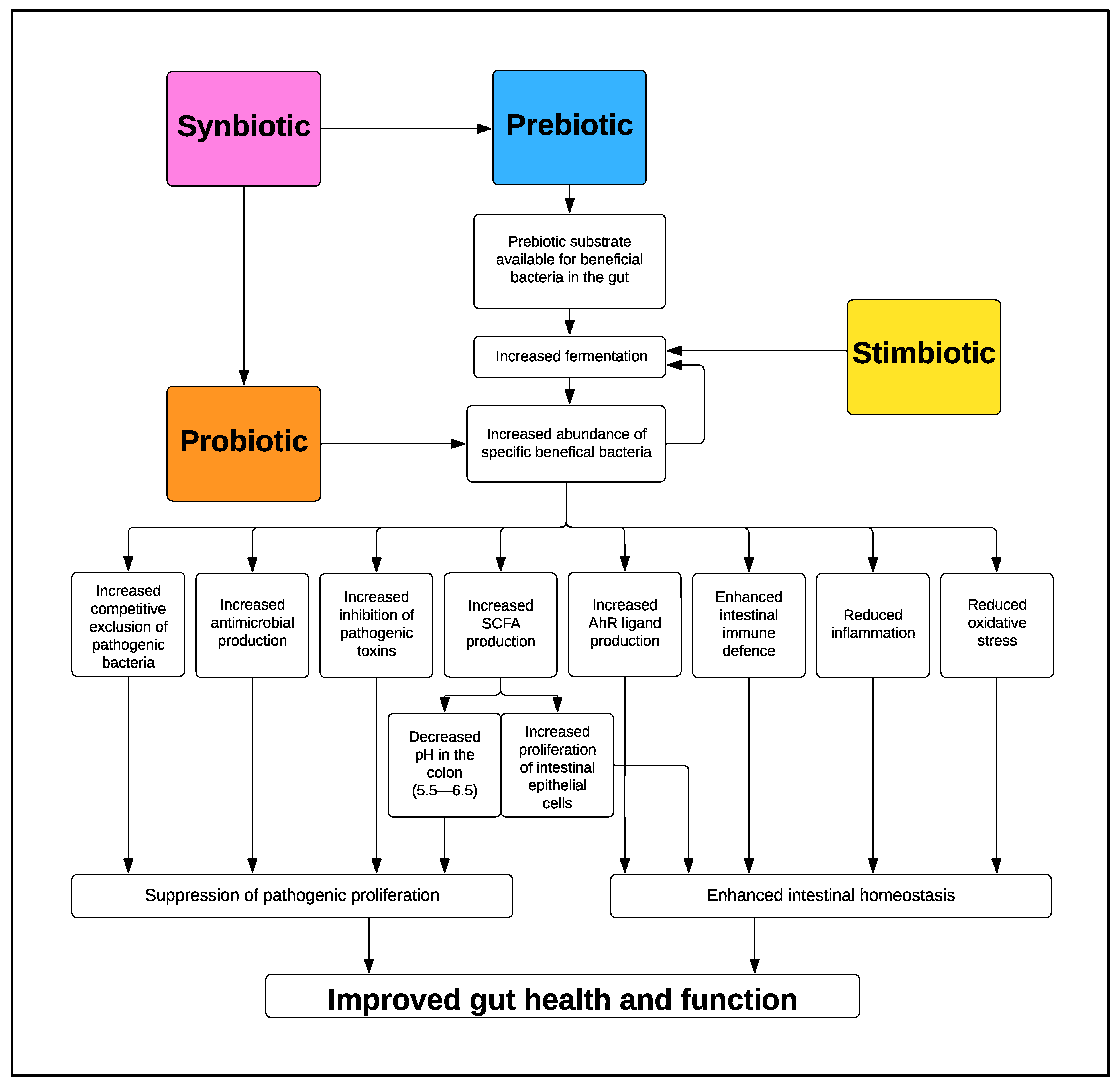
2. Prebiotics
2.1. Traditional Prebiotics
2.1.1. Beta Glucans (β-Glucans)
2.1.2. Non-Digestible Oligosaccharides
2.1.3. Inulin
2.1.4. Resistant Starch
2.1.5. Pectin
2.2. Novel Prebiotics
2.2.1. Proteins, Hydrolysates, Peptides, and Amino Acids
2.2.2. Nucleotides
2.2.3. Polyphenols
3. Stimbiotics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deng, F.; Li, Y.; Peng, Y.; Wei, X.; Wang, X.; Howe, S.; Yang, H.; Xiao, Y.; Li, H.; Zhao, J.; et al. The Diversity, Composition, and Metabolic Pathways of Archaea in Pigs. Animals 2021, 11, 2139. [Google Scholar] [CrossRef]
- Qin, J.; Ji, B.; Ma, Y.; Liu, X.; Wang, T.; Liu, G.; Li, B.; Wang, G.; Gao, P. Diversity and potential function of pig gut DNA viruses. Heliyon 2023, 9, e14020. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Prenafeta-Boldú, F.; Zingaretti, L.M.; Gonzalez-Rodriguez, O.; Dalmau, A.; Quintanilla, R.; Ballester, M. Gut eukaryotic communities in pigs: Diversity, composition and host genetics contribution. Anim. Microbiome 2020, 2, 18. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef]
- Oh, J.K.; Chae, J.P.; Pajarillo, E.A.B.; Kim, S.H.; Kwak, M.J.; Eun, J.S.; Chee, S.W.; Whang, K.Y.; Kim, S.H.; Kang, D.K. Association between the body weight of growing pigs and the functional capacity of their gut microbiota. Anim. Sci. J. 2020, 91, e13418. [Google Scholar] [CrossRef]
- Déru, V.; Bouquet, A.; Zemb, O.; Blanchet, B.; De Almeida, M.L.; Cauquil, L.; Carillier-Jacquin, C.; Gilbert, H. Genetic relationships between efficiency traits and gut microbiota traits in growing pigs being fed with a conventional or a high-fiber diet. J. Anim. Sci. 2022, 100, skac183. [Google Scholar] [CrossRef]
- Yang, Q.; Huang, X.; Zhao, S.; Sun, W.; Yan, Z.; Wang, P.; Li, S.; Huang, W.; Zhang, S.; Liu, L. Structure and function of the fecal microbiota in diarrheic neonatal piglets. Front. Microbiol. 2017, 8, 502. [Google Scholar] [CrossRef]
- McCormack, U.M.; Curião, T.; Wilkinson, T.; Metzler-Zebeli, B.U.; Reyer, H.; Ryan, T.; Calderon-Diaz, J.A.; Crispie, F.; Cotter, P.D.; Creevey, C.J. Fecal microbiota transplantation in gestating sows and neonatal offspring alters lifetime intestinal microbiota and growth in offspring. MSystems 2018, 3, e00134-17. [Google Scholar] [CrossRef]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. Probiotic Strains as a Potential Tool for Limiting the Use of Antibiotics, and Improving the Growth and Health of Pigs and Chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- Leblois, J.; Massart, S.; Li, B.; Wavreille, J.; Bindelle, J.; Everaert, N. Modulation of piglets’ microbiota: Differential effects by a high wheat bran maternal diet during gestation and lactation. Sci. Rep. 2017, 7, 7426. [Google Scholar] [CrossRef]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- Ardissone, A.N.; de la Cruz, D.M.; Davis-Richardson, A.G.; Rechcigl, K.T.; Li, N.; Drew, J.C.; Murgas-Torrazza, R.; Sharma, R.; Hudak, M.L.; Triplett, E.W. Meconium microbiome analysis identifies bacteria correlated with premature birth. PLoS ONE 2014, 9, e90784. [Google Scholar] [CrossRef]
- Chen, X.; Xu, J.; Ren, E.; Su, Y.; Zhu, W. Co-occurrence of early gut colonization in neonatal piglets with microbiota in the maternal and surrounding delivery environments. Anaerobe 2018, 49, 30–40. [Google Scholar] [CrossRef]
- Salcedo, J.; Frese, S.A.; Mills, D.A.; Barile, D. Characterization of porcine milk oligosaccharides during early lactation and their relation to the fecal microbiome. J. Dairy Sci. 2016, 99, 7733–7743. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning Stress Perturbs Gut Microbiome and Its Metabolic Profile in Piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning Anorexia May Contribute to Local Inflammation in the Piglet Small Intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef]
- Luppi, A.; Gibellini, M.; Gin, T.; Vangroenweghe, F.; Vandenbroucke, V.; Bauerfeind, R.; Bonilauri, P.; Labarque, G.; Hidalgo, Á. Prevalence of virulence factors in enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea in Europe. Porc. Health Manag. 2016, 2, 20. [Google Scholar] [CrossRef]
- Dou, S.; Gadonna-Widehem, P.; Rome, V.; Hamoudi, D.; Rhazi, L.; Lakhal, L.; Larcher, T.; Bahi-Jaber, N.; Pinon-Quintana, A.; Guyonvarch, A.; et al. Characterisation of Early-Life Fecal Microbiota in Susceptible and Healthy Pigs to Post-Weaning Diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef]
- Rattigan, R.; Lawlor, P.G.; Cormican, P.; Crespo-Piazuelo, D.; Cullen, J.; Phelan, J.P.; Ranjitkar, S.; Crispie, F.; Gardiner, G.E. Maternal and/or post-weaning supplementation with Bacillus altitudinis spores modulates the microbial composition of colostrum, digesta and faeces in pigs. Sci. Rep. 2023, 13, 8900. [Google Scholar] [CrossRef]
- Kiernan, D.P.; O’Doherty, J.V.; Sweeney, T. The effect of maternal probiotic or synbiotic supplementation on sow and offspring microbiota, health, and performance. Animals 2023, 13, 2996. [Google Scholar] [CrossRef]
- Gibson, G.R.; Scott, K.P.; Rastall, R.A.; Tuohy, K.M.; Hotchkiss, A.; Dubert-Ferrandon, A.; Gareau, M.; Murphy, E.F.; Saulnier, D.; Loh, G. Dietary prebiotics: Current status and new definition. Food Sci. Technol. Bull. Funct. Foods 2010, 7, 1–19. [Google Scholar] [CrossRef]
- González-Ortiz, G.; Gomes, G.; Dos Santos, T.; Bedford, M. New Strategies Influencing Gut Functionality and Animal Performance. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 233–254. [Google Scholar]
- Shin, D.; Chang, S.Y.; Bogere, P.; Won, K.; Choi, J.-Y.; Choi, Y.-J.; Lee, H.K.; Hur, J.; Park, B.-Y.; Kim, Y.; et al. Beneficial roles of probiotics on the modulation of gut microbiota and immune response in pigs. PLoS ONE 2019, 14, e0220843. [Google Scholar] [CrossRef]
- Chlebicz-Wójcik, A.; Śliżewska, K. The Effect of Recently Developed Synbiotic Preparations on Dominant Fecal Microbiota and Organic Acids Concentrations in Feces of Piglets from Nursing to Fattening. Animals 2020, 10, 1999. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- FAO/WHO Expert Consultation, Amerian Córdoba Park Hotel, Córdoba, Argentina. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Prevention 2001, 5, 1–10. Available online: https://www.iqb.es/digestivo/pdfs/probioticos.pdf (accessed on 10 August 2023).
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]
- Luo, J.; Chen, D.; Mao, X.; He, J.; Yu, B.; Cheng, L.; Zeng, D. Purified β-glucans of Different Molecular Weights Enhance Growth Performance of LPS-challenged Piglets via Improved Gut Barrier Function and Microbiota. Animals 2019, 9, 602. [Google Scholar] [CrossRef]
- Liu, J.B.; Cao, S.C.; Liu, J.; Xie, Y.N.; Zhang, H.F. Effect of probiotics and xylo-oligosaccharide supplementation on nutrient digestibility, intestinal health and noxious gas emission in weanling pigs. Asian Australas J. Anim. Sci. 2018, 31, 1660–1669. [Google Scholar] [CrossRef]
- Dowley, A.; Sweeney, T.; Conway, E.; Vigors, S.; Ryan, M.T.; Yadav, S.; Wilson, J.; O’Doherty, J.V. The effects of dietary supplementation with mushroom or selenium enriched mushroom powders on the growth performance and intestinal health of post-weaned pigs. J. Anim. Sci. Biotechnol. 2022, 14, 12. [Google Scholar] [CrossRef]
- Dowley, A.; Sweeney, T.; Conway, E.; Vigors, S.; Yadav, S.; Wilson, J.; Gabrielli, W.; O’Doherty, J.V. Effects of Dietary Supplementation with Mushroom or Vitamin D2-Enriched Mushroom Powders on Gastrointestinal Health Parameters in the Weaned Pig. Animals 2021, 11, 3603. [Google Scholar] [CrossRef]
- Conway, E.; Sweeney, T.; Dowley, A.; Vigors, S.; Ryan, M.; Yadav, S.; Wilson, J.; O’Doherty, J.V. Selenium-Enriched Mushroom Powder Enhances Intestinal Health and Growth Performance in the Absence of Zinc Oxide in Post-Weaned Pig Diets. Animals 2022, 12, 1503. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, Y.; Zhang, G.; Liu, L.; Lai, C. Relationship between Dietary Fiber Fermentation and Volatile Fatty Acids’ Concentration in Growing Pigs. Animals 2020, 10, 263. [Google Scholar] [CrossRef]
- Beaumont, M.; Roura, E.; Lambert, W.; Turni, C.; Michiels, J.; Chalvon-Demersay, T. Selective nourishing of gut microbiota with amino acids: A novel prebiotic approach? Front. Nutr. 2022, 9, 1066898. [Google Scholar] [CrossRef]
- Ding, T.; Xu, M.; Li, Y. An Overlooked Prebiotic: Beneficial Effect of Dietary Nucleotide Supplementation on Gut Microbiota and Metabolites in Senescence-Accelerated Mouse Prone-8 Mice. Front. Nutr. 2022, 9, 820799. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-Mediated Gut Microbiota Modulation: Toward Prebiotics and Further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef]
- Ferreira-Lazarte, A.; Moreno, F.J.; Villamiel, M. Bringing the digestibility of prebiotics into focus: Update of carbohydrate digestion models. Crit. Rev. Food Sci. Nutr. 2021, 61, 3267–3278. [Google Scholar] [CrossRef]
- Julio-Gonzalez, L.C.; Hernandez-Hernandez, O.; Moreno, F.J.; Olano, A.; Jimeno, M.L.; Corzo, N. Trans-β-galactosidase activity of pig enzymes embedded in the small intestinal brush border membrane vesicles. Sci. Rep. 2019, 9, 960. [Google Scholar] [CrossRef]
- Song, H.; Jeon, D.; Unno, T. Evaluation of Prebiotics through an In Vitro Gastrointestinal Digestion and Fecal Fermentation Experiment: Further Idea on the Implementation of Machine Learning Technique. Foods 2022, 11, 2490. [Google Scholar] [CrossRef]
- Park, J.-h.; Kotani, T.; Konno, T.; Setiawan, J.; Kitamura, Y.; Imada, S.; Usui, Y.; Hatano, N.; Shinohara, M.; Saito, Y.; et al. Promotion of Intestinal Epithelial Cell Turnover by Commensal Bacteria: Role of Short-Chain Fatty Acids. PLoS ONE 2016, 11, e0156334. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yu, B.; Sun, J.; Liu, Z.; Chen, H.; Ge, L.; Chen, D. Short-chain fatty acids can improve lipid and glucose metabolism independently of the pig gut microbiota. J. Anim. Sci. Biotechnol. 2021, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.R.; Diao, H.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Huang, Z.Q.; Luo, Y.H.; Luo, J.Q.; Mao, X.B.; et al. Oral administration of short chain fatty acids could attenuate fat deposition of pigs. PLoS ONE 2018, 13, e0196867. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, Y.; Luo, J.; Mao, X.; Chen, D. Short chain fatty acids could prevent fat deposition in pigs via regulating related hormones and genes. Food Funct. 2020, 11, 1845–1855. [Google Scholar] [CrossRef]
- Souza da Silva, C.; Haenen, D.; Koopmans, S.J.; Hooiveld, G.J.E.J.; Bosch, G.; Bolhuis, J.E.; Kemp, B.; Müller, M.; Gerrits, W.J.J. Effects of resistant starch on behaviour, satiety-related hormones and metabolites in growing pigs. Animal 2014, 8, 1402–1411. [Google Scholar] [CrossRef]
- Souza da Silva, C.; van den Borne, J.J.G.C.; Gerrits, W.J.J.; Kemp, B.; Bolhuis, J.E. Effects of dietary fibers with different physicochemical properties on feeding motivation in adult female pigs. Physiol. Behav. 2012, 107, 218–230. [Google Scholar] [CrossRef]
- Astó, E.; Méndez, I.; Rodríguez-Prado, M.; Cuñé, J.; Espadaler, J.; Farran-Codina, A. Effect of the Degree of Polymerization of Fructans on Ex Vivo Fermented Human Gut Microbiome. Nutrients 2019, 11, 1293. [Google Scholar] [CrossRef]
- Ito, H.; Takemura, N.; Sonoyama, K.; Kawagishi, H.; Topping, D.L.; Conlon, M.A.; Morita, T. Degree of Polymerization of Inulin-Type Fructans Differentially Affects Number of Lactic Acid Bacteria, Intestinal Immune Functions, and Immunoglobulin A Secretion in the Rat Cecum. J. Agric. Food Chem. 2011, 59, 5771–5778. [Google Scholar] [CrossRef]
- Li, J.; Xing, J.; Li, D.; Wang, X.; Zhao, L.; Lv, S.; Huang, D. Effects of β-glucan extracted from Saccharomyces cerevisiae on humoral and cellular immunity in weaned piglets. Arch. Anim. Nutr. 2005, 59, 303–312. [Google Scholar] [CrossRef]
- Shang, H.-M.; Zhou, H.-Z.; Yang, J.-Y.; Li, R.; Song, H.; Wu, H.-X. In vitro and in vivo antioxidant activities of inulin. PLoS ONE 2018, 13, e0192273. [Google Scholar] [CrossRef]
- Khasina, E.; Kolenchenko, E.; Sgrebneva, M.; Kovalev, V.; Khotimchenko, Y.S. Antioxidant activities of a low etherified pectin from the seagrass Zostera marina. Russ. J. Mar. Biol. 2003, 29, 259–261. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, G.; Zhang, S.; Zhao, J. Fructooligosaccharide reduces weanling pig diarrhea in conjunction with improving intestinal antioxidase activity and tight junction protein expression. Nutrients 2022, 14, 512. [Google Scholar] [CrossRef] [PubMed]
- Farabegoli, F.; Santaclara, F.J.; Costas, D.; Alonso, M.; Abril, A.G.; Espiñeira, M.; Ortea, I.; Costas, C. Exploring the Anti-Inflammatory Effect of Inulin by Integrating Transcriptomic and Proteomic Analyses in a Murine Macrophage Cell Model. Nutrients 2023, 15, 859. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-Glucan Metabolic and Immunomodulatory Properties and Potential for Clinical Application. J. Fungi 2020, 6, 356. [Google Scholar] [CrossRef]
- Zhao, J.; Cheung, P.C.K. Fermentation of β-Glucans Derived from Different Sources by Bifidobacteria: Evaluation of Their Bifidogenic Effect. J. Agric. Food Chem. 2011, 59, 5986–5992. [Google Scholar] [CrossRef]
- Johansson, L.; Virkki, L.; Maunu, S.; Lehto, M.; Ekholm, P.; Varo, P. Structural characterization of water soluble β-glucan of oat bran. Carbohydr. Polym. 2000, 42, 143–148. [Google Scholar] [CrossRef]
- Manners, D.J.; Masson, A.J.; Patterson, J.C. The structure of a β-(1→3)-D-glucan from yeast cell walls. Biochem. J. 1973, 135, 19–30. [Google Scholar] [CrossRef]
- Lazaridou, A.; Kritikopoulou, K.; Biliaderis, C.G. Barley β-glucan cryogels as encapsulation carriers of proteins: Impact of molecular size on thermo-mechanical and release properties. Bioact. Carbohydr. Diet. Fibre 2015, 6, 99–108. [Google Scholar] [CrossRef]
- Mukhopadhya, A.; O’Doherty, J.V.; Sweeney, T. A combination of yeast beta-glucan and milk hydrolysate is a suitable alternative to zinc oxide in the race to alleviate post-weaning diarrhoea in piglets. Sci. Rep. 2019, 9, 616. [Google Scholar] [CrossRef]
- Bouwhuis, M.A.; Sweeney, T.; Mukhopadhya, A.; McDonnell, M.J.; O’Doherty, J.V. Maternal laminarin supplementation decreases Salmonella Typhimurium shedding and improves intestinal health in piglets following an experimental challenge with S. Typhimurium post-weaning. Anim. Feed. Sci. Technol. 2017, 223, 156–168. [Google Scholar] [CrossRef]
- Heim, G.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of maternal dietary supplementation of laminarin and fucoidan, independently or in combination, on pig growth performance and aspects of intestinal health. Anim. Feed. Sci. Technol. 2015, 204, 28–41. [Google Scholar] [CrossRef]
- Gyawali, R.; Minor, R.C.; Donovan, B.; Ibrahim, S.A. Inclusion of Oat in Feeding Can Increase the Potential Probiotic Bifidobacteria in Sow Milk. Animals 2015, 5, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.W.; Hong, J.; Kim, H.J.; Kang, S.W.; Kim, Y.Y. Effects of β-glucan with vitamin E supplementation on the physiological response, litter performance, blood profiles, immune response, and milk composition of lactating sows. Anim. Biosci. 2023, 36, 264–274. [Google Scholar] [CrossRef]
- dos Santos, M.C.; da Silva, K.F.; Bastos, A.P.A.; Félix, A.P.; de Oliveira, S.G.; Maiorka, A. Effect of yeast extracted β-glucans on the immune response and reproductive performance of gilts in the adaptation, gestation, and lactation periods. Livest. Sci. 2023, 275, 105289. [Google Scholar] [CrossRef]
- Arapovic, L.; Huang, Y.; Manell, E.; Verbeek, E.; Keeling, L.; Sun, L.; Landberg, R.; Lundh, T.; Lindberg, J.E.; Dicksved, J. Age Rather Than Supplementation with Oat β-Glucan Influences Development of the Intestinal Microbiota and SCFA Concentrations in Suckling Piglets. Animals 2023, 13, 1349. [Google Scholar] [PubMed]
- Kim, K.; Ehrlich, A.; Perng, V.; Chase, J.A.; Raybould, H.; Li, X.; Atwill, E.R.; Whelan, R.; Sokale, A.; Liu, Y. Algae-derived β-glucan enhanced gut health and immune responses of weaned pigs experimentally infected with a pathogenic E. coli. Anim. Feed. Sci. Technol. 2019, 248, 114–125. [Google Scholar] [CrossRef]
- Zhou, Y.; Luo, Y.; Yu, B.; Zheng, P.; Yu, J.; Huang, Z.; Mao, X.; Luo, J.; Yan, H.; He, J. Agrobacterium sp. ZX09 Beta-Glucan Attenuates Enterotoxigenic Escherichia coli-Induced Disruption of Intestinal Epithelium in Weaned Pigs. Int. J. Mol. Sci. 2022, 23, 10290. [Google Scholar] [CrossRef]
- Conway, E.; Sweeney, T.; Dowley, A.; Maher, S.; Rajauria, G.; Yadav, S.; Wilson, J.; Gabrielli, W.; O’Doherty, J.V. The effects of mushroom powder and vitamin D2-enriched mushroom powder supplementation on the growth performance and health of newly weaned pigs. J. Anim. Physiol. Anim. Nutr. 2022, 106, 517–527. [Google Scholar] [CrossRef]
- Park, J.-H.; Lee, S.-I.; Kim, I.-H. Effect of dietary β-glucan supplementation on growth performance, nutrient digestibility, and characteristics of feces in weaned pigs. J. Appl. Anim. Res. 2018, 46, 1193–1197. [Google Scholar] [CrossRef]
- de Vries, H.; Geervliet, M.; Jansen, C.A.; Rutten, V.P.M.G.; van Hees, H.; Groothuis, N.; Wells, J.M.; Savelkoul, H.F.J.; Tijhaar, E.; Smidt, H. Impact of Yeast-Derived β-Glucans on the Porcine Gut Microbiota and Immune System in Early Life. Microorganisms 2020, 8, 1573. [Google Scholar] [CrossRef]
- Zhou, T.X.; Jung, J.H.; Zhang, Z.F.; Kim, I.H. Effect of dietary β-glucan on growth performance, fecal microbial shedding and immunological responses after lipopolysaccharide challenge in weaned pigs. Anim. Feed. Sci. Technol. 2013, 179, 85–92. [Google Scholar] [CrossRef]
- Sweeney, T.; Collins, C.B.; Reilly, P.; Pierce, K.M.; Ryan, M.; O’Doherty, J.V. Effect of purified β-glucans derived from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae on piglet performance, selected bacterial populations, volatile fatty acids and pro-inflammatory cytokines in the gastrointestinal tract of pigs. Br. J. Nutr. 2012, 108, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Bearson, S.M.D.; Trachsel, J.M.; Bearson, B.L.; Loving, C.L.; Kerr, B.J.; Shippy, D.C.; Kiros, T.G. Effects of β-glucan on Salmonella enterica serovar Typhimurium swine colonization and microbiota alterations. Porc. Health Manag. 2023, 9, 7. [Google Scholar] [CrossRef]
- Luo, J.; Zeng, D.; Cheng, L.; Mao, X.; Yu, J.; Yu, B.; Chen, D. Dietary β-glucan supplementation improves growth performance, carcass traits and meat quality of finishing pigs. Anim. Nutr. 2019, 5, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Dowley, A.; Sweeney, T.; Conway, E.; Maher, S.; Rajauria, G.; Yadav, S.; Wilson, J.; Gabrielli, W.; O’Doherty, J.V. The effects of dietary supplementation with mushroom or vitamin D2 enriched mushroom powders on finisher pig performance and meat quality. Anim. Feed. Sci. Technol. 2022, 288, 115313. [Google Scholar] [CrossRef]
- Tiwari, U.P.; Singh, A.K.; Jha, R. Fermentation characteristics of resistant starch, arabinoxylan, and β-glucan and their effects on the gut microbial ecology of pigs: A review. Anim. Nutr. 2019, 5, 217–226. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Zhou, X.; Xu, Y. Selective Production of Xylooligosaccharides by Xylan Hydrolysis Using a Novel Recyclable and Separable Furoic Acid. Front. Bioeng. Biotechnol. 2021, 9, 660266. [Google Scholar] [CrossRef]
- Chikkerur, J.; Samanta, A.K.; Kolte, A.P.; Dhali, A.; Roy, S. Production of Short Chain Fructo-oligosaccharides from Inulin of Chicory Root Using Fungal Endoinulinase. Appl. Biochem. Biotechnol. 2020, 191, 695–715. [Google Scholar] [CrossRef]
- Xing, Y.; Li, K.; Xu, Y.; Wu, Y.; Shi, L.; Guo, S.; Yan, S.; Jin, X.; Shi, B. Effects of galacto-oligosaccharide on growth performance, feacal microbiota, immune response and antioxidant capability in weaned piglets. J. Appl. Anim. Res. 2020, 48, 63–69. [Google Scholar] [CrossRef]
- Capitán-Cañadas, F.; Ortega-González, M.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; de Medina, F.S.; Martínez-Augustin, O. Prebiotic oligosaccharides directly modulate proinflammatory cytokine production in monocytes via activation of TLR4. Mol. Nutr. Food Res. 2014, 58, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Eiwegger, T.; Stahl, B.; Haidl, P.; Schmitt, J.; Boehm, G.; Dehlink, E.; Urbanek, R.; Szépfalusi, Z. Prebiotic oligosaccharides: In vitro evidence for gastrointestinal epithelial transfer and immunomodulatory properties. Pediatr. Allergy Immunol. 2010, 21, 1179–1188. [Google Scholar] [CrossRef]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. HUMAN MILK GLYCANS PROTECT INFANTS AGAINST ENTERIC PATHOGENS. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Zeng, D.; Wang, H.; Sun, N.; Zhao, Y.; Dan, Y.; Pan, K.; Jing, B.; Ni, X. Probiotic Lactobacillus johnsonii BS15 Promotes Growth Performance, Intestinal Immunity, and Gut Microbiota in Piglets. Probiotics Antimicrob. Proteins 2020, 12, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Pahumunto, N.; Dahlen, G.; Teanpaisan, R. Evaluation of Potential Probiotic Properties of Lactobacillus and Bacillus Strains Derived from Various Sources for Their Potential Use in Swine Feeding. Probiotics Antimicrob. Proteins 2021, 15, 479–490. [Google Scholar] [CrossRef]
- Xie, C.; Guo, X.; Long, C.; Fan, Z.; Xiao, D.; Ruan, Z.; Deng, Z.-y.; Wu, X.; Yin, Y. Supplementation of the sow diet with chitosan oligosaccharide during late gestation and lactation affects hepatic gluconeogenesis of suckling piglets. Anim. Reprod. Sci. 2015, 159, 109–117. [Google Scholar] [CrossRef]
- Duan, X.; Tian, G.; Chen, D.; Yang, J.; Zhang, L.; Li, B.; Huang, L.; Zhang, D.; Zheng, P.; Mao, X.; et al. Effects of diet chitosan oligosaccharide on performance and immune response of sows and their offspring. Livest. Sci. 2020, 239, 104114. [Google Scholar] [CrossRef]
- Le Bourgot, C.; Le Normand, L.; Formal, M.; Respondek, F.; Blat, S.; Apper, E.; Ferret-Bernard, S.; Le Huërou-Luron, I. Maternal short-chain fructo-oligosaccharide supplementation increases intestinal cytokine secretion, goblet cell number, butyrate concentration and Lawsonia intracellularis humoral vaccine response in weaned pigs. Br. J. Nutr. 2017, 117, 83–92. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, X.; Pi, Y.; Han, D.; Feng, C.; Zhao, J.; Chen, L.; Che, D.; Bao, H.; Xie, Z. Maternal galactooligosaccharides supplementation programmed immune defense, microbial colonization and intestinal development in piglets. Food Funct. 2021, 12, 7260–7270. [Google Scholar] [CrossRef]
- Duan, X.D.; Chen, D.W.; Zheng, P.; Tian, G.; Wang, J.P.; Mao, X.B.; Yu, J.; He, J.; Li, B.; Huang, Z.Q.; et al. Effects of dietary mannan oligosaccharide supplementation on performance and immune response of sows and their offspring. Anim. Feed. Sci. Technol. 2016, 218, 17–25. [Google Scholar] [CrossRef]
- Duan, X.; Tian, G.; Chen, D.; Huang, L.; Zhang, D.; Zheng, P.; Mao, X.; Yu, J.; He, J.; Huang, Z.; et al. Mannan oligosaccharide supplementation in diets of sow and (or) their offspring improved immunity and regulated intestinal bacteria in piglet1. J. Anim. Sci. 2019, 97, 4548–4556. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Piao, X.; Kim, S.; Wang, L.; Shen, Y.; Lee, H.; Li, S. Effects of chito-oligosaccharide supplementation on the growth performance, nutrient digestibility, intestinal morphology, and fecal shedding of Escherichia coli and Lactobacillus in weaning pigs. J. Anim. Sci. 2008, 86, 2609–2618. [Google Scholar] [CrossRef] [PubMed]
- Walsh, A.; Sweeney, T.; Bahar, B.; Flynn, B.; O’doherty, J. The effects of supplementing varying molecular weights of chitooligosaccharide on performance, selected microbial populations and nutrient digestibility in the weaned pig. Animal 2013, 7, 571–579. [Google Scholar] [CrossRef]
- Suthongsa, S.; Pichyangkura, R.; Kalandakanond-Thongsong, S.; Thongsong, B. Effects of dietary levels of chito-oligosaccharide on ileal digestibility of nutrients, small intestinal morphology and crypt cell proliferation in weaned pigs. Livest. Sci. 2017, 198, 37–44. [Google Scholar] [CrossRef]
- Mikkelsen, L.L.; Jakobsen, M.; Jensen, B.B. Effects of dietary oligosaccharides on microbial diversity and fructo-oligosaccharide degrading bacteria in faeces of piglets post-weaning. Anim. Feed. Sci. Technol. 2003, 109, 133–150. [Google Scholar] [CrossRef]
- Liu, L.; Chen, D.; Yu, B.; Yin, H.; Huang, Z.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; Luo, J. Fructooligosaccharides improve growth performance and intestinal epithelium function in weaned pigs exposed to enterotoxigenic Escherichia coli. Food Funct. 2020, 11, 9599–9612. [Google Scholar] [CrossRef]
- Sampath, V.; Park, J.; Shanmugam, S.; Kim, I. Lactulose as a potential additive to enhance the growth performance, nutrient digestibility, and microbial shedding, and diminish noxious odor emissions in weaning pigs. Korean J. Agric. Sci. 2021, 48, 965–973. [Google Scholar]
- Zhao, P.Y.; Jung, J.H.; Kim, I.H. Effect of mannan oligosaccharides and fructan on growth performance, nutrient digestibility, blood profile, and diarrhea score in weanling pigs1. J. Anim. Sci. 2012, 90, 833–839. [Google Scholar] [CrossRef]
- Sun, F.; Li, H.; Sun, Z.; Liu, L.; Zhang, X.; Zhao, J. Effect of Arabinoxylan and Xylo-Oligosaccharide on Growth Performance and Intestinal Barrier Function in Weaned Piglets. Animals 2023, 13, 964. [Google Scholar] [CrossRef]
- Cui, S.W.; Roberts, K.T. CHAPTER 13—Dietary Fiber: Fulfilling the Promise of Added-Value Formulations. In Modern Biopolymer Science; Kasapis, S., Norton, I.T., Ubbink, J.B., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 399–448. [Google Scholar]
- Van Bekkum, H.; Röper, H.; Voragen, A. Carbohydrates as Organic Raw Materials III; John Wiley & Sons: New York, NY, USA, 2008. [Google Scholar]
- Meyer, D.; Blaauwhoed, J.P. 30—Inulin. In Handbook of Hydrocolloids, 2nd ed.; Phillips, G.O., Williams, P.A., Eds.; Woodhead Publishing: Cambridge, UK, 2009; pp. 829–848. [Google Scholar]
- van Loo, J.; Coussement, P.; de Leenheer, L.; Hoebregs, H.; Smits, G. On the presence of inulin and oligofructose as natural ingredients in the western diet. Crit. Rev. Food Sci. Nutr. 1995, 35, 525–552. [Google Scholar] [CrossRef]
- Mensink, M.A.; Frijlink, H.W.; van der Voort Maarschalk, K.; Hinrichs, W.L. Inulin, a flexible oligosaccharide I: Review of its physicochemical characteristics. Carbohydr. Polym. 2015, 130, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Ronkart, S.N.; Blecker, C.S.; Fourmanoir, H.; Fougnies, C.; Deroanne, C.; Van Herck, J.-C.; Paquot, M. Isolation and identification of inulooligosaccharides resulting from inulin hydrolysis. Anal. Chim. Acta 2007, 604, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Faqih, M.N.; Wang, S.S. Factors affecting gel formation of inulin. Carbohydr. Polym. 2001, 46, 135–145. [Google Scholar] [CrossRef]
- Franck, A. Technological functionality of inulin and oligofructose. Br. J. Nutr. 2002, 87, S287–S291. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, Z.; Lyu, H.; Gu, X.; Song, Z.; He, X.; Fan, Z. Effects of dietary inulin during late gestation on sow physiology, farrowing duration and piglet performance. Anim. Reprod. Sci. 2020, 219, 106531. [Google Scholar] [CrossRef]
- Pierce, K.M.; Callan, J.J.; McCarthy, P.; O’Doherty, J.V. Performance of weanling pigs offered low or high lactose diets supplemented with avilamycin or inulin. Anim. Sci. 2005, 80, 313–318. [Google Scholar] [CrossRef]
- Yasuda, K.; Dawson, H.D.; Wasmuth, E.V.; Roneker, C.A.; Chen, C.; Urban, J.F.; Welch, R.M.; Miller, D.D.; Lei, X.G. Supplemental Dietary Inulin Influences Expression of Iron and Inflammation Related Genes in Young Pigs. J. Nutr. 2009, 139, 2018–2023. [Google Scholar] [CrossRef]
- Patterson, J.K.; Yasuda, K.; Welch, R.M.; Miller, D.D.; Lei, X.G. Supplemental Dietary Inulin of Variable Chain Lengths Alters Intestinal Bacterial Populations in Young Pigs. J. Nutr. 2010, 140, 2158–2161. [Google Scholar] [CrossRef]
- Eberhard, M.; Hennig, U.; Kuhla, S.; Brunner, R.M.; Kleessen, B.; Metges, C.C. Effect of inulin supplementation on selected gastric, duodenal, and caecal microbiota and short chain fatty acid pattern in growing piglets. Arch. Anim. Nutr. 2007, 61, 235–246. [Google Scholar] [CrossRef]
- Paßlack, N.; Vahjen, W.; Zentek, J. Dietary inulin affects the intestinal microbiota in sows and their suckling piglets. BMC Vet. Res. 2015, 11, 51. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, L.; Xia, B.; Tang, S.; Liu, L.; Xie, J.; Zhang, H. Bioregional Alterations in Gut Microbiome Contribute to the Plasma Metabolomic Changes in Pigs Fed with Inulin. Microorganisms 2020, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.R.; Świątkiewicz, M.; Florek, M.; Bąkowski, M.; Skiba, G. Effect of Inulin Source and a Probiotic Supplement in Pig Diets on Carcass Traits, Meat Quality and Fatty Acid Composition in Finishing Pigs. Animals 2021, 11, 2438. [Google Scholar] [CrossRef]
- Tan, F.P.Y.; Beltranena, E.; Zijlstra, R.T. Resistant starch: Implications of dietary inclusion on gut health and growth in pigs: A review. J. Anim. Sci. Biotechnol. 2021, 12, 124. [Google Scholar] [CrossRef]
- Regassa, A.; Nyachoti, C.M. Application of resistant starch in swine and poultry diets with particular reference to gut health and function. Anim. Nutr. 2018, 4, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Canibe, N.; Montagne, L.; Freire, J.; Bosi, P.; Prates, J.A.M.; Tanghe, S.; Trevisi, P. Resistant starch reduces large intestinal pH and promotes fecal lactobacilli and bifidobacteria in pigs. Animal 2019, 13, 64–73. [Google Scholar] [CrossRef]
- Trachsel, J.; Briggs, C.; Gabler, N.K.; Allen, H.K.; Loving, C.L. Dietary resistant potato starch alters intestinal microbial communities and their metabolites, and markers of immune regulation and barrier function in swine. Front. Immunol. 2019, 10, 1381. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Agyekum, A.K.; Yin, Y.L.; Rideout, T.C.; Nyachoti, C.M. Feeding a diet containing resistant potato starch influences gastrointestinal tract traits and growth performance of weaned pigs. J. Anim. Sci. 2014, 92, 3906–3913. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.K.; Nyachoti, C.M.; Krause, D.O. Raw potato starch in weaned pig diets and its influence on postweaning scours and the molecular microbial ecology of the digestive tract1. J. Anim. Sci. 2009, 87, 984–993. [Google Scholar] [CrossRef]
- Yan, H.; Lu, H.; Almeida, V.V.; Ward, M.G.; Adeola, O.; Nakatsu, C.H.; Ajuwon, K.M. Effects of dietary resistant starch content on metabolic status, milk composition, and microbial profiling in lactating sows and on offspring performance. J. Anim. Physiol. Anim. Nutr. 2017, 101, 190–200. [Google Scholar] [CrossRef]
- Leblois, J.; Massart, S.; Soyeurt, H.; Grelet, C.; Dehareng, F.; Schroyen, M.; Li, B.; Wavreille, J.; Bindelle, J.; Everaert, N. Feeding sows resistant starch during gestation and lactation impacts their faecal microbiota and milk composition but shows limited effects on their progeny. PLoS ONE 2018, 13, e0199568. [Google Scholar] [CrossRef]
- Schroyen, M.; Leblois, J.; Uerlings, J.; Li, B.; Sureda, E.A.; Massart, S.; Wavreille, J.; Bindelle, J.; Everaert, N. Maternal dietary resistant starch does not improve piglet’s gut and liver metabolism when challenged with a high fat diet. BMC Genom. 2020, 21, 439. [Google Scholar] [CrossRef] [PubMed]
- Souza da Silva, C.; Bosch, G.; Bolhuis, J.E.; Stappers, L.J.N.; van Hees, H.M.J.; Gerrits, W.J.J.; Kemp, B. Effects of alginate and resistant starch on feeding patterns, behaviour and performance in ad libitum-fed growing pigs. Animal 2014, 8, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.A. Reassessment of some fruit and vegetable pectin levels. J. Food Sci. 1997, 62, 225–229. [Google Scholar] [CrossRef]
- Wen, X.; Zhong, R.; Dang, G.; Xia, B.; Wu, W.; Tang, S.; Tang, L.; Liu, L.; Liu, Z.; Chen, L.; et al. Pectin supplementation ameliorates intestinal epithelial barrier function damage by modulating intestinal microbiota in lipopolysaccharide-challenged piglets. J. Nutr. Biochem. 2022, 109, 109107. [Google Scholar] [CrossRef] [PubMed]
- Dang, G.; Wen, X.; Zhong, R.; Wu, W.; Tang, S.; Li, C.; Yi, B.; Chen, L.; Zhang, H.; Schroyen, M. Pectin modulates intestinal immunity in a pig model via regulating the gut microbiota-derived tryptophan metabolite-AhR-IL22 pathway. J. Anim. Sci. Biotechnol. 2023, 14, 38. [Google Scholar] [CrossRef]
- Xu, R.; Li, Q.; Wang, H.; Su, Y.; Zhu, W. Reduction of Redox Potential Exerts a Key Role in Modulating Gut Microbial Taxa and Function by Dietary Supplementation of Pectin in a Pig Model. Microbiol. Spectr. 2023, 11, e0328322. [Google Scholar] [CrossRef]
- Low, D.Y.; Pluschke, A.M.; Flanagan, B.; Sonni, F.; Grant, L.J.; Williams, B.A.; Gidley, M.J. Isolated pectin (apple) and fruit pulp (mango) impact gastric emptying, passage rate and short chain fatty acid (SCFA) production differently along the pig gastrointestinal tract. Food Hydrocoll. 2021, 118, 106723. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, L.; Xia, B.; Tang, S.; Xie, J.; Zhang, H. Modulation of Pectin on Mucosal Innate Immune Function in Pigs Mediated by Gut Microbiota. Microorganisms 2020, 8, 535. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef]
- Larsen, N.; Bussolo de Souza, C.; Krych, L.; Barbosa Cahú, T.; Wiese, M.; Kot, W.; Hansen, K.M.; Blennow, A.; Venema, K.; Jespersen, L. Potential of Pectins to Beneficially Modulate the Gut Microbiota Depends on Their Structural Properties. Front. Microbiol. 2019, 10, 223. [Google Scholar] [CrossRef]
- Wiese, M. The potential of pectin to impact pig nutrition and health: Feeding the animal and its microbiome. FEMS Microbiol. Lett. 2019, 366, fnz029. [Google Scholar] [CrossRef] [PubMed]
- Dowley, A.; O’Doherty, J.V.; Mukhopadhya, A.; Conway, E.; Vigors, S.; Maher, S.; Ryan, M.T.; Sweeney, T. Maternal Supplementation With a Casein Hydrolysate and Yeast Beta-glucan from Late Gestation through Lactation Improves Gastrointestinal Health of Piglets at Weaning. Sci. Rep. 2022, 12, 17407. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.; O’Doherty, J.V.; Mukhopadhya, A.; Dowley, A.; Vigors, S.; Maher, S.; Ryan, M.T.; Sweeney, T. Maternal and/or direct supplementation with a combination of a casein hydrolysate and yeast β-glucan on post-weaning performance and intestinal health in the pig. PLoS ONE 2022, 17, e0265051. [Google Scholar] [CrossRef]
- Seifert, A.; Freilich, S.; Kashi, Y.; Livney, Y.D. Protein-oligosaccharide conjugates as novel prebiotics. Polym. Adv. Technol. 2019, 30, 2577–2585. [Google Scholar] [CrossRef]
- Ferrario, C.; Duranti, S.; Milani, C.; Mancabelli, L.; Lugli, G.A.; Turroni, F.; Mangifesta, M.; Viappiani, A.; Ossiprandi, M.C.; van Sinderen, D.; et al. Exploring Amino Acid Auxotrophy in Bifidobacterium bifidum PRL2010. Front. Microbiol. 2015, 6, 1331. [Google Scholar] [CrossRef]
- Saguir, F.M.; de Nadra, M.C.M. Improvement of a Chemically Defined Medium for the Sustained Growth of Lactobacillus plantarum: Nutritional Requirements. Curr. Microbiol. 2007, 54, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Nsogning, S.D.; Fischer, S.; Becker, T. Investigating on the fermentation behavior of six lactic acid bacteria strains in barley malt wort reveals limitation in key amino acids and buffer capacity. Food Microbiol. 2018, 73, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Rattigan, R.; Sweeney, T.; Maher, S.; Ryan, M.T.; Thornton, K.; O’Doherty, J.V. Effects of reducing dietary crude protein concentration and supplementation with either laminarin or zinc oxide on the growth performance and intestinal health of newly weaned pigs. Anim. Feed. Sci. Technol. 2020, 270, 114693. [Google Scholar] [CrossRef]
- Heo, J.; Kim, J.; Hansen, C.; Mullan, B.; Hampson, D.; Maribo, H.; Kjeldsen, N.; Pluske, J. Effects of dietary protein level and zinc oxide supplementation on the incidence of post-weaning diarrhoea in weaner pigs challenged with an enterotoxigenic strain of Escherichia coli. Livest. Sci. 2010, 133, 210–213. [Google Scholar] [CrossRef]
- Luise, D.; Chalvon-Demersay, T.; Lambert, W.; Bosi, P.; Trevisi, P. Meta-analysis to evaluate the impact of the reduction of dietary crude protein on the gut health of post-weaning pigs. Ital. J. Anim. Sci. 2021, 20, 1386–1397. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Ghyselinck, J.; Marzorati, M.; Koch, A.-M.; Lambert, W.; Michiels, J.; Chalvon-Demersay, T. The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota. Microorganisms 2022, 10, 762. [Google Scholar] [CrossRef]
- Goodarzi, P.; Wileman, C.M.; Habibi, M.; Walsh, K.; Sutton, J.; Shili, C.N.; Chai, J.; Zhao, J.; Pezeshki, A. Effect of Isoleucine and Added Valine on Performance, Nutrients Digestibility and Gut Microbiota Composition of Pigs Fed with Very Low Protein Diets. Int. J. Mol. Sci. 2022, 23, 14886. [Google Scholar] [CrossRef]
- Markman, G.; Livney, Y.D. Maillard-conjugate based core–shell co-assemblies for nanoencapsulation of hydrophobic nutraceuticals in clear beverages. Food Funct. 2012, 3, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, A.; Noronha, N.; Bahar, B.; Ryan, M.; Murray, B.; Kelly, P.; O’Loughlin, I.; O’Doherty, J.; Sweeney, T. The anti-inflammatory potential of a moderately hydrolysed casein and its 5 kDa fraction in in vitro and ex vivo models of the gastrointestinal tract. Food Funct. 2015, 6, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Su, Y.; Jia, F.; Jin, H. Characterization of casein hydrolysates derived from enzymatic hydrolysis. Chem. Cent. J. 2013, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Pinho, S. Viability of the microencapsulation of a casein hydrolysate in lipid microparticles of cupuacu butter and stearic acid. Int. J. Food Stud. 2013, 2, 48–59. [Google Scholar] [CrossRef]
- Vidal-Lletjós, S.; Beaumont, M.; Tomé, D.; Benamouzig, R.; Blachier, F.; Lan, A. Dietary protein and amino acid supplementation in inflammatory bowel disease course: What impact on the colonic mucosa? Nutrients 2017, 9, 310. [Google Scholar] [CrossRef]
- Liang, H.; Dai, Z.; Liu, N.; Ji, Y.; Chen, J.; Zhang, Y.; Yang, Y.; Li, J.; Wu, Z.; Wu, G. Dietary L-Tryptophan Modulates the Structural and Functional Composition of the Intestinal Microbiome in Weaned Piglets. Front. Microbiol. 2018, 9, 01736. [Google Scholar] [CrossRef]
- Liang, H.; Dai, Z.; Kou, J.; Sun, K.; Chen, J.; Yang, Y.; Wu, G.; Wu, Z. Dietary l-Tryptophan Supplementation Enhances the Intestinal Mucosal Barrier Function in Weaned Piglets: Implication of Tryptophan-Metabolizing Microbiota. Int. J. Mol. Sci. 2018, 20, 20. [Google Scholar] [CrossRef]
- Lamas, B.; Richard, M.L.; Leducq, V.; Pham, H.-P.; Michel, M.-L.; Da Costa, G.; Bridonneau, C.; Jegou, S.; Hoffmann, T.W.; Natividad, J.M.; et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 2016, 22, 598–605. [Google Scholar] [CrossRef]
- Rao, Z.; Li, J.; Shi, B.; Zeng, Y.; Liu, Y.; Sun, Z.; Wu, L.; Sun, W.; Tang, Z. Dietary Tryptophan Levels Impact Growth Performance and Intestinal Microbial Ecology in Weaned Piglets via Tryptophan Metabolites and Intestinal Antimicrobial Peptides. Animals 2021, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Lamas, B.; Natividad, J.M.; Sokol, H. Aryl hydrocarbon receptor and intestinal immunity. Mucosal Immunol. 2018, 11, 1024–1038. [Google Scholar] [CrossRef] [PubMed]
- Tossou, M.C.B.; Liu, H.; Bai, M.; Chen, S.; Cai, Y.; Duraipandiyan, V.; Liu, H.; Adebowale, T.O.; Al-Dhabi, N.A.; Long, L.; et al. Effect of High Dietary Tryptophan on Intestinal Morphology and Tight Junction Protein of Weaned Pig. BioMed Res. Int. 2016, 2016, 2912418. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Ma, Y.; Zhang, Z.; Fan, X.; Li, T.; Li, L.; Xiao, W.; Cai, Y.; Sun, L.; Xu, P.; et al. AhR activation protects intestinal epithelial barrier function through regulation of Par-6. J. Mol. Histol. 2018, 49, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Meng, D.; Sommella, E.; Salviati, E.; Campiglia, P.; Ganguli, K.; Djebali, K.; Zhu, W.; Walker, W.A. Indole-3-lactic acid, a metabolite of tryptophan, secreted by Bifidobacterium longum subspecies infantis is anti-inflammatory in the immature intestine. Pediatr. Res. 2020, 88, 209–217. [Google Scholar] [CrossRef]
- Zhang, L.; Nichols, R.G.; Correll, J.; Murray, I.A.; Tanaka, N.; Smith, P.B.; Hubbard, T.D.; Sebastian, A.; Albert, I.; Hatzakis, E. Persistent organic pollutants modify gut microbiota–host metabolic homeostasis in mice through aryl hydrocarbon receptor activation. Environ. Health Perspect. 2015, 123, 679–688. [Google Scholar] [CrossRef]
- Liu, G.; Lu, J.; Sun, W.; Jia, G.; Zhao, H.; Chen, X.; Kim, I.H.; Zhang, R.; Wang, J. Tryptophan supplementation enhances intestinal health by improving gut barrier function, alleviating inflammation, and modulating intestinal microbiome in lipopolysaccharide-challenged piglets. Front. Microbiol. 2022, 13, 2393. [Google Scholar] [CrossRef]
- Liu, G.; Tao, J.; Lu, J.; Jia, G.; Zhao, H.; Chen, X.; Tian, G.; Cai, J.; Zhang, R.; Wang, J. Dietary Tryptophan Supplementation Improves Antioxidant Status and Alleviates Inflammation, Endoplasmic Reticulum Stress, Apoptosis, and Pyroptosis in the Intestine of Piglets after Lipopolysaccharide Challenge. Antioxidants 2022, 11, 872. [Google Scholar] [CrossRef]
- Luise, D.; Bertocchi, M.; Bosi, P.; Correa, F.; Spinelli, E.; Trevisi, P. Contribution of L-Arginine supplementation during gestation on sow productive performance and on sow microbial faecal profile. Ital. J. Anim. Sci. 2020, 19, 330–340. [Google Scholar] [CrossRef]
- Kyoung, H.; Lee, J.J.; Cho, J.H.; Choe, J.; Kang, J.; Lee, H.; Liu, Y.; Kim, Y.; Kim, H.B.; Song, M. Dietary Glutamic Acid Modulates Immune Responses and Gut Health of Weaned Pigs. Animals 2021, 11, 504. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Xu, B.; Yin, B.; Xu, X.; Niu, Y.; Tang, Y.; Wang, X.; Xie, C.; Yang, T.; Zhou, S.; et al. Modulation of Gut Microbial Community and Metabolism by Dietary Glycyl-Glutamine Supplementation May Favor Weaning Transition in Piglets. Front. Microbiol. 2020, 10, 03125. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Gan, M.; Xie, Z.; Zhou, C.; Jing, Y.; Li, M.; Liu, C.; Wang, M.; Dai, H.; Huang, Z. Effects of Dietary L-Citrulline Supplementation on Growth Performance, Meat Quality and Fecal Microbial Composition in Finishing Pigs. Front. Microbiol. 2023, 14, 1209389. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, F.; Duan, Y.; Yin, Y.; Kong, X. Glutamic acid supplementation reduces body fat weight in finishing pigs when provided solely or in combination with arginine and it is associated with colonic propionate and butyrate concentrations. Food Funct. 2019, 10, 4693–4704. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, F.; Duan, Y.; Yin, Y.; Kong, X. Dietary Supplementation With Leucine or in Combination With Arginine Decreases Body Fat Weight and Alters Gut Microbiota Composition in Finishing Pigs. Front. Microbiol. 2019, 10, 01767. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Swine; The National Academic Press: Washington, DC, USA, 2012. [Google Scholar]
- Wu, C.; Yang, Z.; Song, C.; Liang, C.; Li, H.; Chen, W.; Lin, W.; Xie, Q. Effects of dietary yeast nucleotides supplementation on intestinal barrier function, intestinal microbiota, and humoral immunity in specific pathogen-free chickens. Poult. Sci. 2018, 97, 3837–3846. [Google Scholar] [CrossRef]
- Mateo, C.D.; Peters, D.N.; Stein, H.H. Nucleotides in sow colostrum and milk at different stages of lactation1,2,3. J. Anim. Sci. 2004, 82, 1339–1342. [Google Scholar] [CrossRef]
- Correa, F.; Luise, D.; Archetti, I.; Bosi, P.; Trevisi, P. Investigation of Early Supplementation of Nucleotides on the Intestinal Maturation of Weaned Piglets. Animals 2021, 11, 1489. [Google Scholar] [CrossRef]
- Gao, L.; Xie, C.; Liang, X.; Li, Z.; Li, B.; Wu, X.; Yin, Y. Yeast-based nucleotide supplementation in mother sows modifies the intestinal barrier function and immune response of neonatal pigs. Anim. Nutr. 2021, 7, 84–93. [Google Scholar] [CrossRef]
- Liu, G.; Liu, H.; Tian, W.; Liu, C.; Yang, H.; Wang, H.; Gao, L.; Huang, Y. Dietary nucleotides influences intestinal barrier function, immune responses and microbiota in 3-day-old weaned piglets. Int. Immunopharmacol. 2023, 117, 109888. [Google Scholar] [CrossRef] [PubMed]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbarek, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary supplementation with a nucleotide-rich yeast extract modulates gut immune response and microflora in weaned pigs in response to a sanitary challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- Sauer, N.; Eklund, M.; Bauer, E.; Gänzle, M.G.; Field, C.J.; Zijlstra, R.T.; Mosenthin, R. The effects of pure nucleotides on performance, humoral immunity, gut structure and numbers of intestinal bacteria of newly weaned pigs1. J. Anim. Sci. 2012, 90, 3126–3134. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, dietary sources and bioavailability. Ann. Ist. Super Sanita 2007, 43, 348–361. [Google Scholar] [PubMed]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Xu, X.; Hua, H.; Wang, L.; He, P.; Zhang, L.; Qin, Q.; Yu, C.; Wang, X.; Zhang, G.; Liu, Y. Holly polyphenols alleviate intestinal inflammation and alter microbiota composition in lipopolysaccharide-challenged pigs. Br. J. Nutr. 2020, 123, 881–891. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The two-way polyphenols-microbiota interactions and their effects on obesity and related metabolic diseases. Front. Nutr. 2019, 6, 188. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef]
- Jang, S.; Sun, J.; Chen, P.; Lakshman, S.; Molokin, A.; Harnly, J.M.; Vinyard, B.T.; Urban, J.F., Jr.; Davis, C.D.; Solano-Aguilar, G. Flavanol-Enriched Cocoa Powder Alters the Intestinal Microbiota, Tissue and Fluid Metabolite Profiles, and Intestinal Gene Expression in Pigs. J. Nutr. 2016, 146, 673–680. [Google Scholar] [CrossRef]
- Fiesel, A.; Gessner, D.K.; Most, E.; Eder, K. Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet. Res. 2014, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, P.; Wojtczak, L. Short-and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Sun, J.; Ge, L.; Liu, Z.; Chen, H.; Yu, B.; Chen, D. Exogenous infusion of short-chain fatty acids can improve intestinal functions independently of the gut microbiota. J. Anim. Sci. 2020, 98, skaa371. [Google Scholar] [CrossRef] [PubMed]
- Saleri, R.; Borghetti, P.; Ravanetti, F.; Cavalli, V.; Ferrari, L.; De Angelis, E.; Andrani, M.; Martelli, P. Effects of different short-chain fatty acids (SCFA) on gene expression of proteins involved in barrier function in IPEC-J2. Porc. Health Manag. 2022, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Lencina, C.; Painteaux, L.; Viémon-Desplanque, J.; Phornlaphat, O.; Lambert, W.; Chalvon-Demersay, T. A mix of functional amino acids and grape polyphenols promotes the growth of piglets, modulates the gut microbiota in vivo and regulates epithelial homeostasis in intestinal organoids. Amino Acids 2021, 54, 1357–1369. [Google Scholar] [CrossRef]
- Paniagua, M.; Villagómez-Estrada, S.; Crespo, F.J.; Pérez, J.F.; Arís, A.; Devant, M.; Solà-Oriol, D. Citrus Flavonoids Supplementation as an Alternative to Replace Zinc Oxide in Weanling Pigs&rsquo—Diets Minimizing the Use of Antibiotics. Animals 2023, 13, 967. [Google Scholar] [PubMed]
- Williams, A.R.; Krych, L.; Fauzan Ahmad, H.; Nejsum, P.; Skovgaard, K.; Nielsen, D.S.; Thamsborg, S.M. A polyphenol-enriched diet and Ascaris suum infection modulate mucosal immune responses and gut microbiota composition in pigs. PLoS ONE 2017, 12, e0186546. [Google Scholar] [CrossRef]
- Zheng, S.; Song, J.; Qin, X.; Yang, K.; Liu, M.; Yang, C.; Nyachoti, C.M. Dietary supplementation of red-osier dogwood polyphenol extract changes the ileal microbiota structure and increases Lactobacillus in a pig model. AMB Express 2021, 11, 145. [Google Scholar] [CrossRef]
- Cho, H.M.; González-Ortiz, G.; Melo-Durán, D.; Heo, J.M.; Cordero, G.; Bedford, M.R.; Kim, J.C. Stimbiotic supplementation improved performance and reduced inflammatory response via stimulating fiber fermenting microbiome in weaner pigs housed in a poor sanitary environment and fed an antibiotic-free low zinc oxide diet. PLoS ONE 2020, 15, e0240264. [Google Scholar] [CrossRef]
- Ebringerová, A.; Heinze, T. Xylan and xylan derivatives–biopolymers with valuable properties, 1. Naturally occurring xylans structures, isolation procedures and properties. Macromol. Rapid Commun. 2000, 21, 542–556. [Google Scholar] [CrossRef]
- Sutton, T.A.; O’Neill, H.V.M.; Bedford, M.R.; McDermott, K.; Miller, H.M. Effect of xylanase and xylo-oligosaccharide supplementation on growth performance and faecal bacterial community composition in growing pigs. Anim. Feed. Sci. Technol. 2021, 274, 114822. [Google Scholar] [CrossRef]
- Yin, J.; Li, F.; Kong, X.; Wen, C.; Guo, Q.; Zhang, L.; Wang, W.; Duan, Y.; Li, T.; Tan, Z. Dietary xylo-oligosaccharide improves intestinal functions in weaned piglets. Food Funct. 2019, 10, 2701–2709. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Wu, D.; Dai, Q. Effects of dietary xylo-oligosaccharide on growth performance, serum biochemical parameters, antioxidant function, and immunological function of nursery piglets. Rev. Bras. De Zootec. 2020, 49, e20190170. [Google Scholar] [CrossRef]
- Chen, W.; Yin, C.; Li, J.; Sun, W.; Li, Y.; Wang, C.; Pi, Y.; Cordero, G.; Li, X.; Jiang, X. Stimbiotics Supplementation Promotes Growth Performance by Improving Plasma Immunoglobulin and IGF-1 Levels and Regulating Gut Microbiota Composition in Weaned Piglets. Biology 2023, 12, 441. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Yin, J.; Zhang, K.; Xie, P.; Ding, H.; Huang, X.; Blachier, F.; Kong, X. Dietary xylo-oligosaccharide supplementation alters gut microbial composition and activity in pigs according to age and dose. AMB Express 2019, 9, 134. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| β-Glucan | Animal | Inclusion Rate * | Effect on Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Laminarin BG | Sows | 1 g/day | Improved Lactobacillus spp. numbers in the offspring colon. Reduced offspring fecal counts of Salmonella Typhimurium post-challenge with Salmonella Typhimurium. | Improved offspring ADG and feed efficiency post-weaning. Improved offspring fecal scores. Increased total SCFA in offspring feces post-weaning. | [61] |
| Laminarin BG | Sows | 1 g/day | Increased colonic Lactobacillus spp. gene numbers in offspring at weaning. | Increased villous height in offspring ileum D8 PW. Beneficial effect on immune gene expression markers in offspring pre-weaning and D8 PW. Increased offspring BW on D67 PW and improved G:F ratio from D16 to D117 PW. | [62] |
| Oat BG | Sows | 150,000 g/ton | Significantly higher levels of Bifidobacteria in milk. | The Bifidobacteria isolate from the sow’s milk survived low pH and bile salts exposure when tested. | [63] |
| Yeast BG | Sows | 1,000/2,000 g/ton | Not recorded. | Increased ADFI in both 1000 and 2000 g/ton BG groups compared to control. Increased ADFI in 1000 compared to 2000 g/ton. Weaning weight tended to increase in both groups. Inclusion rate of 1000 g/ton BG decreased TNF-α in sow and piglet serum compared to 2000 g/ton BG. | [64] |
| Yeast BG | Gilts | 300 g/ton | Not recorded. | Increased IgA concentration and proliferation rate of intestinal epithelial cells from colostrum and milk. Did not modulate vaccine response. No effect on reproductive performance. | [65] |
| Oats BG | Suckling pigs | 40 mg/kg bodyweight | No significant changes. | No effect on performance or gut morphology. | [66] |
| Algal BG | Weaned pigs | 54/108 g/ton | Not recorded. | Inclusion rate of 108 g/ton reduced diarrhea frequency, decreased GIT permeability. Both inclusion rates enhanced the mRNA expression of tight junction proteins and boosted immune response against Escherichia coli infection. | [67] |
| Bacterial BG (High and low MW) | Weaned pigs | 50 g/ton | Attenuated the impact of LPS infusion on total bacteria number and copy numbers of Lactobacillus, Bifidobacterium, Bacillus and Escherichia coli in the colonic digesta. Increased Bifidobacterium and Bacillus in the colonic digesta of LPS challenged pigs. High MW BG decreased Escherichia coli in colonic digesta of LPS challenged pigs. | Inhibited LPS-mediated depression in the growth performance, possibly via the Dectin-1 receptor and the TLR4/NF-κB pathway. Different effects were noted between high and low MW BG. Increased butyrate concentration. | [30] |
| Bacterial BG | Weaned pigs | 500 g/ton | Increased abundance of Lactobacillus and Bacillus in the colon after ETEC challenge. | Increased TJP1 in jejunal epithelium following ETEC challenge. Decreased expression of inflammation related proteins in jejunal and ileal mucosa. Increased propanoic acid content in the colon. | [68] |
| Mushroom BG | Weaned pigs | Mushroom powder included at 2,000 g/ton (equivalent of 200 ppm BG) | No significant change. | Reduced feed intake. | [69] |
| Mushroom BG | Weaned pigs | Mushroom powder included at 2,000 g/ton (equivalent of 200 ppm BG) | Decreased relative abundance of Prevotella. | Improved gastrointestinal morphology and upregulated expression of nutrient transporters SLC15A and FABP2 and tight junction protein CLDN1. Reduced ADFI no negative impact the ADG. | [33] |
| Rice Bran BG | Weaned pigs | 1,000/2,000/4,000 g/ton | Linearly decreased coliform bacterial counts in feces with increasing BG inclusion. | Improved ADG and G:F ratio. Linear increase in nutrient digestibility from D0–D42 PW with increasing BG inclusion. | [70] |
| Yeast BG | Suckling/weaned pigs | Oral dose of 50–300 mg every two days. Dose rate started at 50 mg and increased weekly by 50 mg. | Modest effect on fecal microbiota with increases in Methanobrevibacter, Fusobacterium and a genus within the family of Ruminococcaceae. Difficult to state if positive or negative. | Did not affect vaccination response. | [71] |
| Yeast BG | Weaned pigs | 250 g/ton | Increased abundance of pathogenic attaching and effacing Escherichia coli and decreased abundance of Bifidobacterium spp. in cecal digesta. | No effect on performance. Suppressed IL10 expression in ileum. | [60] |
| Yeast BG | Weaned pigs | 100 g/ton | Decreased fecal Escherichia coli. | Increased plasma leucocytes, increased lymphocyte proliferation and decreased TNF-α in the blood plasma at 2 and 4 h post LPS challenge. | [72] |
| Yeast BG | Weaned pigs | 50 g/ton | Not recorded. | Attenuated the increase of IL-6 and TNF-α and enhanced IL-10 when pigs were challenged with LPS. | [50] |
| Laminarin Hyperborean BG | Grower pigs | 250 g/ton | Reduced Enterobacteriaceae population in the ileum and colon. | Downregulated pro-inflammatory (TNF-α, IL-1α, and IL-17A) and anti-inflammatory (IL-10) markers in the colon. | [73] |
| Laminarin digitata BG | Grower pigs | 250 g/ton | Reduced Enterobacteriaceae population in the ileum and colon. | Downregulated pro-inflammatory (TNF-α, IL-1α, and IL-17A) and anti-inflammatory (IL-10) markers in the colon. Reduced total volatile fatty acid concentration in ileum. Ex-vivo model showed an increase in CXCL8 following LPS challenge. | |
| Yeast BG | Grower pigs | 250 g/ton | Reduced Enterobacteriaceae population in the ileum and colon. | Downregulated pro-inflammatory (TNF-α, IL-1α, and IL-17A) and anti-inflammatory (IL-10) markers in the colon. | |
| Yeast BG | Grower pigs | 500 g/ton | When challenged with Salmonella enterica serovar Typhimurium, the supplemented group had reduced shedding counts at D16 post-inoculation. Increase in several potential beneficial microorganisms in feces post-inoculation. | Not recorded. | [74] |
| Bacterial BG | Grower/finisher pigs | 50/100/200 g/ton | Not recorded. | Inclusion rate of 100 g/ton improved ADG, FCR and nutrient digestibility. 100 and 200 g/ton inclusion rate increased carcass length. 100 g/ton improved pork quality. | [75] |
| Mushroom BG | Finisher pigs | Mushroom powder included at 1,000 g/ton (equivalent of 100 mg/kg BG) | Not recorded. | Reduced feed intake. Improved gain to feed ratio. Enhanced the color of fresh pork. | [76] |
| NDO | Animal | Inclusion Rate * | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Chito- oligosaccharides | Sows | 30 g/ton | Not recorded. | Improved amino acid concentration in milk. Improved ADG and weaning weight of offspring. Reduced offspring hypoglycemia by stimulating hepatic gluconeogenesis. | [87] |
| Chito- oligosaccharides | Sows | 30 g/ton | Not recorded. | Increased IgM in colostrum. Increased IgG and IL-10 in serum of offspring at weaning. | [88] |
| Fructo- oligosaccharide | Sows | 1,500 g/ton | Not recorded. | Increased SCFA concentration in offspring feces during lactation and after weaning. Increased cecal goblet cell number and improved ileal cytokine secretions. | [89] |
| Galacto-oligosaccharide | Sows | 10 g/day | Increased the abundance of Alloprevotella and Ruminoclostridium_1 in sow feces and vertically increased fecal Alloprevotella and Ruminoclostridium_1 in offspring feces. | Improved intestinal barriers, immune defense and ADG of offspring. | [90] |
| Mannan-oligosaccharide | Sows | 400 g/ton | Not recorded. | Shortened wean to service interval. Improved growth performance and immunity in offspring. | [91] |
| Mannan-oligosaccharide | Sows | 400 g/ton | No change. | Reduced inflammation marker expression and improved immune competence in offspring. | [92] |
| Chito- oligosaccharides | Weaned pigs | 100/200/400 g/ton | All inclusion rates increased Lactobacillus counts in feces D14 and D21 PW. 200mg/kg COS decreased Escherichia coli counts in feces at D21 PW. | 100 and 200 g/ton increased overall ADG, ADFI and G:F. All inclusion rates decreased incidence of diarrhea and diarrhea scores. 100 g/ton COS increased villus height in ileum and 200 g/ton COS increased villus height and villus height: crypt depth ratio in jejunum and ileum. | [93] |
| Chito- Oligosaccharides * Varying molecular weights | Weaned pigs | 250 g/ton | The use of 5–10 and 10–50 kDa COS increased lactic acid bacteria populations in feces. Using 50–100 kDa COS decreased lactic acid bacteria populations in feces. Using 5–10, 10–50 and 50–100 kDa COS decreased Escherichia coli populations in feces. | Improved ADG and G:F D18-33 PW. Improved fecal scores D0-14 PW. Using 5–10 and 10–50 kDa COS increased nutrient digestibility of diets. | [94] |
| Chito- oligosaccharides | Weaned pigs | 75/150/225 g/ton | Not recorded. | 150 g/ton increased digestibility on D28 and D56 PW, increased villus height and villus height: crypt depth ratio on D28 and increased active cell division on D56 PW. | [95] |
| Fructo- oligosaccharide | Weaned pigs | 40,000 g/ton | No change in fecal bacterial populations. Increase in fecal population of yeast. | Decreased fecal pH and increased organic acid concentration. | [96] |
| Fructo- oligosaccharide | Weaned pigs | 2,500 g/ton | Reduced Escherichia coli and increased populations of Bacillus and Bifidobacterium in cecal digesta. | Improved ADG, apparent digestibility of crude protein, villus height in duodenum and expression of tight junction genes. Increased SCFA in cecal digesta. Decreased diarrhea incidence. | [97] |
| Galacto-oligosaccharide | Weaned pigs | 500/1,000/1,500/2,000 g/ton | Increased the number of Lactobacilli and Bifidobacteria and decreased Escherichia coli in the feces in a linear or quadratic dose-dependent manner. | Improved growth performance in a linear or quadratic dose-dependent manner. Decreased serum pro-inflammatory cytokines in a quadratic dose-dependent manner and increased anti-inflammatory cytokines in a linear or quadratic dose-dependent manner. Promoted serum antioxidant activities in a linear or quadratic dose-dependent manner. | [81] |
| Lactulose | Weaned pigs | 1,000/2,000 g/ton | Both inclusion rates increased Lactobacillus and reduced Escherichia coli fecal counts. | Both inclusion rates increased overall ADG and G:F but did not affect ADFI. Both inclusion rates improved nitrogen digestibility and gross energy. | [98] |
| Mannan-oligosaccharide | Weaned pigs | 1,000 g/ton | Not recorded. | Improved growth and nutrient digestibility. Reduced diarrhea. | [99] |
| Xylo- oligosaccharide 1 | Weaned pigs | 10,000 g/ton | Increased the abundance of Lactobacillus and Bifidobacterium in the ileal digesta. | Reduced diarrhea. Increased ileal villus height and intestinal activity of antioxidizes. Reduced ileal and colonic content of IL-6 and increased colonic sIgA and IL-10 concentrations. | [100] |
| Animal | Inulin Inclusion Rate * | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|
| Sows | 8,000/16,000/24,000 g/ton | Not recorded. | 16,000 g/ton inclusion rate increased litter birth weight, reduced farrowing duration and reduced piglet deaths at birth. 8,000 and 16,000 g/ton inclusion rate increased litter weaning weight, piglet ADG and sow feed intake. Linear improvement in antioxidative status of the sow with increasing inclusion of inulin. | [109] |
| Sows | 30,000 g/ton | Increased Enterococci in feces of sow and caecum of offspring. Deceased Enterobacteria and Lactobacillus amylovorus and increased Eubacteria and Clostridium leptum in offspring stomach digesta. | Decrease in sow fecal pH. Decreased concentrations of ammonia, n-butyric acid and i-valeric acid in the stomach digesta of offspring. | [114] |
| Weaned pigs | 17,000 g/ton | Not recorded. | Improved daily gain and food efficiency D0–D7 PW. | [110] |
| Weaned/grower pigs | 40,000 g/ton (Short-chain/long-chain/50:50 mixture of both) | All 3 types of inulin increased Lactobacilli and Bifidobacteria in the lumen contents in the distal colon. There was a strong effect of inulin on the abundance of Lactobacilli and Bifidobacteria in the mucosal microbiota. These mucosal microbiota alterations were evident as proximal as the jejunum in the short-chain inulin group. However, in the long-chain inulin group changes were not evident until the distal ileum or cecum. | All 3 types of inulin resulted in similar improvements. Improved iron utilization. Increased hemoglobin repletion efficiency. The cecum was the main site of inulin disappearance. | [111,112] |
| Grower pigs | 30,000 g/ton | Did not alter numbers of Lactobacilli, Bifidobacteria, Enterococci, Enterobacteria or bacteria of the Clostridium Coccoides/Eubacterium rectale group in the duodenum, jejunum, or cecum. Reduced Lactobacilli in the stomach. | Reduced cecal acetate. | [113] |
| 50,000 g/ton | Increased β-diversity in colon and cecum. In total, 18 genera altered in the cecum, 17 in the colon and 6 in the ileum. | Increased SCFA in colon and caecum. | [115] | |
| Finisher pigs | 20,000 g/ton | Not recorded. | Improved antioxidant status and water holding capacity of meat, increased intramuscular fat. | [116] |
| Type of Resistant Starch | Animal | Inclusion Rate * | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| RS1—High amylose corn | Sows | 76.5% of total starch | Increased bacterial diversity in the sow feces. | Reduced birthweight but no difference in weaning weight. Increased serum triacylglycerol. Increased non-esterified fatty acids concentration and fat content in milk. | [123] |
| RS2—Field pea starch | Sows | 33% of starch | Increased Firmicutes: Bacteroidetes ratio and abundance of Bifidobacterium in the sow feces. No microbiota differences in offspring. | Milk protein decreased and lactose increased in week 1 of lactation. Increased expression of TJP1 in offspring. No other health benefits observed in offspring. | [124] |
| RS2—Field pea starch | Sows | 33% of starch | No major differences could be distinguished. | Higher SCFA in colon of offspring. | [125] |
| RS2—Raw potato starch | Weaned pigs | 5% | Increased Clostridia in feces. | Increased intestinal concentration of butyrate. Increased T-cell abundance and enhanced mucosal defense status in cecum. | [120] |
| RS2—Raw potato starch | Weaned pigs | 0.5/1% | Not recorded. | Both inclusion rates improved fecal scores, however the 1% group had more solid feces D0–D14 PW. Both inclusion rates decreased ileal and cecal digesta pH. | [121] |
| RS2—Raw potato starch | Weaned pigs | 7/14% | Both inclusion rates increased Lactobacilli and Bacteroides prevalence in the colon. No effect on colon lactic acid bacterial counts. | Increased ileum ammonia N concentrations. Resistant starch content of 7% and 14% improved fecal score D0–7 PW. Resistant starch content of 7% improved fecal score D0–D21 compared to control and 14% RS. | [122] |
| RS3—Retrograded tapioca starch | Grower pigs | Pregelatinized potato starch replaced by 34% retrograded tapioca starch | Not recorded. | Change in eating patterns but no increase in feed intake. Reduction in DE intake but no reduction in ADG. | [126] |
| Pectin Source | Animal | Inclusion Rate * | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Source not stated (Yuzhung Biotech Corporation, China) | Weaned pigs (challenged with LPS) | 50,000 g/ton | Improved α diversity and enriched anti-inflammatory and SCFA-producing bacterial groups in the ileal mucosa. | Ameliorated the LPS-induced inflammation response and damage to ileal morphology. Upregulated expression of MUC2. Increased acetate concentrations. | [128] |
| Citrus peel | Weaned pigs | 50,000 g/ton | Increased Lactococcus and Enterococcus in the jejunum. | Improved intestinal integrity and reduced proinflammatory cytokines. Increased microbiota metabolites skatole, 3-indoleacetic acid, 3-indolepropionic acid, 5-hydroxyindole-3-acetic acid, and tryptamine. Metabolites activated the AhR pathway. | [129] |
| Apple | Weaned pigs | 80,000 g/ton | Increased abundance of Desulfovibrio spp. and Methanobrevibacter spp. in the colon. The abundance of fungal keystone taxa with oxidative phosphorylation was decreased in the colon. | Decreased fecal redox potential. Increased the microbiota-derived antioxidant inosine. | [130] |
| Apple | Weaned pigs | 11,800 g/ton | Not recorded. | Reduced gastric emptying and passage rate through the GIT. Increased digesta water content. Decreased retention time in small intestine. Increased SCFA content. | [131] |
| Apple | Grower pigs | 50,000 g/ton | Reduced diversity at the genera level in the ileal mucosa. Increased abundance of potentially beneficial bacterial populations in the ileal and colonic mucosa. The alterations in the bacterial genera and fermentation metabolites were associated with the differentially expressed genes and cytokine in the ileum and cecum of pigs. | Reduced IL-6, IL-8, IL-12, and IL-18 and tended to reduce IFN-γ in the ileal mucosa. Reduced IL-1β and IFN-γ in the cecal mucosa and tended to reduce IL-8 and IL-1α. Reduced IL-6 in both the ileal and cecal mucosa. Upregulated CLDN2, tended to upregulate expression of MUC2, and downregulated TLR2 and NFKB expression in the ileum. Increased MUC2, TFF3, AMPK and TAK1 expression in the cecal mucosa. Increased sIgA content. Increased SCFA concentration in the cecum. | [132] |
| Amino Acid | Animal | Inclusion Rate * | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Arginine | Sows | 0.25% | Fecal α and β diversities remained the same. Increased both the Bacteroidaceae family and the Bacteroides genera in the feces. | Improved total number of pigs born. Tended to improve total born alive, reduce intrauterine growth restriction and mortality D0–D3. | [165] |
| Glutamate | Weaned pigs | 0.5% | Increased relative composition of bacterial communities of the genus Prevotella and Anaerovibrio and decreased the genus Clostridium and Terrisporobacter. | Increased ADG, ADFI, and nutrient digestibility D0-D14 PW. Increased villus height: crypt depth ratio and number of goblet cells in the duodenum. Tended to increase villus height: crypt depth ratio and number of goblet cells in the ileum. Increased ileal gene expression of claudin family and Occludin. Decreased serum TNF-α and IL-6 and ileal gene expression of TNF. | [166] |
| Glycyl-glutamine | Weaned pigs | 0.25% | Increased α diversity and abundance of beneficial anaerobes and fiber-degrading bacteria in the feces (stool from rectum). | Increased SCFA in the ileum and colon. Improved BW on D10 and D21 PW. Increased ADFI and ADG D0–D10 and D10–D21 PW. Reduced diarrhea ratio. | [167] |
| Tryptophan | Weaned pigs | 0.2/0.4% | Increased α and β diversities and enriched abundances of Prevotella, Roseburia and Sussinivibriogenera in the cecum. Decreased Clostridium sensustricto and Clostridium XI in the caecum. | Inclusion rates of 0.2 and 0.4% increased the ADFI and ADG D0–D14 PW. Inclusion rates of 0.2 and 0.4% increased isobutyrate, isovalerate and indoleacetic acid in the colonic contents. An inclusion rate of 0.2% increased propionate in the colonic contents and indole in the cecal and colonic contents. | [152] |
| Tryptophan | Weaned pigs | 0.1/0.2/0.4% | Inclusion rates of 0.2 and 0.4% enhanced Chao1 α diversity, reduced the abundances of Clostridium sensustricto and Streptococcus and increased the abundances of Lactobacillus and Clostridium XI in the jejunum. | Concentration of tryptophan in the serum increased in a dose dependent manner. Inclusion rates of 0.2 and 0.4% increased the abundances of ZO-1 and ZO-3, and the presence of claudin-1 proteins in the jejunum of weaned pigs was enhanced. An inclusion rate of 0.4% increased abundance of Occludin in the jejunum. An inclusion rate of 0.2% increased ZO-1 in the duodenum. An inclusion rate of 0.2% increased sIgA in the jejunum. Inclusion rates of 0.2 and 0.4% increased expression of porcine β-defensin genes in the jejunum. | [153] |
| Valine * In a low-protein diet | Weaned pigs | 0.48% added to low-protein diet. (% SID valine: SID Lysine 0.12 above NRC recommended level) | Increased abundance Mogibacterium in colon. | Increased ADFI. | [146] |
| Valine + isoleucine * In a low-protein diet | Weaned pigs | 0.48% valine and 0.33% isoleucine added. (% SID valine: SID lysine 0.12 above NRC recommended level, isoleucine equals NRC recommended level) | Increased abundance of Actinobacteria, Enterococcus, and Brevibacillus in colon. | Increased ADFI. Tended to increase final BW. Tended to increase ADG. | [146] |
| Citrulline | Finisher pigs | 1% | Increased α diversity and microbiota composition of the feces. In particular, the altered gut microbiota at the phylum and genus level may be mainly involved in metabolic process of carbohydrate, energy, and amino acid, and exhibited a significant association with final weight, carcass weight and backfat thickness. | Drastically increased final BW, liveweight gain, carcass weight and average backfat. Decreased drip loss. | [168] |
| Glutamate ± arginine | Finisher pigs | 1% glutamate ± 1% arginine | Glutamate in combination with arginine increased the abundance of Actinobacteria in the colon. | Glutamate alone or in combination with arginine decreased bodyfat weight and increased SCFA concentration in the colon. | [169] |
| Leucine | Finisher pigs | 1% leucine | Increased the abundance of Actinobacteria in the colon. | Decreased body fat weight and increased colonic SCFA production. | [170] |
| Leucine and arginine | 1% leucine + 1% arginine | Reduced abundance of Bacteroidetes and increased abundance of Proteobacteria in the colon. Increased Clostridium_sensu_stricto_1, Terrisporobacter, and Escherichia-Shigella. | Decreased body fat weight and increased colonic SCFA production. | ||
| Leucine and glutamate | 1% leucine + 1% glutamate | Increased propanoate concentration in colon. | No effect on performance or meat quality parameters. |
| Nucleotide | Animal | Inclusion Rate * | Effect on Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Yeast nucleotides (SwineMOD®) | Suckling pigs | Oral dose of 100mg on D10, D15, D18 and D21 of life. | Increased abundance of Campylobacteraceae, and decreased abundance of Streptococcaceae in the feces at weaning but not at D12 PW. | A time- and tissue-dependent effect. Reduced inflammatory activation at weaning and increased erythropoietic activity post-weaning in blood transcriptome PW. | [174] |
| Equal ratios of CMP, UMP, AMP, GMP, IMP | Weaned pigs | 350 g/ton | Reduced the Firmicutes/Bacteroidetes ratio in the colon. At genus level, enriched the relative abundance of Prevotella, Faecalibacterium and Olsenella. | Decrease the diarrhea rate. Increased villus height and the villus height: crypt depth ratio in the ileum. Upregulated protein expression of tight junction proteins and the mRNA expression of MUC2 while the mRNA expression of MUC4 was downregulated in the ileal mucosa. Increased the ileal mucosa genes expression of IL21, INFG, IL10, IL4, IL6 and TNF and increased the protein expression of NF-κB, IL-6 and TNF-α. Increased short chain fatty acid in the colon. | [176] |
| Yeast nucleotides (Maxi-Gen®) | Weaned pigs | 1000 g/ton | Clean room: suppressed growth of cecal Enterobacteriaceae members and colonic Enterococcus spp, improved proliferation of cecal Lactobacillus spp. and colonic Clostridium cluster IV and XVIa members. Unclean room: improved proliferation of cecal Clostridium cluster IV and suppressed proliferation of colonic Enterococcus spp. | No effect on growth performance in clean or unclean conditions. Clean room: tended to improve the villus height: crypt depth ratio. Unclean room: upregulation of ileal PDCD1, IL1B, IL6, IL10 and TNF. | [177] |
| 45.1 mg AMP, 22.4 mg CMP, 65.8 mg GMP, 9.5 mg IMP, and 1202.0 mg UMP | Weaned pigs | Oral dose of 1.34 g per day | Increased bacterial numbers of Enterococcus spp. in the cecal digesta. No difference in jejunum, cecum, or feces for numbers of total bacteria, Lactobacillus group, Enterobacteriaceae, Bifidobacteria spp., Clostridium Cluster XIV, or Clostridium Cluster IV. | Increased ADFI. Increased plasma IgA concentrations. No change in gut morphology. | [178] |
| Polyphenol Source | Animal | Inclusion Rate * | Effect on Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Holly | Weaned pigs | 250 g/ton | Increased the abundance of Firmicutes and reduced Bacteroidetes in the colon. Increased relative abundance of Prevotella_9 in the caecum and Lactobacillus both in the caecum and colon of LPS challenged pigs. | Decreased villus height: crypt depth ratio. Increased jejunal lactase activity of LPS challenged pigs. Higher activity of sucrase and lactase in jejunum and sucrase in ileum. Reduced concentration of TNF-α, IL-6 and insulin in plasma. Increased glucagon concentration in plasma. Increased the mRNA expression of tight junction proteins. Increased the concentrations of cecal valerate and colonic acetate and isovalerate in LPS challenged piglets. | [181] |
| Citrus flavonoids | Weaned pigs | 0.3 g/ton | Increased the abundance of several genera of bacteria such as Lactobacillus, Roseburia, and Clostridium in the cecum. Decreased the relative abundance of Dorea, Desulfovibrio, and Actinobacillus, among others, in the cecum. | Increased BW and ADG during the starter and entire period. Increased the expression of genes related to barrier function, digestive enzymes, and nutrient transport. | [191] |
| Grape pomace | Grower pigs | 50,000 g/ton | Decreased Lactobacillus and Ruminicoccus and increased Treponema and Campylobacter in the colon. | Increased numbers of eosinophils induced by Ascaris suum infection in the duodenum, jejunum and ileum, and modulated gene expression in the jejunal mucosa of infected pigs. | [192] |
| Flavonoid enriched cocoa powder | Finisher pigs 1 | 0/2.5/10/20 g/d (flavanol concentration of 20.5 mg/g) | Rates of 10 and 20 g/d increased the abundance of Lactobacillus in the species and Bifidobacterium in the proximal colon. | No effect on bodyweights. Expression of TNF and TLR2, TLR4, and TLR9 was reduced in the ileal Peyer’s patches, mesenteric lymph nodes and proximal colon. | [185] |
| Grape pomace (Anta®Ox E) or spent hops (Anta®Phyt H) | Weaned/ grower pigs | 10,000 g/ton | Lower counts of Streptococcus spp. and Clostridium Cluster XIVa in the faecal microbiota. | Both showed an improved gain: feed ratio in comparison to the control group Both increased fecal pH value and lowered levels of volatile fatty acids. | [186] |
| Red-osier dogwood polyphenol | Finisher pigs | 5,000 g/ton | Increased α-diversity, class Bacilli, order Lactobacillales and family Lactobacillaceae in ileal digesta. Within family Lactobacillaceae, Lactobacillus was the main responder by increasing from 5.92% to 35.09% in ileal digesta. | No effect on performance. Increased propionate in ileal digesta. | [193] |
| Stimbiotic | Inclusion Rate * | Animal | Effect on GIT Microbiota | Other Effects | Reference |
|---|---|---|---|---|---|
| Xylo- oligosaccharide | 100 g/ton | Weaned pigs | Enhanced α-diversity, reduced Lactobacillus, and increased Streptococcus and Turicibacter at the genus level in the distal gut digesta. | Reduction in the inflammatory marker IFN-γ and improved intestinal barrier function through up regulation of TJP1. Little effect on growth performance, intestinal morphology, blood cells and biochemical markers. | [197] |
| Xylo- oligosaccharide | 200 g/ton | Weaned pigs | Decreased Escherichia coli and increased Lactobacilli fecal shedding on D14 PW but not on D28 PW. | Improved ADG, FCR and digestibility. Increased villus height: crypt depth ratio in jejunum. | [31] |
| Xylo- oligosaccharide | 400 g/ton | Weaned pigs | Not recorded. | Improved ADG and weight D28 PW. Increased serum glucose content. Decreased blood urea nitrogen and triglyceride. Increased serum IgG. Improved antioxidant and immune function of pigs. | [198] |
| Xylo- Oligosaccharide + β-1,4-endo xylanase (VistaPros®) | 100 g/ton | Weaned/grower pigs | Increased relative abundance of beneficial bacteria norank_f_Muribaculaceae, Rikenellaceae_RC9_gut_group, Parabacteroides, and unclassified_f__Oscillospiraceae in the feces on D42 PW. | Improved BW of piglets on D28 and D42 PW and increased ADG and ADFI from D14–28 PW and from D0–D42 PW. Increased plasma insulin-like growth factor on D42 PW. | [199] |
| Xylo- Oligosaccharide + β-1,4-endo xylanase (Signis®) | 100 g/ton | Weaned/grower pigs | Increased proportion of Clostridiales Family XIII Incertae Sedis and Clostridiaceae (Families contain beneficial fibrolytic and butyrate-producing bacteria) in the feces on D35 PW. | Increased weight at D42 PW under poor sanitary conditions (prebiotic comparisons included at 10- and 20-times higher doses did not increase bodyweight). Increased VFA:BCFA higher than prebiotics. Reduced plasma TNF-α content in pigs raised under poor sanitary conditions. | [194] |
| Xylo- Oligosaccharide (XOS) + β-1,4-endo xylanase (XYL) | XYL: 150 g/ton XOS: 200 g/ton XYL + XOS: 150 g/ton + 200 g/ton | Grower pigs | Limited effects. No effect on α or β diversity. Operational taxonomic units associated with Muribaculaceae_ge and Prevotellaceae NK3B31 group were higher in the feces of pigs from all 3 diets. | XYL: no effect on performance.. XOS: limited effect on growth performance. Reduced G:F D0-D7 of trial. Improved ADG D7-D14 of trial. No overall affect (D0-D35). XYL + XOS: no interaction. | [196] |
| Xylo- oligosaccharide | 100/250/500 1 g/ton | Grower/finisher pigs | 100 g/ton decreased presumed pathogenic bacteria, Proteobacteria and Citrobacter, and increased likely beneficial bacteria, Firmicutes and Lactobacillus, abundance in the colonic contents of pigs in the grower to finisher phase (30–100 kg). The dose and exposure time to XOS affected colonic microbial communities. | 100 g/ton increased acetic acid and total SCFA concentration in the intestinal contents. | [200] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiernan, D.P.; O’Doherty, J.V.; Sweeney, T. The Effect of Prebiotic Supplements on the Gastrointestinal Microbiota and Associated Health Parameters in Pigs. Animals 2023, 13, 3012. https://doi.org/10.3390/ani13193012
Kiernan DP, O’Doherty JV, Sweeney T. The Effect of Prebiotic Supplements on the Gastrointestinal Microbiota and Associated Health Parameters in Pigs. Animals. 2023; 13(19):3012. https://doi.org/10.3390/ani13193012
Chicago/Turabian StyleKiernan, Dillon P., John V. O’Doherty, and Torres Sweeney. 2023. "The Effect of Prebiotic Supplements on the Gastrointestinal Microbiota and Associated Health Parameters in Pigs" Animals 13, no. 19: 3012. https://doi.org/10.3390/ani13193012
APA StyleKiernan, D. P., O’Doherty, J. V., & Sweeney, T. (2023). The Effect of Prebiotic Supplements on the Gastrointestinal Microbiota and Associated Health Parameters in Pigs. Animals, 13(19), 3012. https://doi.org/10.3390/ani13193012