
Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Organism
2.2. RNA Extraction and Illumina HiSeq Sequencing
2.3. Transcriptomic Assembly and Functional Annotation
2.4. Differentially Expressed Gene (DEG) Comparison and Function Enrichment
2.5. Histological Analysis
2.6. Validation of Transcriptomic Data and Glucose Metabolism-Related Gene Expression by qRT-PCR
2.7. Determination of Glycometabolic Enzyme Activities
2.8. Statistical Analysis
3. Results
3.1. Basic Growth Index
3.2. Transcriptome Sequencing and Assembly
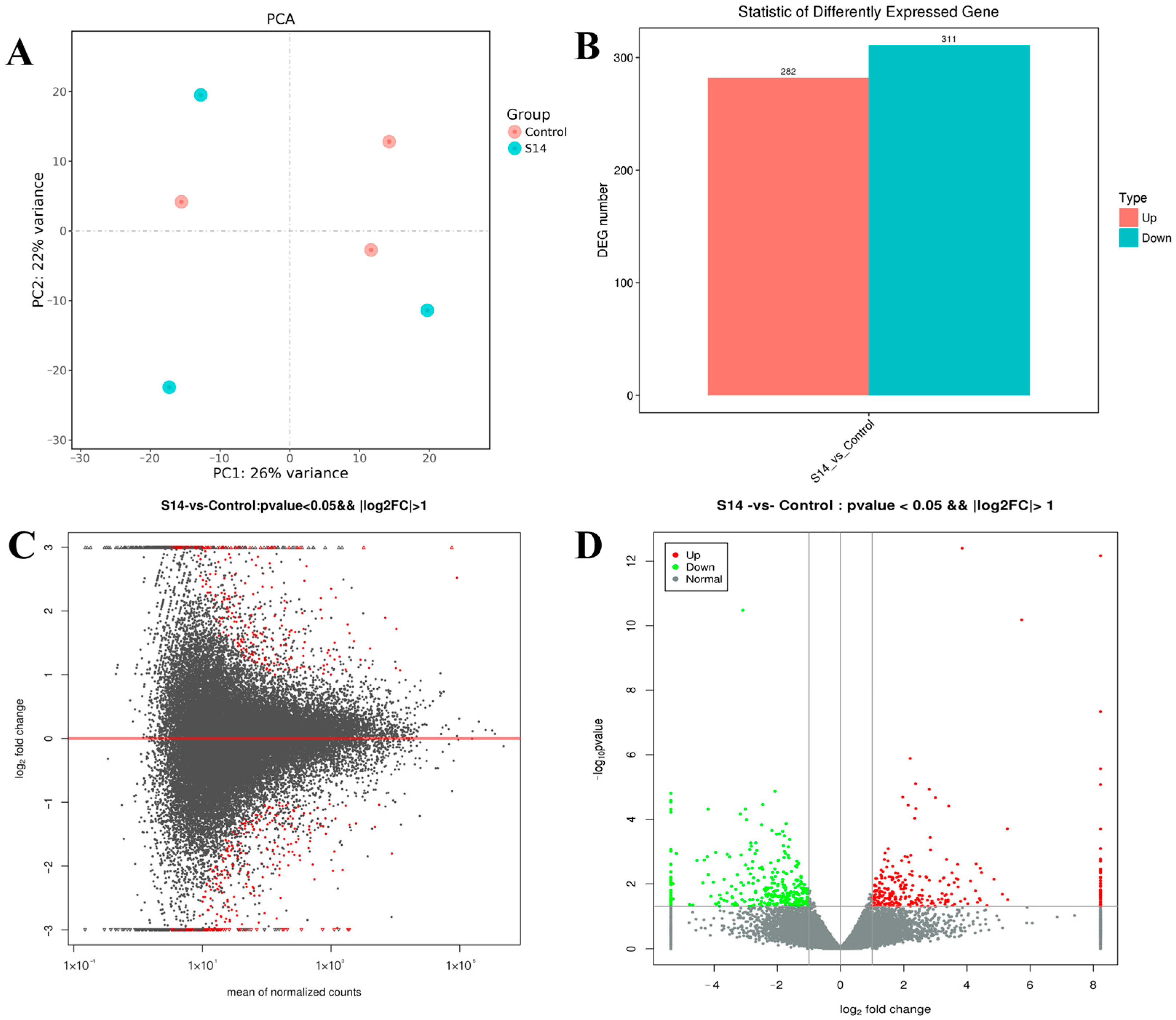
3.3. Analysis of Differentially Expressed Genes (DEGs)
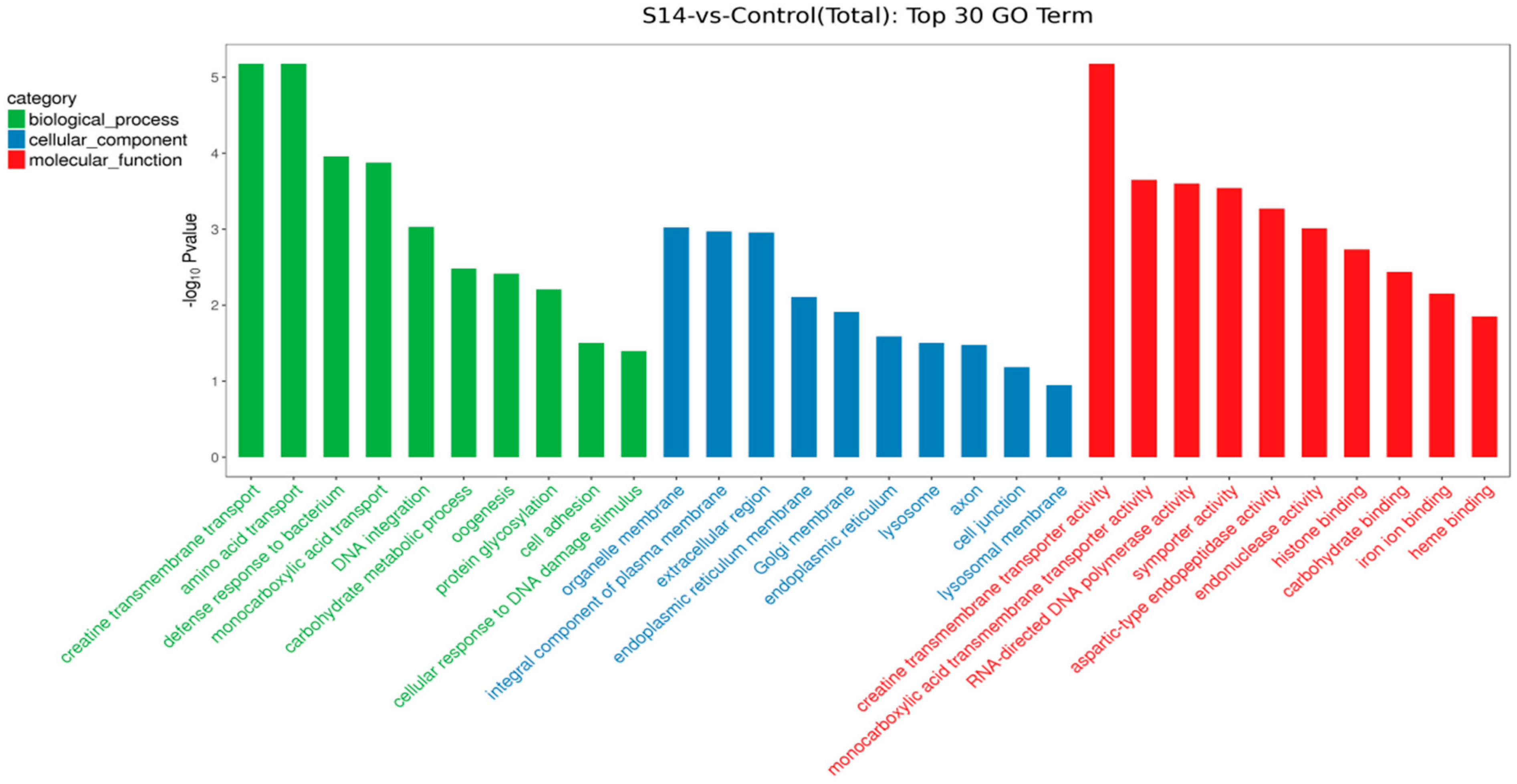
3.4. GO and KEGG Enrichment Analysis
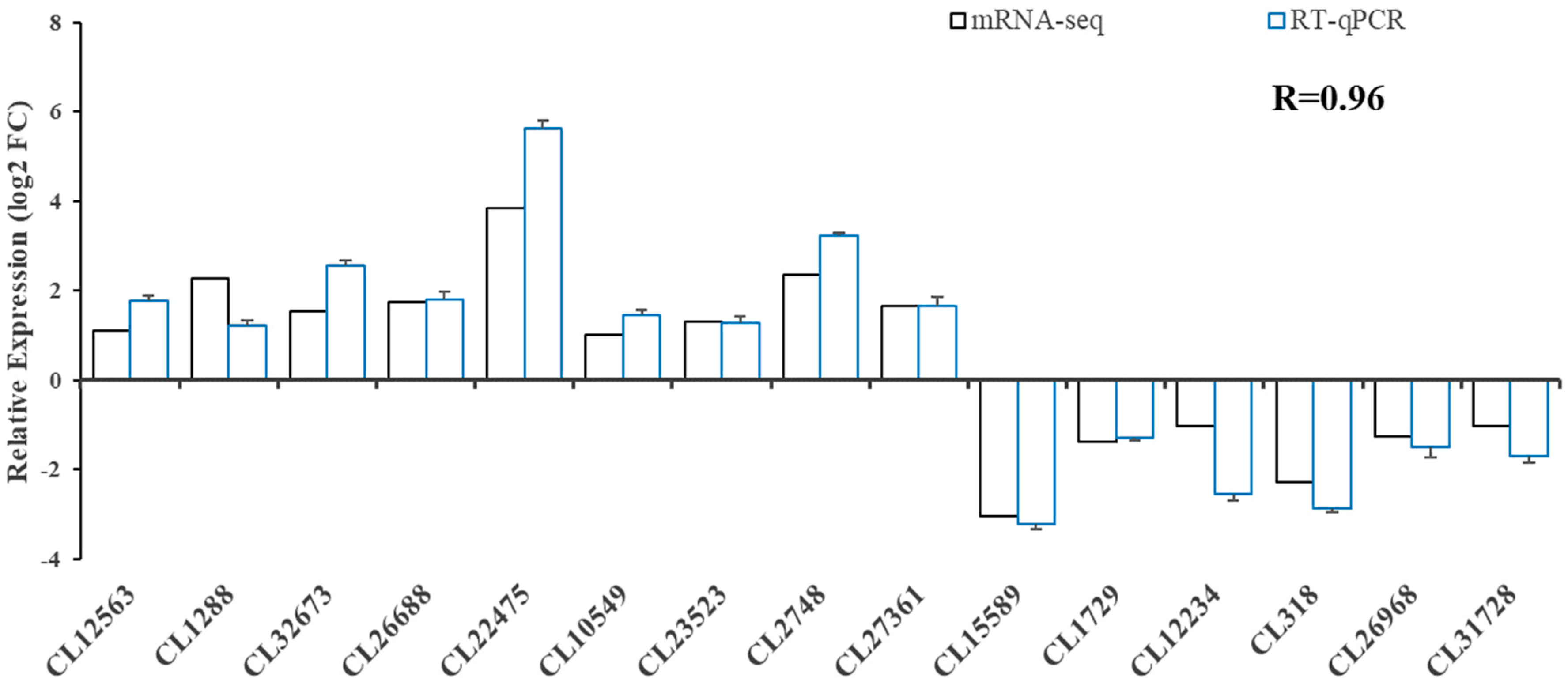
3.5. Validation of DEG and Glucose Metabolism-Related Gene Analysis
3.6. Histological Analysis of Gills
3.7. Glucose Metabolism-Related Enzyme Activity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhuyan, I.; Mia, S.; Supit, I.; Ludwig, F. Spatio-temporal variability in soil and water salinity in the south-central coast of Bangladesh. CATENA 2023, 222, 106786. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, C.; Hu, Y.; Shao, K.; Tang, X.; Zhang, L.; Gao, G.; Qin, B. Climate-induced salinization may lead to increased lake nitrogen retention. Water Res. 2023, 228 Pt A, 119354. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Z.; Li, Z.; Yang, B.; Li, B.; Tang, X.; Lai, Y. Comprehensive study on saline-alkali soil amelioration with sediment of irrigation area in northeast China. Arab. J. Chem. 2023, 16, 104608. [Google Scholar] [CrossRef]
- Re, A.D.; Díaz, F.; Ponce-Rivas, E.; Giffard, I.; Muñoz-Marquez, M.; Sigala-Andrade, H.M. Combined effect of temperature and salinity on the Thermotolerance and osmotic pressure of juvenile white shrimp Litopenaeus vannamei (Boone). J. Therm. Biol. 2012, 37, 413–418. [Google Scholar] [CrossRef]
- Kovács, Z.; Samhaber, W. Contribution of pH dependent osmotic pressure to amino acid transport through nanofiltration membranes. Sep. Purif. Technol. 2008, 61, 243–248. [Google Scholar] [CrossRef]
- Yang, Z.; Zhu, L.; Zhao, X.; Cheng, Y. Effects of salinity stress on osmotic pressure, free amino acids, and immune-associated parameters of the juvenile Chinese mitten crab, Eriocheir sinensis. Aquaculture 2022, 549, 737776. [Google Scholar] [CrossRef]
- Kumari, A.; Koner, D.; Lanong, A.S.; Khongmawloh, E.; Snaitang, R.; Saha, N. Changes in the expression of certain osmosensitive channel and transporter genes in primary hepatocytes of air-breathing catfish, Clarias magur: A strategy to adapt under osmotic stress. Aquaculture 2023, 562, 738756. [Google Scholar] [CrossRef]
- Wu, D.-L.; Wang, T.-S.; Liu, H.-J.; Zhang, W.; Tong, X.-H.; Peng, D.-Y.; Kong, L.-Y. Study on the mechanism of Wuzi-Yanzong-Wan-medicated serum interfering with the mitochondrial permeability transition pore in the GC-2 cell induced by atractyloside. Chin. J. Nat. Med. 2022, 20, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Qi, D.; Li, S.; Zhai, M.; Li, Z.; Guo, X.; Ruan, M.; Yu, X.; Zhao, P.; Zhang, P.; et al. The cassava (Manihot-esculenta Crantz)’s nitrate transporter NPF4.5, expressed in seedling roots, involved in nitrate flux and osmotic stress. Plant Physiol. Biochem. 2023, 194, 122–133. [Google Scholar] [CrossRef]
- Wen, Z.; Li, Y.; Bian, C.; Shi, Q.; Li, Y. Genome-wide identification of a novel elovl4 gene and its transcription in response to nutritional and osmotic regulations in rabbitfish (Siganus canaliculatus). Aquaculture 2020, 529, 735666. [Google Scholar] [CrossRef]
- Oron, G.; Appelbaum, S.; Guy, O. Reuse of brine from inland desalination plants with duckweed, fish and halophytes toward increased food production and improved environmental control. Desalination 2023, 549, 116317. [Google Scholar] [CrossRef]
- Bhagwati, A.; Shah, M.; Prajapati, M. Emerging technologies to sustainability: A comprehensive study on solar desalination for sustainable development. Sustain. Manuf. Serv. Econ. 2023, 5, 100007. [Google Scholar] [CrossRef]
- Heng, T.; He, X.-L.; Yang, L.-L.; Xu, X.; Feng, Y. Mechanism of Saline–Alkali land improvement using subsurface pipe and vertical well drainage measures and its response to agricultural soil ecosystem. Environ. Pollut. 2022, 293, 118583. [Google Scholar] [CrossRef]
- Li, Z.-H.; Xing, S.; Li, P.; He, S.; Cao, Z.; Wang, X.; Cao, X.; Liu, B.; You, H. Systematic toxicological analysis of the effect of salinity on the physiological stress induced by triphenyltin in Nile tilapia. Aquat. Toxicol. 2023, 257, 106441. [Google Scholar] [CrossRef] [PubMed]
- Rahmah, S.; Liew, H.J.; Napi, N.; Rahmat, S.A. Metabolic cost of acute and chronic salinity response of hybrid red tilapia Oreochromis sp. larvae. Aquac. Rep. 2020, 16, 100233. [Google Scholar] [CrossRef]
- Qin, H.; Zhu, Z.X.; Lin, H.R.; Xia, J.H.; Jia, Y.X. Exploring candidate genes in a major QTL region associated with salinity tolerance in the skin of Nile tilapia based on transcriptomic analysis. Aquaculture 2020, 526, 735380. [Google Scholar] [CrossRef]
- Zou, H.; He, F.; Lan, Z.; Tang, L.; Lu, W. The personality of Japanese flounder (Paralichthys olivaceus) and gene expression related with osmoregulatory capacity in the gills. Aquaculture 2019, 500, 221–227. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W. RNA-seq resolving host-pathogen interactions: Advances and applications. Ecol. Genet. Genom. 2020, 15, 100057. [Google Scholar] [CrossRef]
- Li, J.; Ma, P.; Liu, P.; Chen, P.; Li, J. The roles of Na+/K+-ATPase α-subunit gene from the ridgetail white prawn Exopalaemon carinicauda in response to salinity stresses. Fish Shellfish Immunol. 2015, 42, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Qi, T.; Liu, J.; Liu, Q.; Jiang, S.; Zhang, H.; Wang, Z.; Ding, G.; Tang, B. Adaptively differential expression analysis in gill of Chinese mitten crabs (Eriocheir japonica sinensis) associated with salinity changes. Int. J. Biol. Macromol. 2018, 120 Pt B, 2242–2246. [Google Scholar] [CrossRef]
- Yang, L.; Li, X.; Jiang, S.; Qiu, L.; Zhou, F.; Liu, W.; Jiang, S. Characterization of Argonaute2 gene from black tiger shrimp (Penaeus monodon) and its responses to immune challenges. Fish Shellfish Immunol. 2014, 36, 261–269. [Google Scholar] [CrossRef]
- Luo, Z.; Yu, Y.; Zhang, Q.; Bao, Z.; Xiang, J.; Li, F. Comparative Transcriptome Analysis Reveals the Adaptation Mechanism to High Salinity in Litopenaeus vannamei. Front. Mar. Sci. 2022, 9, 864338. [Google Scholar] [CrossRef]
- Farhadi, A.; Liu, Y.; Xu, C.; Wang, X.; Li, E. The role of the renin-angiotensin system (RAS) in salinity adaptation in Pacific white shrimp (Litopenaeus vannamei). Front. Endocrinol. 2022, 13, 1089419. [Google Scholar] [CrossRef]
- Ali, A.; Azom, G.; Sarker, B.S.; Rani, H.; Alam, S.; Islam, M.S. Repercussion of salinity on hematological parameters and tissue morphology of gill and kidney at early life of tilapia. Aquac. Fish. 2022, 4, 1016. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Ding, Q.; Teame, T.; Yang, Y.; Ran, C.; Zhang, Z.; Zhou, Z. Research advances in the structure, function, and regulation of the gill barrier in teleost fish. Water Biol. Secur. 2023, 2, 100139. [Google Scholar] [CrossRef]
- Hu, Z.; Xu, L.; Song, H.; Feng, J.; Zhou, C.; Yang, M.-J.; Shi, P.; Li, Y.-R.; Guo, Y.-J.; Li, H.-Z.; et al. Effect of heat and hypoxia stress on mitochondrion and energy metabolism in the gill of hard clam. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 266, 109556. [Google Scholar] [CrossRef]
- Chao, D.H.M.; Kirchner, M.K.; Pham, C.; Foppen, E.; Denis, R.G.; Castel, J.; Morel, C.; Montalban, E.; Hassouna, R.; Bui, L.-C.; et al. Hypothalamic astrocytes control systemic glucose metabolism and energy balance. Cell Metab. 2022, 34, 1532–1547.e6. [Google Scholar] [CrossRef]
- Nain-Perez, A.; Nilsson, O.; Lulla, A.; Håversen, L.; Brear, P.; Liljenberg, S.; Hyvönen, M.; Borén, J.; Grøtli, M. Tuning liver pyruvate kinase activity up or down with a new class of allosteric modulators. Eur. J. Med. Chem. 2023, 250, 115177. [Google Scholar] [CrossRef]
- Jiao, N.; Ji, W.S.; Zhang, B.; Shang, Y.K.; Zhang, Y.C.; Yu, W.Q.; Jin, H.L.; Li, C.; Zhang, C.Y.; Yan, C.; et al. Overexpression of Protein Phosphatase 2 Regulatory Subunit B″ Alpha Promotes Glycolysis by Regulating Hexokinase 1 in Hepatocellular Carcinoma. Biomed. Environ. Sci. 2022, 35, 622–632. [Google Scholar] [CrossRef]
- Fulghum, K.L.; Audam, T.N.; Lorkiewicz, P.K.; Zheng, Y.; Merchant, M.; Cummins, T.D.; Dean, W.L.; Cassel, T.A.; Fan, T.W.; Hill, B.G. In vivo deep network tracing reveals phosphofructokinase-mediated coordination of biosynthetic pathway activity in the myocardium. J. Mol. Cell. Cardiol. 2022, 162, 32–42. [Google Scholar] [CrossRef]
- Iacovino, L.G.; Rossi, M.; Di Stefano, G.; Rossi, V.; Binda, C.; Brigotti, M.; Tomaselli, F.; Pasti, A.P.; Piaz, F.D.; Cerini, S.; et al. Allosteric transitions of rabbit skeletal muscle lactate dehydrogenase induced by pH-dependent dissociation of the tetrameric enzyme. Biochimie 2022, 199, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Feng, J.; Lin, J.; Li, J. The complete mitochondrial genome of Macrobrachium nipponense. Gene 2011, 487, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Wang, L.; Guan, T.; Chang, G.; Zhao, H.; Wu, N. Modeling the relationships of growth and morphometrics to dress-out traits in the oriental river prawn, Macrobrachium nipponense. Aquac. Rep. 2022, 26, 101295. [Google Scholar] [CrossRef]
- Zhi, L.; Zihong, Z.; Yudi, H.; Piao, C.; Xianmei, G.; Bin, L.; Chao, T.; Shuannan, X.; Wenjing, Z.; Wenjing, H.; et al. Origin and farming pattern authentication of wild-caught, coast-pond and freshwater farming white shrimp (Litopenaeus vannamei) in Chinese market using multi-stable isotope analysis of tail shell. Food Control 2023, 148, 109646. [Google Scholar] [CrossRef]
- Kaleem, O.; Sabi, A.-F.B.S. Overview of aquaculture systems in Egypt and Nigeria, prospects, potentials, and constraints. Aquac. Fish. 2021, 6, 535–547. [Google Scholar] [CrossRef]
- Sun, S.; Yang, M.; Fu, H.; Ge, X.; Zou, J. Altered intestinal microbiota induced by chronic hypoxia drives the effects on lipid metabolism and the immune response of oriental river prawn Macrobrachium nipponense. Aquaculture 2020, 526, 735431. [Google Scholar] [CrossRef]
- Ding, Z.; Zhou, J.; Kong, Y.; Zhang, Y.; Cao, F.; Luo, N.; Ye, J. Dietary arachidonic acid promotes growth, improves immunity, and regulates the expression of immune-related signaling molecules in Macrobrachium nipponense (De Haan). Aquaculture 2018, 484, 112–119. [Google Scholar] [CrossRef]
- Li, T.; Wu, L.; Jin, M.; Ma, F.; Huang, X.; Ren, Q. Function of two ficolin-like proteins in innate immune defense of the oriental river prawn, Macrobrachium nipponense. Fish Shellfish Immunol. 2017, 68, 488–499. [Google Scholar] [CrossRef]
- Wiszniewski, G.; Jarmołowicz, S.; Hassaan, M.S.; Kamaszewski, M.; Szudrowicz, H.; Terech-Majewska, E.; Kawalski, K.; Martynow, J.; Szczepański, A.; Siwicki, A.K. Dietary effect of actinidin enzyme on growth, digestive enzymes activity, immunity, liver and intestine histology of juvenile sterlet sturgeon (Acipenser ruthenus). Aquac. Rep. 2022, 25, 101196. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, M.; Li, Y.; Wu, D.; Liu, Z.; Jiang, Q.; Zhao, Y. Effects of salinity acclimation on the growth performance, osmoregulation and energy metabolism of the oriental river prawn, Macrobrachium nipponense (De Haan). Aquac. Res. 2018, 50, 685–693. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, N.; Zhang, Y.; Wang, G.; Pu, S.; Guan, T.; Zhu, C.; Wang, H.; Li, J. Effects of copper on non-specific immunity and antioxidant in the oriental river prawn (Macrobrachium nipponense). Ecotoxicol. Environ. Saf. 2022, 236, 113465. [Google Scholar] [CrossRef]
- Fan, Y.; Feng, J.; Wang, Z.; Ling, F.; Ma, K.; Hua, X.; Li, J.; Li, J. Integrated transcriptomic and metabolic analysis response in gills, hepatopancreas, and muscle metabolism in oriental river prawn Macrobrachium nipponense in response to acute high salinity stress. Aquac. Rep. 2022, 27, 101358. [Google Scholar] [CrossRef]
- Zikos, A.; Seale, A.P.; Lerner, D.T.; Grau, E.G.; Korsmeyer, K.E. Effects of salinity on metabolic rate and branchial expression of genes involved in ion transport and metabolism in Mozambique tilapia (Oreochromis mossambicus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 178, 121–131. [Google Scholar] [CrossRef]
- Hongtuo, F.; Sufei, J.; Yiwei, X. Current status and prospects of farming the giant river prawn (Macrobrachium rosenbergii) and the oriental river prawn (Macrobrachium nipponense) in China. Aquac. Res. 2012, 43, 993–998. [Google Scholar] [CrossRef]
- Farhadi, A.; Liu, Y.; Xu, C.; Han, T.; Wang, X.; Li, E. Evidence from transcriptome analysis unravelled the roles of eyestalk in salinity adaptation in Pacific white shrimp (Litopenaeus vannamei). Gen. Comp. Endocrinol. 2022, 329, 114120. [Google Scholar] [CrossRef]
- Maraschi, A.C.; Faria, S.C.; McNamara, J.C. Salt transport by the gill Na+ -K+ -2Cl− symporter in palaemonid shrimps: Exploring physiological, molecular and evolutionary landscapes. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 257, 110968. [Google Scholar] [CrossRef]
- Marques, B.L.; Oliveira-Lima, O.C.; Carvalho, G.A.; de Almeida Chiarelli, R.; Ribeiro, R.I.; Parreira, R.C.; da Madeira Freitas, E.M.; Resende, R.R.; Klempin, F.; Ulrich, H.; et al. Neurobiology of glycine transporters: From molecules to behavior. Neurosci. Biobehav. Rev. 2020, 118, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Olkhova, E.; Raba, M.; Bracher, S.; Hilger, D.; Jung, H. Homology Model of the Na+/Proline Transporter PutP of Escherichia coli and Its Functional Implications. J. Mol. Biol. 2011, 406, 59–74. [Google Scholar] [CrossRef]
- Selcer, K.; Balasubramonian, B.; Miller, D.; Kerr, J.; DiFrancesco, M.; Ojha, S.; Urbano, R. Steroid sulfatase in the mouse NIH-3T3 fibroblast cell line: Characterization, and downregulation by glucocorticoids. Steroids 2021, 174, 108890. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Jeanson, Y.; Barreau, C.; Moro, C.; Peyriga, L.; Cahoreau, E.; Guissard, C.; Arnaud, E.; Galinier, A.; Bouzier-Sore, A.-K.; et al. Lactate fluxes mediated by the monocarboxylate transporter-1 are key determinants of the metabolic activity of beige adipocytes. J. Biol. Chem. 2021, 296, 100137. [Google Scholar] [CrossRef] [PubMed]
- Järvå, M.A.; Dramicanin, M.; Lingford, J.P.; Mao, R.; John, A.; Jarman, K.E.; Grinter, R.; Goddard-Borger, E.D. Structural basis of substrate recognition and catalysis by fucosyltransferase 8. J. Biol. Chem. 2020, 295, 6677–6688. [Google Scholar] [CrossRef]
- Wei, X.; Fu, X.; Xiao, M.; Liu, Z.; Zhang, L.; Mou, H. Dietary galactosyl and mannosyl carbohydrates: In-vitro assessment of prebiotic effects. Food Chem. 2020, 329, 127179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Niu, J.; Li, Q.; Huang, Y.; Jiang, B.; Li, X.; Jian, J.; Huang, Y. A novel C-type lectin (CLEC12B) from Nile tilapia (Oreochromis niloticus) is involved in host defense against bacterial infection. Fish Shellfish Immunol. 2022, 131, 218–228. [Google Scholar] [CrossRef]
- Zhao, C.; Peng, C.; Fan, S.; Wang, P.; Yan, L.; Xie, Z.; Qiu, L. Identification of a novel crustacyanin-like lipocalin in Penaeus monodon: Molecular cloning, tissue distribution and its functional studies in astaxanthin accumulation. Aquaculture 2021, 539, 736615. [Google Scholar] [CrossRef]
- Maravilla, E.; Le, D.P.; Tran, J.J.; Chiu, M.H.; Prenner, E.J.; Weers, P.M. Apolipophorin III interaction with phosphatidylglycerol and lipopolysaccharide: A potential mechanism for antimicrobial activity. Chem. Phys. Lipids 2020, 229, 104909. [Google Scholar] [CrossRef]
- Grabner, G.F.; Zimmermann, R.; Schicho, R.; Taschler, U. Monoglyceride lipase as a drug target: At the crossroads of arachidonic acid metabolism and endocannabinoid signaling. Pharmacol. Ther. 2017, 175, 35–46. [Google Scholar] [CrossRef]
- David, E.; Boudry, P.; Dégremont, L.; Tanguy, A.; Quéré, N.; Samain, J.; Moraga, D. Genetic polymorphism of glutamine synthetase and delta-9 desaturase in families of Pacific oyster Crassostrea gigas and susceptibility to summer mortality. J. Exp. Mar. Biol. Ecol. 2007, 349, 272–283. [Google Scholar] [CrossRef]
- Lee, J.; Kim, N.-H.; Choi, Y.; Yang, E.; Yu, H.; Kwon, C.W.; Chang, P.-S. Divergent substrate specificities and regioselectivities of three lipase isoforms from Cordyceps militaris: Combinatorial advantages for entomopathogenicity and prospects as biocatalysts. Enzym. Microb. Technol. 2022, 161, 110117. [Google Scholar] [CrossRef]
- Udupa, P.; Kumar, A.; Parit, R.; Ghosh, D.K. Acyl-CoA binding protein regulates nutrient-dependent autophagy. Metabolism 2022, 36, 155338. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Song, W.; Yuan, H.; Hu, N.; Tan, B.; Zhang, S. Comparative transcriptome analysis revealed that dietary zymosan-A improved the immunity of Penaeus vannamei by regulating the TLR signaling pathway. Aquaculture 2022, 561, 738603. [Google Scholar] [CrossRef]
- Yin, X.; Zhuang, X.; Liao, M.; Huang, L.; Cui, Q.; Liu, C.; Dong, W.; Wang, F.; Liu, Y.; Wang, W. Transcriptome analysis of Pacific white shrimp (Litopenaeus vannamei) hepatopancreas challenged by Vibrio alginolyticus reveals lipid metabolic disturbance. Fish Shellfish Immunol. 2022, 123, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, J.I.; Tovar, M.O.; Blanco, A.M.; Gómez-Requeni, P.; Unniappan, S.; Canosa, L.F. Influence of water salinity on genes implicated in somatic growth, lipid metabolism and food intake in Pejerrey (Odontesthes bonariensis). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 210, 29–38. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, C.; Zhang, X.; Li, X.; Yao, X.; He, G.; Cao, J.; Chi, S.; Zhang, S.; Tan, B.; et al. Evaluation of ratios of fish-to-soybean oils on growth, lipid and cholesterol metabolism and muscle metabolites of Pacific white shrimp (Litopenaeus vannamei) fed low fishmeal diets containing Clostridium autoethanogenum protein. Aquac. Rep. 2022, 27, 101417. [Google Scholar] [CrossRef]
- Bianchini, A.; Lauer, M.M.; Nery, L.E.M.; Colares, E.P.; Monserrat, J.M.; Filho, E.A.d.S. Biochemical and physiological adaptations in the estuarine crab Neohelice granulata during salinity acclimation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 423–436. [Google Scholar] [CrossRef]
- Ge, Q.; Wang, J.; Li, J.; Li, J. Effect of high alkalinity on shrimp gills: Histopathological alternations and cell specific responses. Ecotoxicol. Environ. Saf. 2023, 256, 114902. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wu, X.; Ling, F.; Wang, Z.; Ma, K.; Xie, N.; Li, J.; Feng, J.; Li, J. Transcriptome, Proteome, Histology, and Biochemistry Analysis of Oriental River Prawn Macrobrachium nipponense under Long-term Salinity Exposure. Front. Mar. Sci. 2023, 9, 1104984. [Google Scholar] [CrossRef]
- Zhang, R.; Shi, X.; Liu, Z.; Sun, J.; Sun, T.; Lei, M. Histological, Physiological and Transcriptomic Analysis Reveal the Acute Alkalinity Stress of the Gill and Hepatopancreas of Litopenaeus vannamei. Mar. Biotechnol. 2023, 25, 588–602. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | Sequence (5′-3′) | Up or Down |
|---|---|---|---|
| CL12563Contig1 | Cytochrome p450 | F: AATGGTTGCGAGTCGAAGGT | Up |
| R: ACATCTGGGTACTTGGCTGC | |||
| CL1288Contig1 | Na+/K+ symporter | F: CGCCTTCGATTTGCATCTGG | Up |
| R: TGCACTCGATTCCTGTTGCT | |||
| CL32673Contig1 | Sodium- and chloride-dependent glycine transporter | F: ATGTACTACAGTGTCGGGCC | Up |
| R: GAGGAGCCCATTCTGGAAGA | |||
| CL26688Contig1 | sodium-dependent proline transporter-like | F: CATGCCGCTCATGTTCTTCG | Up |
| R: GGTCACTGATCGCACCAGAA | |||
| CL22475Contig1 | Sulfatase | F: TCGAACTGGTTACACAGGGC | Up |
| R: CTCCTAGTGGTGGTAGGCCT | |||
| CL10549Contig1 | Monocarboxylate transporter | F: TGGGTCATCGTTTTCGCTGA | Up |
| R: AGCCGTGAAGCACAATACCA | |||
| CL23523Contig1 | Fucosyltransferase | F: GCGACATGAAAACACTGCCA | Up |
| R: AAAACCAAATGTGCCGACGG | |||
| CL2748Contig1 | Mannosyl | F: CGCCTACATCACGCAGAGAT | Up |
| R: TTCGCCACCTGACTCTTTCC | |||
| CL27361Contig1 | C-type lectin | F: ACATGGACCAATGGATCGGG | Up |
| R: AGCTGATCCATCGATGTCTCG | |||
| CL15589Contig1 | Crustacyanin-like lipocalin | F: GTACTAGCAGCCGTCTTGGG | Down |
| R: TCGATGCCCATTTCGGTGAA | |||
| CL1729Contig1 | Apolipophorin | F: GCCAAAGCAAAGTGGGAAGG | Down |
| R: GTGGAAGTCGACCTTGCTCA | |||
| CL12234Contig1 | Monoglyceride lipase | F: GGATATCCCGGGCTTCCTTG | Down |
| R: AGCCTCATGACGAACACTGG | |||
| CL318Contig1 | Delta-9 desaturase | F: GTGTGACAGTCCTGAGGTCG | Down |
| R: AACGCACGTCGAGGATTCTT | |||
| CL26968Contig1 | Lipase 3-like isoform | F: CGAGTCAGCCTAAGAGAGCG | Down |
| R: GCCTTGTAAGGGTCCCACAA | |||
| CL31728Contig1 | Acyl-CoA-binding protein | F: ATGAGGCTGCTGAGAAGGTC | Down |
| R: AAGCCTCCATAGCAGCATCC |
| Primer Name | Primer Sequence (5′–3′) | NCBI Database-Gene Accession Number |
|---|---|---|
| G-6-Pase F | F:CGTGGACCTTTCTTCATTAG | MK307768.1 |
| G-6-Pase R | R:ACCATCAACCATTTGAGAAG | |
| HK F | F:TGTTCCCCAGCCGATTATGG | KT932419.1 |
| HK R | R:CGGCGCACTTGAATCCTTTG | |
| PK F | F:AGAAACCCAGACCAACCC | KP690140.1 |
| PK R | R:TAGTCGCCCTTGGCAGTC | |
| LDH F | F:TCGACATCTTCAAGGGCATC | MF033360.1 |
| LDH R | R:AGGTCAAAACGTCAACGGG | |
| β-actin F | F:CAGGTCGTGACTTGACCGAT | KY780290 |
| β-actin R | R:CGTCAGGGAGCTCGTAAGAC |
| Parameters | Different Salinity | |||
|---|---|---|---|---|
| 0‰ | 8‰ | 14‰ | 22‰ | |
| SR (%) | 79.03 ± 3.82 ab | 83.60 ± 2.61 ab | 87.17 ± 4.30 a | 69.91 ± 1.82 b |
| WGR (%) | 26.69 ± 3.95 a | 31.14 ± 2.59 a | 40.39 ± 5.41 b | 15.82 ± 4.46 c |
| HSI (%) | 3.92 ± 0.14 a | 3.80 ± 0.60 a | 4.76 ± 0.45 b | 4.00 ± 0.20 a |
| Sample | Raw_Reads | Raw_Bases | Clean_Reads | Clean_Bases | Valid_Bases | Q30 | GC |
|---|---|---|---|---|---|---|---|
| S0-1 | 49,299,632 | 7,394,944,800 | 48,187,746 | 7,047,263,043 | 95.30% | 95.50% | 46.06% |
| S0-2 | 49,394,440 | 7,409,166,000 | 48,186,892 | 7,052,820,332 | 95.19% | 95.27% | 47.26% |
| S0-3 | 49,316,632 | 7,397,494,800 | 48,230,800 | 7,052,349,647 | 95.33% | 95.46% | 47.44% |
| S14-1 | 48,933,058 | 7,339,958,700 | 47,935,956 | 7,009,780,552 | 95.50% | 95.68% | 46.58% |
| S14-2 | 48,934,960 | 7,340,244,000 | 47,982,628 | 7,007,194,602 | 95.46% | 95.83% | 46.89% |
| S14-3 | 48,973,892 | 7,346,083,800 | 47,969,240 | 7,018,985,183 | 95.55% | 95.70% | 47.55% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Ye, Y.; Li, W.; Liu, X.; Zhao, Y.; Jiang, Q.; Che, X. Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense. Animals 2023, 13, 2884. https://doi.org/10.3390/ani13182884
Li Y, Ye Y, Li W, Liu X, Zhao Y, Jiang Q, Che X. Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense. Animals. 2023; 13(18):2884. https://doi.org/10.3390/ani13182884
Chicago/Turabian StyleLi, Yiming, Yucong Ye, Wen Li, Xingguo Liu, Yunlong Zhao, Qichen Jiang, and Xuan Che. 2023. "Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense" Animals 13, no. 18: 2884. https://doi.org/10.3390/ani13182884
APA StyleLi, Y., Ye, Y., Li, W., Liu, X., Zhao, Y., Jiang, Q., & Che, X. (2023). Effects of Salinity Stress on Histological Changes, Glucose Metabolism Index and Transcriptomic Profile in Freshwater Shrimp, Macrobrachium nipponense. Animals, 13(18), 2884. https://doi.org/10.3390/ani13182884