
Association between Polymorphism in the Janus Kinase 2 (JAK2) Gene and Selected Performance Traits in Cattle and Sheep
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. DNA Isolation
2.3. Sequencing
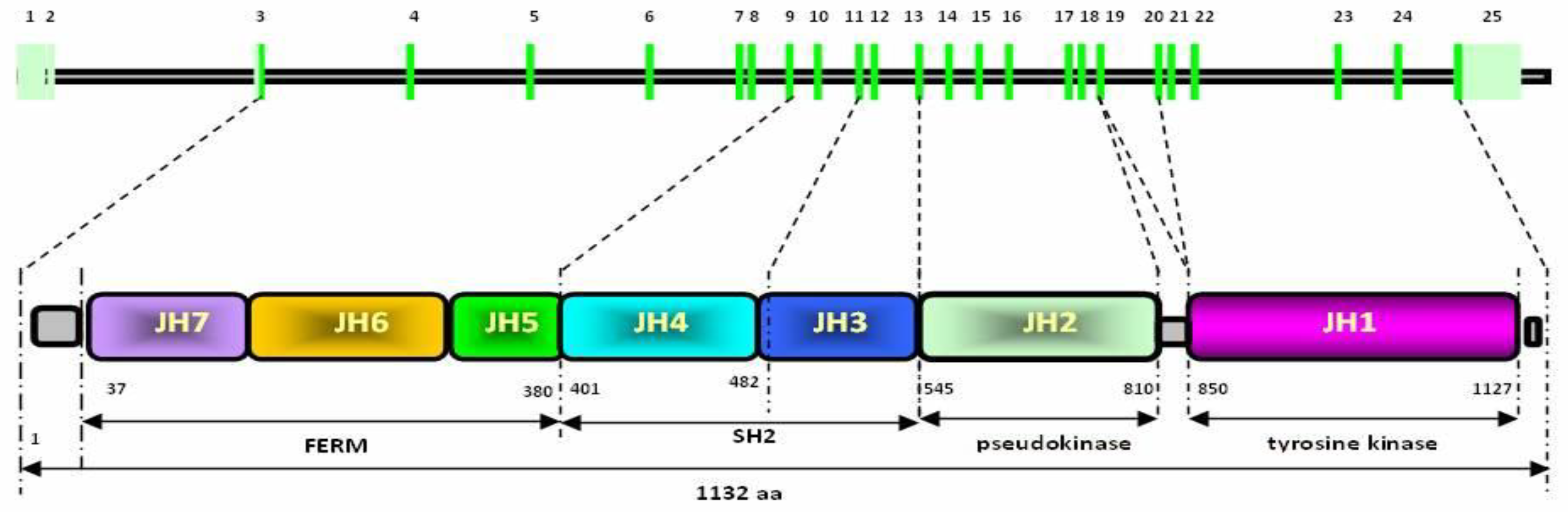
2.4. Bovine JAK2 Gene Polymorphism
2.5. Ovine JAK2 Gene Polymorphism
2.6. PCR Conditions
2.7. Digestion
2.8. Statistical Analysis
- (Growth traits—cattle: BWT, ADG, and WWT210; sheep: BW2D, BW30D, BW56D, BWM, ADG2–56, and ADG30–56)
- (1)
- Yijklm = μ + Gi +sj + BYSk + [LSl] + eijklmwhere Yijklm is the analyzed trait; μ the overall mean; Gi the fixed effect of JAK2 genotype (i = 1, …3); sj the random effect of sire (Hereford j = 1, …31; Angus j = 1, …39; Limousin j = 1, …24; Pomeranian j = 1, …3; Suffolk j = 1, …5); BYSk the fixed effect of birth year/season (Hereford k = 1, …20; Angus k = 1, …20; Limousin k = 1, …20; Pomeranian k = 1, …16; Suffolk k = 1, …16); [LSl] the fixed effect of litter size of mother (only in sheep: 1 single, 2 twins), and eijklm the random error.
- (Cattle: age and body weight at first calving as well as calving interval)
- (2)
- Yijkl = μ + Gi + sj +CYSk + eijklwhere Yijkl is the analyzed trait; μ the overall mean; Gi the fixed effect of JAK2 genotype (i = 1, …3); sj the random effect of sire (Hereford j = 1, …30; Angus j = 1, …39; Limousin j = 1, …21); CYSk the fixed effect of year/season of 1st calving (Hereford k = 1, …22; Angus k = 1, …27; Limousin k = 1, …21); and eijkl the random error.
- (Only sheep: fertility, prolificacy, and lamb survival)
- (3)
- Yijkl = μ + Gi + LSj + LYk + eijklwhere Yijkl is the analyzed trait; μ the overall mean; Gi the fixed effect of JAK2 genotype (i = 1, …3); LSj the fixed effect of litter size (1 single, 2 twins); LYk the fixed effect of lambing year (Pomeranian k = 1, …6; Suffolk k = 1, …8); and eijkl the random error.
3. Results
3.1. Sequencing
3.2. Bovine JAK2 Frequencies
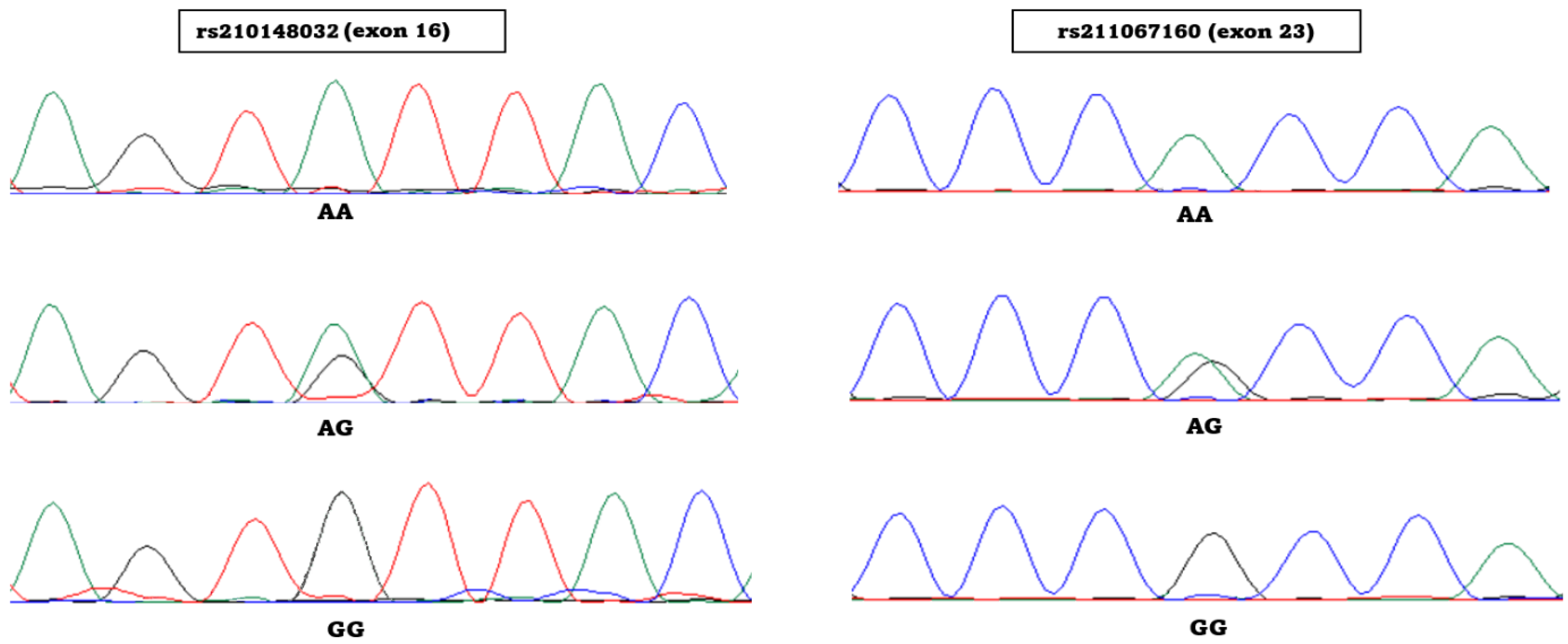
3.3. Bovine JAK2/e16/RsaI Associations
3.4. Bovine JAK2/e23/HaeIII Associations
3.5. Ovine JAK2 Frequencies
3.6. Ovine JAK2 Associations—Growth Traits
3.7. Ovine JAK2 Associations-Reproductive Traits
4. Discussion
4.1. GWAS Studies
4.2. Physiological Implications
4.3. Genetic Implications
4.4. Involvement of JAK2 in the Formation of Reproductive Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de Los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Tong, H.; Küken, A.; Nikoloski, Z. Integrating molecular markers into metabolic models improves genomic selection for Arabidopsis growth. Nat. Commun. 2020, 11, 2410. [Google Scholar] [CrossRef]
- Katoh, K.; Takahashi, T.; Kobauashi, Y.; Obara, Y. Somatropic Axis and Nutrition in Young Ruminants around Weaning Time. Asian-Australas. J. Anim. Sci. 2007, 20, 1156–1168. [Google Scholar] [CrossRef]
- Chmielewska, P.; Barwiak, K.; Dańko, P.; Ciesielka, M.; Beń-Skowronek, I. Insulin-like growth factors and theirs role in pathogenesis od diabetic nephropathy and retinopathy. Pediatr. Endocrinol. 2018, 17, 191–200. [Google Scholar]
- Rosenfeld, R.G.; Hwa, V. Biology of the somatotroph axis (after the pituitary). Ann. Endocrinol. 2017, 78, 80–82. [Google Scholar] [CrossRef]
- Backeljauw, P.F.; Dattani, M.T.; Cohen, P.; Rosenfeld, R.G. Disorders of Growth Hormone/Insulin-Like Growth Factor Secretion and Action, 4th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 392–404. [Google Scholar]
- He, K.; Loesch, K.; Cowan, J.W.; Li, X.; Deng, L.; Wang, X.; Jiang, J.; Frank, S.J. Janus kinase 2 enhances the stability of the mature growth hormone receptor. Endocrinology 2005, 146, 4755–4765. [Google Scholar] [CrossRef]
- Babon, J.J.; Lucet, I.S.; Murphy, J.M.; Nicola, N.A.; Varghese, L.N. The molecular regulation of Janus kinase (JAK) activation. Biochem. J. 2014, 462, 1–13. [Google Scholar] [CrossRef]
- Lacronique, V.; Boureux, A.; Valle, V.D.; Poirel, H.; Quang, C.T.; Mauchauffé, M.; Berthou, C.; Lessard, M.; Berger, R.; Ghysdael, J.; et al. A TEL-JAK2 fusion protein with constitutive kinase activity in human leukemia. Science 1997, 278, 1309–1312. [Google Scholar] [CrossRef]
- Rane, S.G.; Reddy, E.P. Janus kinase: Components of multiple signaling pathways. Oncogene 2000, 19, 5662–5679. [Google Scholar] [CrossRef]
- Khan, M.Z.; Khan, A.; Xiao, J.; Ma, Y.; Ma, J.; Gao, J.; Cao, Z. Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production. Animals 2020, 10, 2107. [Google Scholar] [CrossRef]
- Brenaut, P.; Bangera, R.; Bevilacqua, C.; Rebours, E.; Cebo, C.; Martin, P. Validation of RNA isolated from milk fat globules to profile mammary epithelial cell expression during lactation and transcriptional response to a bacterial infection. J. Dairy Sci. 2012, 95, 6130–6144. [Google Scholar] [CrossRef]
- Yamaji, D.; Kang, K.; Robinson, G.W.; Hennighausen, L. Sequential activation of genetic programs in mouse mammary epithelium during pregnancy depends on STAT5A/Bconcentration. Nucleic Acids Res. 2013, 41, 1622–1636. [Google Scholar] [CrossRef]
- Groner, B.; Gouilleux, F. Prolactin-mediated gene activation in mammary epithelial cells. Curr. Opin. Genet. Dev. 1995, 5, 587–594. [Google Scholar] [CrossRef]
- Bole-Feysot, C. Prolactin (PRL) and Its Receptor: Actions, Signal Transduction Pathways and Phenotypes Observed in PRL Receptor Knockout Mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Szewczuk, M. Association of a genetic marker at the bovine Janus kinase 2 locus (JAK2/RsaI) with milk production traits of four cattle breeds. J. Dairy Res. 2015, 82, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Padzik, N.; Szewczuk, M. Association between polymorphism in the JAK2 gene (JAK2/e20/RsaI) and selected performance parameters in beef cattle. Anim. Sci. Pap. Rep. 2020, 38, 333–344. [Google Scholar]
- Padzik, N.; Szewczuk, M.; Ropka-Molik, K. Distribution of JAK2 genotypes across Suffolk and Pomeranian sheep. Small Rumin. Res. 2021, 194, 106282. [Google Scholar] [CrossRef]
- Strzetelski, J.A.; Brzóska, F.; Kowalski, Z.M.; Osięgłowski, S. Nutritional Recommendations for Ruminants and Feed Value Tables; Praca zbiorowa Instytut Zootechniki PIB: Kraków, Poland, 2014. [Google Scholar]
- Polido, P.B.; Ferreira, G.F.; Alberton, O.; De Souza, S.G.H. Marcadores moleculares aplicados nomelhoramento genético de bovinos. Arq. Ciênc. Vet. Zool. 2012, 15, 161–169. [Google Scholar]
- Grossi, D.A.; Venturini, G.C.; Paz, C.C.; Bezerra, L.A.; Lôbo, R.B.; Oliveira, J.A.; Munari, D.P. Genetic associations between age at first calving and heifer body weight and scrotal circumference in Nelore cattle. J. Anim. Breed. Genet. 2009, 126, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Menezes, I.P.P.; Gaiotto, F.; Suassuna, N.; Hoffmann, L.; Barroso, P. Susceptibility of Gossypium mustelinum Populations to the Main Cotton Diseases in Brazil. J. Agric. Sci. 2014, 6, 39–47. [Google Scholar] [CrossRef]
- de Oliveira Silva, R.M.; Stafuzza, N.B.; de Oliveira Fragomeni, B.; de Camargo, G.M.F.; Ceacero, T.M.; Cyrillo, J.N.D.S.G.; Baldi, F.; Boligon, A.A.; Mercadante, M.E.Z.; Lourenco, D.L.; et al. Genome-Wide Association Study for Carcass Traits in an Experimental Nelore Cattle Population. PLoS ONE 2017, 12, e0169860. [Google Scholar] [CrossRef] [PubMed]
- Keogh, K.; Carthy, T.R.; McClure, M.C.; Waters, S.M.; Kenny, D.A. Genome-wide association study of economically important traits in Charolais and Limousin beef cows. Animal 2021, 15, 100011. [Google Scholar] [CrossRef] [PubMed]
- Al-Mamun, H.A.; Kwan, P.; Clark, S.A.; Ferdosi, M.H.; Tellam, R.; Gondro, C. Genome-wide association study of body weight in Australian Merino sheep reveals an orthologous region on OAR6 to human and bovine genomic regions affecting height and weight. Genet. Sel. Evol. 2015, 47, 66. [Google Scholar] [CrossRef] [PubMed]
- Abiola, O.; Angel, J.M.; Avner, P.; Bachmanov, A.A.; Belknap, J.K.; Bennett, B.; Blankenhorn, E.P.; Blizard, D.A.; Bolivar, V.; Brockmann, G.A.; et al. Complex Trait Consortium. The nature and identification of quantitative trait loci: A community’s view. Nat. Rev. Genet. 2003, 4, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Yue, Y.; Yuan, C.; Liu, J.; Chen, Z.; Niu, C.; Sun, X.; Zhu, S.; Zhao, H.; Guo, T.; et al. Genome-Wide Association Study of Body Weight Traits in Chinese Fine-Wool Sheep. Animals 2020, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.G.; Webb, R. Ovarian follicular dominance: The role of intraovarian growth factors and novel proteins. Rev. Reprod. 1997, 2, 139–146. [Google Scholar] [CrossRef]
- Rudnicki, M.A.; Williams, B.O. Wnt signaling in bone and muscle. Bone 2015, 80, 60–66. [Google Scholar] [CrossRef]
- Trenerry, M.K.; Della Gatta, P.A.; Cameron-Smith, D. JAK/STAT signaling and human in vitro myogenesis. BMC Physiol. 2011, 9, 6. [Google Scholar] [CrossRef][Green Version]
- Liu, J.; Jing, X.; Gan, L.; Sun, C. The JAK2/STAT3 signal pathway regulates the expression of genes related to skeletal muscle development and energy metabolism in mice and mouse skeletal muscle cells. Biosci. Biotechnol. Biochem. 2012, 76, 1866–1870. [Google Scholar] [CrossRef]
- Huang, Z.; Zhong, L.; Zhu, J.; Xu, H.; Ma, W.; Zhang, L.; Shen, Y.; Law, B.Y.; Ding, F.; Gu, X.; et al. Inhibition of IL-6/JAK/STAT3 pathway rescues denervation-induced skeletal muscle atrophy. Ann. Transl. Med. 2020, 8, 1681. [Google Scholar] [CrossRef]
- Consitt, L.A.; Wideman, L.; Hickey, M.S.; Morrison, R.F. Phosphorylation of the JAK2-STAT5 pathway in response to acute aerobic exercise. Med. Sci. Sports Exerc. 2008, 40, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, X.; Xu, Y.; Feng, Y.; Sheng, W.; Cen, J.; Wu, D.; Han, Y. Impact of JAK2V617F Mutation Burden on Disease Phenotype in Chinese Patients with JAK2V617F-positive Polycythemia Vera (PV) and Essential thrombocythemia (ET). Int. J. Med. Sci. 2016, 13, 85–91. [Google Scholar] [CrossRef]
- Beurlet, S.; Krief, P.; Sansonetti, A.; Briend-Marchal, A.; Kiladjian, J.J.; Padua, R.A.; Chomienne, C.; Cassinat, B. Identification of JAK2 mutations in canine primary polycythemia. Exp. Hematol. 2011, 39, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Yampolsky, L.Y.; Stoltzfus, A. The exchangeability of amino acids in proteins. Genetics 2005, 170, 1459–1472. [Google Scholar] [CrossRef] [PubMed]
- Creixell, P.; Schoof, E.M.; Tan, C.S.; Linding, R. Mutational properties of amino acid residues: Implications for evolvability of phosphorylatable residues. Philos. Trans. R. Soc. B Biol. Sci. 2012, 19, 2584–2593. [Google Scholar] [CrossRef] [PubMed]
- Robert, F.; Pelletier, J. Exploring the Impact of Single-Nucleotide Polymorphisms on Translation. Front Genet. 2018, 9, 507. [Google Scholar] [CrossRef]
- Brule, C.E.; Grayhack, E.J. Synonymous Codons: Choose Wisely for Expression. Trends Genet. 2017, 33, 283–297. [Google Scholar] [CrossRef]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target Ther. 2021, 6, 402. [Google Scholar] [CrossRef]
- Funakoshi-Tago, M.; Pelletier, S.; Moritake, H.; Parganas, E.; Ihle, J.N. Jak2 FERM domain interaction with the erythropoietin receptor regulates Jak2 kinase activity. Mol. Cell Biol. 2008, 28, 5792–5801. [Google Scholar] [CrossRef]
- Boyer, C.N.; Griffith, A.P.; DeLong, K.L. Reproductive Failure and Long-Term Profitability of Spring- and Fall-Calving Beef Cows. J. Agric. Resour. Econ. 2020, 45, 78–91. [Google Scholar] [CrossRef]
- Moorey, S.E.; Biase, F.H. Beef heifer fertility: Importance of management practices and technological advancements. J. Anim. Sci. 2020, 11, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Lee, J.; Do, C. Genetic relationship of age at first calving with conformation traits and calving interval in Hanwoo cows. J. Anim. Sci. Technol. 2021, 63, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Brzáková, M.; Čítek, J.; Svitáková, A.; Veselá, Z.; Vostrý, L. Genetic Parameters for Age at First Calving and First Calving Interval of Beef Cattle. Animals 2020, 10, 2122. [Google Scholar] [CrossRef]
- Krupová, Z.; Krupa, E.; Wolfová, M. Economic weights of current and new breeding objective traits in Aberdeen Angus. J. Anim. Sci. 2020, 65, 77–85. [Google Scholar] [CrossRef]
- Ghiasi, H.; Abdollahi-Arpanahi, R. The candidate genes and pathways affecting litter size in sheep. Small Rumin. Res. 2021, 205, 106546. [Google Scholar] [CrossRef]
- Luridiana, S.; Cosso, G.; Pulinas, L.; Di Stefano, M.V.; Curone, G.; Carcangiu, V.; Mura, M.C. New polymorphisms at MTNR1A gene and their association with reproductive resumption in sarda breed sheep. Theriogenology 2020, 158, 438–444. [Google Scholar] [CrossRef]
- Sánchez-Ramos, R.; Trujano-Chavez, M.Z.; Gallegos-Sánchez, J.; Becerril-Pérez, C.M.; Cadena-Villegas, S.; Cortez-Romero, C. Detection of Candidate Genes Associated with Fecundity through Genome-Wide Selection Signatures of Katahdin Ewes. Animals 2023, 13, 272. [Google Scholar] [CrossRef]
- Yang, Q.; Yan, H.; Li, J.; Xu, H.; Wang, K.; Zhu, H.; Lan, X. A novel 14-bp duplicated deletion within goat GHR gene is significantly associated with growth traits and litter size. Anim. Genet. 2017, 48, 499–500. [Google Scholar] [CrossRef]
- Sebastiano, L.; Consuelo, M.M.; Veronica, D.S.M.; Luisa, P.; Giovanni, C.; Michella, N.; Vincenzo, C. Polymorphism of insulin-like growth factor 1 gene and its relationship with reproductive performances and milk yield in Sarda dairy sheep. Vet. Anim. Sci. 2019, 15, 100084. [Google Scholar] [CrossRef]
- Abate, Z. Review of the reproductive performances of indigenous sheep in Ethiopia. Glob. J. Biol. Agric. 2016, 6, 117–126. [Google Scholar]
- Núñez-Torres, O.P.; Almeida-Secaira, R.I. Quantitative genetics: Principles of farming in livestock production Genética cuantitativa: Principios de la crianza en la producción pecuaria. J. Selva Andin. Anim. 2022, 9, 23–36. [Google Scholar]
- Metzler-Zebeli, B.U.; Hollmann, M.; Sabitzer, S.; Podstatzky-Lichtenstein, L.; Klein, D.; Zebeli, Q. Epithelial response to high-grain diets involves alteration in nutrient transporters and Na+/K+-ATPase Mrna expression in rumen and colon of goats. J. Anim. Sci. 2013, 91, 4256–4266. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Chu, M.; Li, Q.; Jin, M.; Fei, X.; Ma, L.; Zhang, L.; Wei, C. Transcriptomic Analysis Provides Novel Insights into Heat Stress Responses in Sheep. Animals 2019, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Gootwine, E. Invited review: Opportunities for genetic improvement toward higher prolificacy in sheep. Small Rumin. Res. 2020, 186, 106090. [Google Scholar] [CrossRef]
- Ivanova, T.; Stoikova-Grigorova, R.; Bozhilova-Sakova, M.; Ignatova, M.; Dimitrova, I.; Koutev, V. Phenotypic and genetic characteristics of fecundity in sheep. A review. Bulg. J. Agric. Sci. 2021, 27, 1002–1008. [Google Scholar]
- Lipecka, C.Z. Weaning and Care of Offspring. In Breeding, Rearing and Use of Sheep, 3rd ed.; Niżnikowski, R., Ed.; Wieś Jutra: Warszawa, Poland, 2011; pp. 89–103. [Google Scholar]
- Clune, T.; Lockwood, A.; Hancock, S.; Thompson, A.N.; Beetson, S.; Campbell, A.J.D.; Glanville, E.; Brookes, D.; Trengove, C.; O’Handley, R.; et al. Abortion and Lamb Mortality between Pregnancy Scanning and Lamb Marking for Maiden Ewes in Southern Australia. Animals 2022, 12, 10. [Google Scholar] [CrossRef]
- Hutchison, D.; Clarke, B.E.; Hancock, S.; Thompson, A.N.; Bowen, E.; Jacobson, C. Lower Reproductive Rate and Lamb Survival Contribute to Lower Lamb Marking Rate in Maiden Ewes Compared to Multiparous Ewes. Animals 2022, 12, 513. [Google Scholar] [CrossRef]
- Fesseha, H.; Gebremichael, G.; Asefa, I.; Edaso, T. Study on incidence of lamb morbidity and mortality and associated risk factors in the mixed crop-livestock production system of Gewata District, Kaffa zone, southwestern Ethiopia. Anim. Dis. 2023, 3, 11. [Google Scholar] [CrossRef]
- Martins, L.; Oliveira, L.C.C. Colostrum as the speed up key for ruminant newborn: What do we know and should further characterize. J. Dairy Vet. Anim. Res. 2020, 9, 95–98. [Google Scholar]
- PZO. Polish Union of Sheep-Farmers. 2022. Available online: http://Pzow.Pl/ (accessed on 27 May 2023).
- Paraponiak, P.; Wieczorek-Dąbrowska, M. Impact of organic farming on improving reproductive parameters in sheep. Wiadomości Zootech. 2019, 4, 30–40. [Google Scholar]
- Lourencon, R.V.; Patra, A.K.; Puchala, R.; Dawson, L.J.; Ribeiro, L.P.D.S.; Encinas, F.; Goetsch, A.L. Effects of Nutritional Plane at Breeding on Feed Intake, Body Weight, Condition Score, Mass Indexes, and Chemical Composition, and Reproductive Performance of Hair Sheep. Animals 2023, 13, 735. [Google Scholar] [CrossRef] [PubMed]
- Łozicki, A. Rules for Feeding Sheep. In Breeding, Rearing and Use of Sheep, 3rd ed.; Niżnikowski, R., Ed.; Wieś Jutra: Warszawa, Poland, 2011; pp. 120–129. [Google Scholar]
- Skliarov, P.; Perez, C.; Petruscha, V.; Fedorenko, S.; Bilyi, D. Induction and synchronization of oestrus in sheep and goats. J. Cent. Eur. Agric. 2021, 22, 39–53. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Sequence (5′ → 3′) | Polymorphism Position ** and Type | Codon and Amino Acid(s) *** | PCR-ACRS (Restriction Enzyme, Cleavage Site, and Genotypes) |
|---|---|---|---|
| JAK2e16mF gggccctggacatactaagtg JAK2e16mR gtcttttggcaaaactgtaG * | rs210148032 g.39400906A>G Missense | ATT → GTT p.Ile704Val | RsaI (gt^ac) PCR amplicon 211 bp AA 190 bp + 21 bp AG 211 bp + 190 bp + 21 bp GG 211 bp (no cut) |
| JAK2e23mF catatattgacaagagtaaaagccG * JAK2e23mR tccccacctttcaaaacttc | rs211067160 g.39383281A>G Synonymous | CCA → CCG p.Pro1057 | HaeIII (gg^cc) PCR amplicon 211 bp AA 211 bp (no cut) AG 211 bp + 185 bp + 26 bp GG 185 bp + 26 bp |
| Primer Sequence (5′ → 3′) | Polymorphism Position * and Type | Codon and Amino Acid ** | PCR-RFLP (Restriction Enzyme, Cleavage Site, and Genotypes) |
|---|---|---|---|
| oJAK2e6F ttgaccttgttaaatgtatatgttctg | rs160146162 g.73397955A>G Synonymous | GAA → GAG p.Glu177 | EarI (CTCTTC(1/4)^) PCR amplicon 280 bp AA 280 bp (no cut) AG 280 bp + 170 bp + 110 bp GG 170 bp + 110 bp |
| oJAK2e6R ttgcataagaaaattacctgatagagc | rs160146160 g.73398012G>A Synonymous | ACG → ACA p.Thr196 | PCR amplicon 280 bp not applicable (sequencing) |
| oJAK2e24F tctgcttgaaattaaatgtaccaaa oJAK2e24R tcagtgaactgcataaactgacc | rs160146116 g.73442093A>G Synonymous | CTA → CTG p.Leu1082 | Hpy188III (TC^NNGA) PCR amplicon 261 bp AA 205 bp + 56 bp AG 205 bp + 137 bp + 68 bp + 56 bp GG 137 bp + 68 bp + 56 bp |
| Breed | JAK2/e16/RsaI | Total | allele | ||||
| AA | AG | GG | A | G | |||
| Hereford | n | 161 | 95 | 20 | 276 | 0.7554 | 0.2446 |
| Frequency | 0.5833 | 0.3442 | 0.0725 | 1.0000 | |||
| Angus | n | 195 | 110 | 28 | 333 | 0.7508 | 0.2492 |
| Frequency | 0.5856 | 0.3303 | 0.0841 | 1.0000 | |||
| Limousin | n | 95 | 53 | 12 | 160 | 0.7594 | 0.2406 |
| Frequency | 0.5937 | 0.3313 | 0.0750 | 1.0000 | |||
| Breed | JAK2/e23/HaeIII | Total | allele | ||||
| AA | AG | GG | A | G | |||
| Hereford | n | 165 | 83 | 23 | 271 | 0.7620 | 0.2380 |
| Frequency | 0.6088 | 0.3063 | 0.0849 | 1.0000 | |||
| Angus | n | 61 | 132 | 152 | 345 | 0.3681 | 0.6319 |
| Frequency | 0.1768 | 0.3826 | 0.4406 | 1.0000 | |||
| Limousin | n | 28 | 72 | 43 | 143 | 0.4476 | 0.5524 |
| Frequency | 0.1958 | 0.5035 | 0.3007 | 1.0000 | |||
| Breed | Genotype | Birth Weight (kg) | Average Daily Gain (g) | Weaning Weight (kg) | Age at First Calving (Days) | Body Weight at First Calving (kg) | Calving Interval (Days) |
|---|---|---|---|---|---|---|---|
| Hereford | AA | 34.7 a (0.36) | 1091 AB (13.11) | 262.2 a (2.85) | 1039 A (28.20) | 589.4 Aa (2.11) | 499 ab (16.06) |
| AG | 34.0 (0.33) | 959 A (14.84) | 254.3 (3.50) | 975 a (31.00) | 571.5 a (5.90) | 420 a (13.57) | |
| GG | 32.9 a (0.51) | 926 B (16.60) | 251.1 a (6.97) | 813 Aa (37.69) | 560.6 A (6.79) | 399 b (16.36) | |
| Angus | AA | 36.2 a (0.30) | 994 (7.76) | 247.1 (1.87) | 1076 (19.20) | 564.6 (3.33) | 416 Aa (2.02) |
| AG | 37.3 (0.39) | 1011 (8.13) | 251.6 (2.50) | 1057 (26.91) | 562.3 (3.81) | 403 ab (2.23) | |
| GG | 38.4 a (0.59) | 1012 (8.58) | 255.5 (7.89) | 1011 (25.83) | 574.5 (8.46) | 389 Ab (1.22) | |
| Limousin | AA | 35.2 a (0.41) | 1001 (23.62) | 275.5 A (3.22) | 1077 (14.55) | 610.8 a (10.27) | 469 (21.51) |
| AG | 34.2 (0.57) | 1001 (35.12) | 261.3 (3.61) | 1045 (14.83) | 610.3 b (13.77) | 479 (27.96) | |
| GG | 32.6 a (0.65) | 954 (10.35) | 248.3 A (6.19) | 1090 (15.67) | 581.2 ab (5.69) | 466 (6.69) |
| Breed | Genotype | Birth Weight (kg) | Average Daily Gain (g) | Weaning Weight (kg) | Age at First Calving (Days) | Body Weight at First Calving (kg) | Calving Interval (Days) |
|---|---|---|---|---|---|---|---|
| Hereford | AA | 34.5 (0.34) | 1060 A (12.86) | 260.8 a (2.53) | 1020 a (27.83) | 584.9 (3.26) | 487 a (15.87) |
| AG | 34.1 (0.38) | 1018 B (24.03) | 257.9 (4.95) | 988 (37.85) | 575.6 (4.05) | 430 (14.72) | |
| GG | 33.9 (1.03) | 906 AB (16.51) | 244.4 a (3.47) | 874 a (51.07) | 571.9 (7.08) | 397 a (16.73) | |
| Angus | AA | 37.16 (0.71) | 1005 (14.27) | 249.6 (3.66) | 1065 (25.94) | 559.4 (5.42) | 423 (4.37) |
| AG | 37.40 (0.42) | 1017 (11.25) | 250.8 (2.92) | 1021 (25.59) | 563.9 (4.72) | 400 (1.71) | |
| GG | 36.3 (0.29) | 993 (7.37) | 248.4 (1.99) | 1084 (20.98) | 566.0 (3.28) | 410 (2.08) | |
| Limousin | AA | 35.5 (0.77) | 1058 a (17.42) | 274.5 a (4.38) | 1075 (18.39) | 580.5 ab (3.13) | 413 a (28.82) |
| AG | 34.2 (0.40) | 990 (32.90) | 271.3 (3.91) | 1078 (15.23) | 618.8 a (12.95) | 504 a (27.69) | |
| GG | 35.2 (0.73) | 967 a (36.69) | 261.2 a (4.13) | 1056 (25.25) | 617.3 b (17.77) | 483 (32.79) |
| Breed | JAK2/e6/EarI | Total | allele | ||||
| AA | AG | GG | A | G | |||
| Pomeranian | n | 32 | 58 | 48 | 138 | 0.4420 | 0.5580 |
| Frequency | 0.2319 | 0.4203 | 0.3478 | 1.0000 | |||
| Suffolk | n | 29 | 43 | 123 | 195 | 0.2590 | 0.7410 |
| Frequency | 0.1487 | 0.2205 | 0.6308 | 1.0000 | |||
| Breed | JAK2/e6/seq | Total | allele | ||||
| AA | AG | GG | A | G | |||
| Pomeranian | n | 55 | 59 | 24 | 138 | 0.6123 | 0.3877 |
| Frequency | 0.3986 | 0.4275 | 0.1739 | 1.0000 | |||
| Suffolk | n | 49 | 110 | 36 | 195 | 0.5333 | 0.4667 |
| Frequency | 0.2513 | 0.5641 | 0.1846 | 1.0000 | |||
| Breed | JAK2/e24/Hpy188III | Total | allele | ||||
| AA | AG | GG | A | G | |||
| Pomeranian | n | 43 | 67 | 28 | 138 | 0.5544 | 0.4456 |
| Frequency | 0.3116 | 0.4855 | 0.2029 | 1.0000 | |||
| Suffolk | n | 49 | 128 | 18 | 195 | 0.5795 | 0.4205 |
| Frequency | 0.2513 | 0.6564 | 0.0923 | 1.0000 | |||
| Breed | Genotype | Body Weight at 2 Days (kg) | Body Weight at 30 Days (kg) | Body Weight at 56 Days (kg) | Average Daily Gains between 2–56 Days (g) | Average Daily Gains between 30–56 Days (g) | Body Weight at Mating (kg) | |
|---|---|---|---|---|---|---|---|---|
| JAK2/e6/EarI | Pomeranian | AA | 4.3 AB (0.04) | n/a | 19.3 AB (0.19) | 278 AB (3.52) | n/a | 54.4 (0.43) |
| AG | 4.0 BC (0.03) | n/a | 18.1 BC (0.15) | 262 Ba (2.78) | n/a | 53.8 (0.28) | ||
| GG | 3.6 AC (0.02) | n/a | 17.1 AC (0.18) | 251 Aa (3.20) | n/a | 53.2 (0.43) | ||
| Suffolk | AA | 4.8 AB (0.05) | 14.2 AB (0.13) | 23.5 AB (0.21) | 347 AB (3.44) | 359 AB (7.24) | 62.0 AB (0.55) | |
| AG | 4.1 B (0.02) | 13.0 B (0.19) | 20.7 Ba (0.25) | 307 Ba (4.39) | 297 Ba (9.86) | 58.9 B (0.39) | ||
| GG | 4.0 A (0.03) | 12.9 A (0.15) | 19.9 Aa (0.18) | 294 Aa (2.96) | 269 Aa (4.95) | 57.9 A (0.26) | ||
| JAK2/e6/seq | Pomeranian | AA | 3.8 A (0.04) | n/a | 17.9 A (0.18) | 261 A (3.16) | n/a | 53.8 (0.31) |
| AG | 3.9 B (0.04) | n/a | 17.7 B (0.16) | 256 B (2.73) | n/a | 54.2 (0.34) | ||
| GG | 4.3 AB (0.05) | n/a | 19.4 AB (0.24) | 280 AB (4.01) | n/a | 54.9 (0.53) | ||
| Suffolk | AA | 4.1 A (0.06) | 12.7 A (0.25) | 19.8 A (0.34) | 292 A (5.32) | 275 A (8.56) | 58.4 A (0.54) | |
| AG | 4.1 B (0.03) | 13.1 a (0.14) | 20.3 B (0.17) | 301 B (2.93) | 278 B (5.79) | 58.4 B (0.25) | ||
| GG | 4.6 AB (0.06) | 13.7 Aa (0.20) | 22.5 AB (0.32) | 333 AB (5.29) | 338 AB (8.67) | 60.2 AB (0.59) | ||
| JAK2/e24/Hpy188III | Pomeranian | AA | 4.2 AB (0.05) | n/a | 19.2 AB (0.20) | 279 AB (3.83) | n/a | 54.4 (0.43) |
| AG | 3.9 AC (0.04) | n/a | 17.8 AC (0.13) | 258 AC (2.14) | n/a | 53.8 (0.28) | ||
| GG | 3.6 BC (0.05) | n/a | 16.8 BC (0.15) | 244 BC (2.98) | n/a | 53.2 (0.43) | ||
| Suffolk | AA | 4.5 AB (0.06) | 14.1 AB (0.16) | 22.7 AB (0.26) | 338 AB (4.07) | 332 AB (9.04) | 60.1 Aa (0.54) | |
| AG | 4.1 AC (0.02) | 13.0 AC (0.12) | 20.2 AC (0.15) | 298 AC (2.46) | 278 AC (4.99) | 58.7 Ba (0.23) | ||
| GG | 3.7 BC (0.05) | 11.4 BC (0.40) | 17.7 BC (0.35) | 260 BC (5.98) | 243 BC (11.28) | 54.9 AB (0.46) |
| Breed | Genotype | Fertility (%) | Prolificacy (n/ewe) | Lamb Survival (%) | |
|---|---|---|---|---|---|
| JAK2/e6/EarI | Pomeranian | AA | 94.30 A (2.23) | 1.15 (0.05) | 93.55 A (2.21) |
| AG | 95.88 B (1.45) | 1.11 (0.04) | 94.28 B (1.67) | ||
| GG | 82.34 AB (2.70) | 1.18 (0.05) | 81.38 AB (3.86) | ||
| Suffolk | AA | 91.03 (2.89) | 1.19 A (0.06) | 95.40 A (1.92) | |
| AG | 94.41 (1.11) | 1.18 a (0.05) | 96.34 B (1.48) | ||
| GG | 94.40 (1.66) | 1.09 Aa (0.02) | 84.52 AB (2.00) | ||
| JAK2/e6/seq | Pomeranian | AA | 88.30 A (2.35) | 1.11 (0.04) | 86.20 A (2.88) |
| AG | 90.31 a (2.05) | 1.18 (0.04) | 89.69 (2.64) | ||
| GG | 97.83 Aa (1.59) | 1.13 (0.05) | 97.68 A (1.66) | ||
| Suffolk | AA | 94.62 a (1.77) | 1.13 (0.04) | 87.06 (2.84) | |
| AG | 94.74 b (1.11) | 1.14 (0.03) | 92.09 A (1.53) | ||
| GG | 90.34 ab (2.47) | 1.06 (0.03) | 80.82 A (4.23) | ||
| JAK2/e24/Hpy188III | Pomeranian | AA | 95.17 A (1.76) | 1.13 A (0.04) | 95.77 A (1.66) |
| AG | 93.48 B (1.79) | 1.08 B (0.03) | 90.66 (1.93) | ||
| GG | 81.33 AB (3.74) | 1.27 AB (0.08) | 84.00 A (4.46) | ||
| Suffolk | AA | 95.05 a (1.44) | 1.15 (0.03) | 92.64 A (2.43) | |
| AG | 94.17 (1.13) | 1.11 (0.03) | 89.44 B (1.66) | ||
| GG | 88.89 a (3.81) | 1.11 (0.05) | 73.15 AB (5.43) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oster, N.; Szewczuk, M.A.; Zych, S.; Stankiewicz, T.; Błaszczyk, B.; Wieczorek-Dąbrowska, M. Association between Polymorphism in the Janus Kinase 2 (JAK2) Gene and Selected Performance Traits in Cattle and Sheep. Animals 2023, 13, 2470. https://doi.org/10.3390/ani13152470
Oster N, Szewczuk MA, Zych S, Stankiewicz T, Błaszczyk B, Wieczorek-Dąbrowska M. Association between Polymorphism in the Janus Kinase 2 (JAK2) Gene and Selected Performance Traits in Cattle and Sheep. Animals. 2023; 13(15):2470. https://doi.org/10.3390/ani13152470
Chicago/Turabian StyleOster, Nicola, Małgorzata Anna Szewczuk, Sławomir Zych, Tomasz Stankiewicz, Barbara Błaszczyk, and Marta Wieczorek-Dąbrowska. 2023. "Association between Polymorphism in the Janus Kinase 2 (JAK2) Gene and Selected Performance Traits in Cattle and Sheep" Animals 13, no. 15: 2470. https://doi.org/10.3390/ani13152470
APA StyleOster, N., Szewczuk, M. A., Zych, S., Stankiewicz, T., Błaszczyk, B., & Wieczorek-Dąbrowska, M. (2023). Association between Polymorphism in the Janus Kinase 2 (JAK2) Gene and Selected Performance Traits in Cattle and Sheep. Animals, 13(15), 2470. https://doi.org/10.3390/ani13152470