



Functional Additives in a Selected European Sea Bass (Dicentrarchus labrax) Genotype: Effects on the Stress Response and Gill Antioxidant Response to Hydrogen Peroxide (H2O2) Treatment

 , , , , , and
, , , , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Population Design and Fish Production
2.3. Experimental Conditions
2.4. Oxidative Stress Challenge
2.5. Sampling Methodology
2.6. RNA Extraction and Real-Time PCR Analysis
2.7. Statistical Analyses
3. Results
3.1. Feeding Experience
3.2. Stress Response
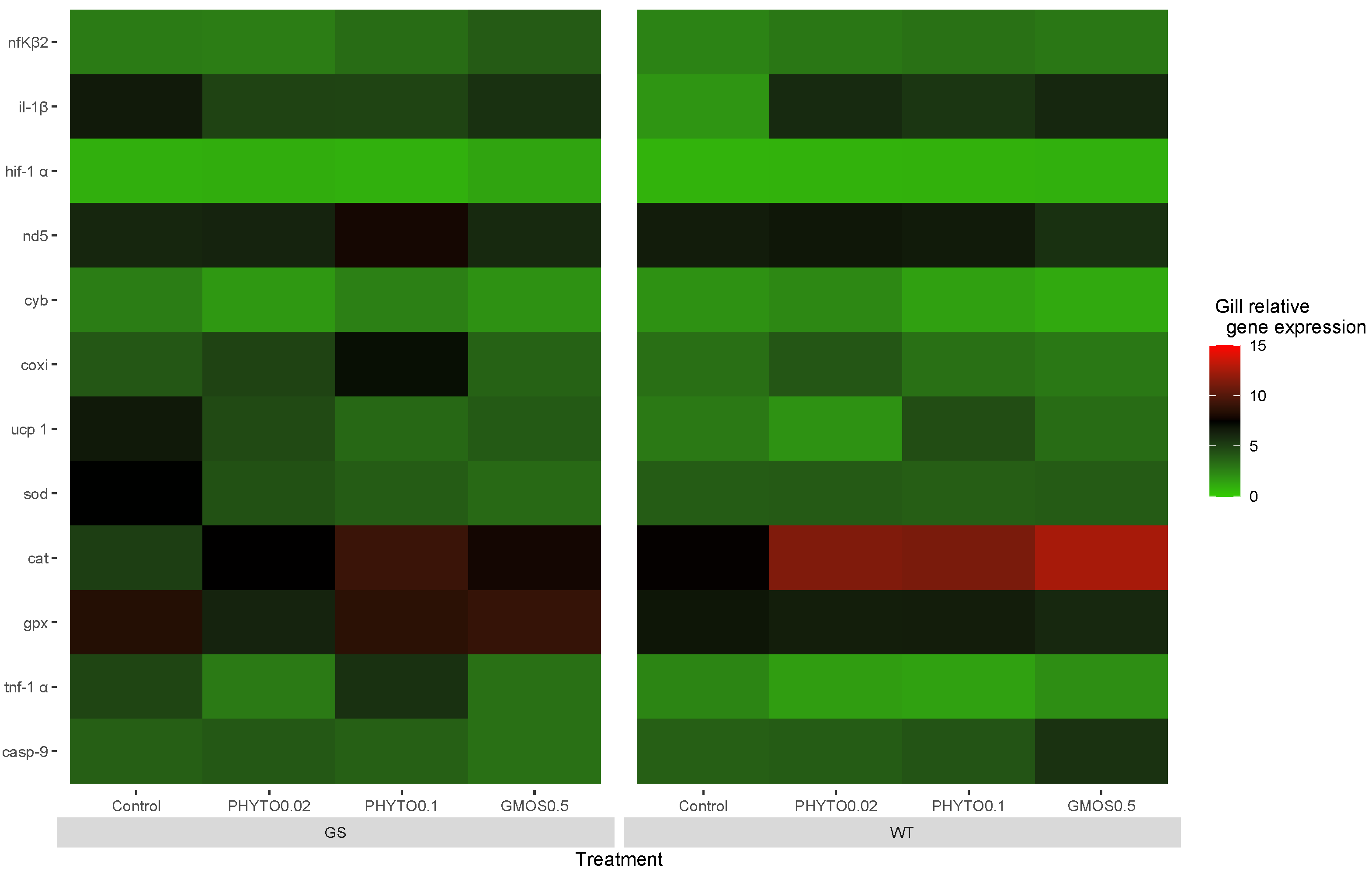
3.3. Gill Relative Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| High-Growth Selected Genotype (GS) | Wild Type Genotype (WT) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | ||
| Sampling Point | Target Gene | ||||||||
| t = 0 h (pre-H2O2 exposure) | nfΚβ2 | 0.89 1 ± 0.26 | 1.36 1 ± 0.19 | 1.17 1 ± 0.13 | 1.49 1 ± 0.35 | 1.02 1 ± 0.17 | 0.90 1 ± 0.14 | 1.41 1 ± 0.48 | 0.98 1 ± 0.58 |
| il-1β | 2.58 ± 1.04 | 1.94 ± 0.43 | 1.94 ± 0.45 | 2.48 ± 0.48 | 0.95 1 ± 0.33 | 0.93 1 ± 0.33 | 1.42 1 ± 0.49 | 0.93 1 ± 0.23 | |
| hif-1α | 0.60 ± 0.16 | 0.64 ± 0.07 | 0.45 ± 0.19 | 0.33 a1 ± 0.20 | 0.78 A ± 0.41 | 1.22 AB ± 0.44 | 0.73 A ± 0.13 | 1.61 bB ± 0.16 | |
| nd5 | 1.42 ± 0.76 | 1.24 ± 0.31 | 1.01 1 ± 0.14 | 1.34 ± 0.41 | 0.89 1 ± 0.13 | 1.05 1 ± 0.16 | 1.28 1 ± 0.15 | 0.85 ± 0.31 | |
| cyb | 1.32 ± 0.07 | 0.53 ± 0.11 | 0.54 1 ±0.18 | 0.46 ± 0.18 | 0.92 ± 0.20 | 1.20 ± 1.12 | 1.50 ± 0.55 | 1.39 ± 0.63 | |
| cox | 1.16 1 ± 0.11 | 1.63 1 ± 0.42 | 1.59 1 ± 0.46 | 0.91 1 ± 0.21 | 0.84 1 ± 0.11 | 1.45 1 ± 0.32 | 1.56 1 ± 0.47 | 1.01 1 ± 0.50 | |
| ucp1 | 1.46 1 ± 0.65 | 2.07 ± 1.27 | 2.39 ± 1.24 | 2.98 ± 1.52 | 0.75 ± 0.09 | 0.53 ± 0.07 | 1.10 1 ± 0.20 | 0.68 ± 0.30 | |
| sod | 1.06 a1± 0.23 | 1.32 1 ± 0.20 | 1.24 1 ± 0.27 | 1.44 a1± 0.17 | 0.81 bA1 ± 0.23 | 2.14 B1 ± 0.19 | 2.00 AB1 ± 1.13 | 4.28 bC ± 0.88 | |
| cat | 3.40 a1± 0.62 | 4.59 1 ± 0.86 | 3.97 a1 ± 0.34 | 3.44 1 ± 0.50 | 0.85 b1 ± 0.26 | 2.16 1 ± 1.28 | 1.25 b1 ± 0.45 | 1.60 1 ± 0.74 | |
| gpx | 1.62 1 ± 0.46 | 1.85 1 ± 0.29 | 1.88 1 ± 0.26 | 2.00 1 ± 0.36 | 0.85 A1 ± 0.18 | 1.80 A1 ± 0.06 | 3.21 C1 ± 1.59 | 2.55 B1 ± 1.27 | |
| tnf-1α | 0.55 1 ± 0.55 | 0.29 ± 0.11 | 0.28 1 ± 0.07 | 0.30 1 ± 0.28 | 0.96 ± 0.28 | 1.03 ± 0.62 | 1.15 ± 0.92 | 2.32 ± 0.80 | |
| casp-9 | 1.69 ± 1.68 | 1.36 ± 0.24 | 1.33 ± 0.34 | 1.46 ± 0.21 | 0.82 ± 0.26 | 0.98 ± 0.17 | 1.70 ± 0.58 | 1.01 1 ± 0.34 | |
| t = 2 h (post-H2O2 exposure) | nfΚβ2 | 2.75 2 ± 0.39 | 2.66 2 ± 0.33 | 3.34 2 ± 0.32 | 3.90 2 ± 0.96 | 2.55 2 ± 0.25 | 2.92 2 ± 0.40 | 3.08 2 ± 0.58 | 2.95 2 ± 0.10 |
| il-1β | 5.75 ± 4.14 | 4.80 ± 1.42 | 4.78 ± 1.28 | 5.65 ± 0.62 | 5.47 12 ± 0.19 | 5.66 2 ± 2.25 | 5.38 ± 1.04 | 6.06 2 ± 0.79 | |
| hif-1α | 0.96 ± 0.12 | 1.02 ± 0.14 | 0.94 ± 0.11 | 1.29 2 ± 0.41 | 0.84 ± 0.13 | 0.88 ± 0.06 | 0.89 ± 018 | 0.93 ± 0.06 | |
| nd5 | 6.03 ± 1.46 | 5.99 ± 2.15 | 6.69 2 ± 5.13 | 5.95 ± 1.08 | 6.56 2 ± 0.34 | 6.70 2 ± 1.50 | 6.62 2 ± 0.52 | 5.55 ± 1.35 | |
| cyb | 2.59 ± 0.86 | 1.72 ± 0.48 | 2.59 2 ± 0.23 | 1.90 ± 0.82 | 1.89 ± 0.87 | 2.21 ± 0.87 | 1.47 ± 0.25 | 1.15 ± 0.30 | |
| cox | 4.13 AB2 ± 0.41 | 4.77 AB2 ± 1.56 | 7.12 aB2 ± 0.45 | 3.69 A2 ± 0.46 | 3.21 2 ± 0.49 | 4.10 2 ± 0.94 | 3.20 b12 ± 0.33 | 2.84 12 ± 0.80 | |
| ucp1 | 6.65 2 ± 1.29 | 4.38 ± 1.93 | 2.94 ± 2.33 | 3.72 ± 1.94 | 2.56 ± 1.54 | 1.84 ± 0.90 | 3.72 12 ± 3.14 | 3.30 ± 0.72 | |
| sod | 7.45 aA2 ± 0.84 | 4.38 B2 ± 0.27 | 3.90 B2 ± 0.82 | 3.41 B2 ± 0.66 | 3.88 b2 ± 0.83 | 4.01 2 ± 0.41 | 3.85 2 ± 0.30 | 3.98 ± 0.37 | |
| cat | 4.97 A2 ± 1.62 | 7.51 AB12 ± 0.49 | 9.18 B2 ± 0.81 | 7.86 AB2 ± 0.42 | 7.39 A2 ± 2.05 | 11.29 AB2 ± 2.06 | 11.23 AB2 ± 1.23 | 12.58 B2 ± 0.11 | |
| gpx | 8.34 2± 1.41 | 6.24 2 ± 0.18 | 8.62 2 ± 1.21 | 8.99 2 ± 0.79 | 6.83 2 ± 0.84 | 6.47 2 ± 0.80 | 6.44 2 ± 1.29 | 6.00 2 ± 0.70 | |
| tnf-1α | 4.70 2 ± 1.34 | 2.47 ± 1.68 | 4.49 2 ± 4.24 | 2.45 12 ± 2.51 | 2.27 ± 1.23 | 1.55 ± 0.51 | 1.42 ± 0.39 | 2.02 ± 0.93 | |
| casp-9 | 3.32 ± 2.36 | 3.64 ± 2.30 | 3.50 ± 1.73 | 2.98 ± 1.33 | 3.72 ± 1.00 | 3.93 ± 0.59 | 4.24 ± 0.51 | 5.34 2 ± 2.14 | |
| t = 24 h (post-H2O2 exposure) | nfΚβ2 | 3.20 2 ± 0.17 | 3.20 2 ± 0.24 | 3.21 2 ± 0.55 | 3.69 a2 ± 0.17 | 2.42 2 ± 0.45 | 2.64 2 ± 0.77 | 2.91 2 ± 0.39 | 2.37 b2 ± 0.15 |
| il-1β | 3.50 ± 0.24 | 3.42 ± 0.22 | 4.88 ± 1.30 | 5.43 ± 1.79 | 3.81 2 ± 1.60 | 5.27 2 ± 2.50 | 4.72 ± 1.20 | 3.73 2 ± 1.43 | |
| hif-1α | 0.91 ± 0.30 | 1.09 ± 0.13 | 0.90 ± 0.15 | 1.24 2 ± 0.09 | 0.94 ± 0.15 | 1.00 ± 0.23 | 0.92 ± 0.14 | 0.93 ± 0.37 | |
| nd5 | 6.05 ± 1.40 | 6.99 ± 1.43 | 3.22 12 ± 1.41 | 6.28 ± 3.56 | 4.45 12 ± 1.31 | 4.45 12 ± 1.11 | 4.44 12 ± 1.44 | 3.70 ± 0.72 | |
| cyb | 1.61 ± 0.28 | 1.53 ± 0.19 | 1.63 12 ± 0.21 | 1.55 ± 0.41 | 1.27 ± 0.43 | 1.41 ± 0.43 | 1.69 ± 0.36 | 2.00 ± 0.86 | |
| cox | 4.88 2 ± 0.71 | 4.33 2 ± 0.47 | 4.05 3 ± 0.33 | 5.47 2 ± 2.84 | 3.30 2 ± 0.71 | 4.00 2 ± 1.03 | 3.92 2 ± 1.36 | 3.14 2 ± 0.22 | |
| ucp1 | 3.42 12 ± 1.82 | 2.31 ± 1.01 | 2.29 ± 0.54 | 3.90 ± 1.55 | 2.99 AB ± 0.58 | 3.22 AB ± 1.43 | 6.96 A2 ± 1.23 | 1.54 B ± 0.74 | |
| sod | 3.60 3 ± 0.66 | 3.35 2 ± 0.30 | 3.64 2 ± 0.43 | 3.41 2 ± 0.28 | 3.93 2 ± 0.57 | 4.16 2 ± 0.68 | 4.32 2 ± 0.38 | 4.77 ± 0.68 | |
| cat | 8.69 2 ± 1.45 | 9.61 2 ± 2.34 | 10.30 2 ± 1.05 | 9.77 2 ± 1.17 | 6.80 2 ± 0.72 | 7.87 2 ± 2.22 | 7.42 2 ± 1.50 | 7.31 3 ± 1.47 | |
| gpx | 8.82 2 ± 0.94 | 8.74 2 ± 1.12 | 8.49 2 ± 0.45 | 7.89 2 ± 0.86 | 5.69 2 ± 0.84 | 5.5 2 ± 1.10 | 6.76 2 ± 0.73 | 6.68 2 ± 0.79 | |
| tnf-1α | 2.44 12 ± 1.08 | 2.04 ± 0.81 | 2.41 12 ± 0.94 | 3.82 2 ± 1.53 | 1.68 ± 0.40 | 2.04 ± 0.89 | 3.28 ± 1.65 | 1.23 ± 0.34 | |
| casp-9 | 3.96 ± 2.26 | 4.06 ± 1.28 | 4.47 ± 1.32 | 5.83 ± 3.45 | 2.88 ± 0.38 | 2.54 ± 0.62 | 2.39 ± 0.33 | 2.63 12 ± 0.30 | |
| Three-way ANOVA | |||||||||
| Diet | Genotype | Time | D × G | D × T | G × T | D × G × T | |||
| nfΚβ2 | F = 4.306 p-val = 0.009 | F = 16.398 p-val = 0.0018 | F = 158.176 p-val = < 2 × 10−16 | F = 3.481 p-val = 0.0229 | ns | F = 3.335 p-val = 0.0474 | ns | ||
| il-1β | ns | F = 8.798 p-val = 0.0047 | F = 65.499 p-val = 1.91 × 10−14 | ns | ns | ns | ns | ||
| hif-1α | F = 5.231 p-val = 0.0033 | F = 4.422 p-val = 0.041 | F = 5.714 p-val = 0.006 | ns | ns | F = 21.397 p-val = 2.27 × 10−7 | F = 4.260 p-val = 0.00162 | ||
| nd5 | ns | ns | F = 63.876 p-val = 2.97 × 10−14 | ns | ns | ns | ns | ||
| cyb | ns | ns | F = 19.325 p-val = 6.97 × 10−7 | ns | ns | F = 5.819 p-val = 0.005 | ns | ||
| cox | F = 4.108 p-val = 0.0113 | F = 21.469 p-val = 2.77 × 10−5 | F = 127.517 p-val = < 2 × 10−16 | ns | ns | F = 4.051 p-val = 0.0237 | F = 2.616 p-val = 0.028 | ||
| ucp1 | ns | F = 7.93 p-val = 0.007 | F = 19.895 p-val = 5.09 × 10−7 | F = 4.471 p-val = 0.007 | ns | F = 5.287 p-val = 0.008 | F = 2.699 p-val = 0.024 | ||
| sod | ns | F = 10.086 p-val = 0.002 | F = 181.372 p-val = < 2 × 10−16 | F = 17.654 p-val = 7.36 × 10−8 | F = 13.457 p-val = 6.98 × 10−9 | F = 19.065 p-val = 8.06 × 10−7 | F = 2.748 p-val = 0.022 | ||
| cat | F = 9.911 p-val = 3.38 × 10−5 | F = 10.017 p-val = 0.002 | F = 242.091 p-val = < 2 × 10−16 | ns | F = 2.914 p-val = 0.017 | F = 44.054 p-val = 1.37 × 10−11 | ns | ||
| gpx | F = 2.859 p-val = 0.046 | F = 35.012 p-val = 3.36 × 107 | F = 294.593 p-val = < 2 × 10−16 | ns | ns | F = 13.893 p-val = 1.74 × 10−5 | F = 3.079 p-val = 0.013 | ||
| tnf-1α | ns | ns | F = 25.759 p-val = 2.51 × 10−8 | ns | ns | F = 10.79 p-val = 0.0001 | ns | ||
| casp-9 | ns | ns | F = 39.156 p-val = 8.23 × 10−11 | ns | ns | F = 5.946 p-val = 0.005 | ns | ||
References
- Acosta, F.; Montero, D.; Izquierdo, M.; Galindo-Villegas, J. High-level biocidal products effectively eradicate pathogenic γ-proteobacteria biofilms from aquaculture facilities. Aquaculture 2021, 532, 736004. [Google Scholar] [CrossRef]
- Magara, G.; Sangsawang, A.; Pastorino, P.; Oddon, S.B.; Caldaroni, B.; Menconi, V.; Kovitvadhi, U.; Gasco, L.; Meloni, D.; Dörr, A.J.M.; et al. First insights into oxidative stress and theoretical environmental risk of Bronopol and Detarox® AP, two biocides claimed to be ecofriendly for a sustainable aquaculture. Sci. Total Environ. 2021, 778, 146375. [Google Scholar] [CrossRef] [PubMed]
- Avendaño-Herrera, R.; Magariños, B.; Irgang, R.; Toranzo, A.E. Use of hydrogen peroxide against the fish pathogen Tenacibaculum maritimum and its effect on infected turbot (Scophthalmus maximus). Aquaculture 2006, 257, 104–110. [Google Scholar] [CrossRef]
- Pedersen, L.F. Investigations of Environmental Benign Aquaculture Therapeutics Replacing Formalin; DTU Aqua Report, no. 218–2010; DTU Aqua: Charlottenlund, Denmark, 2010. [Google Scholar]
- Hodkovicova, N.; Chmelova, L.; Sehonova, P.; Blahova, J.; Doubkova, V.; Plhalova, L.; Fiorino, E.; Vojtek, L.; Vicenova, M.; siroka, Z.; et al. The effects of a therapeutic formalin bath on selected immunological and oxidative stress parameters in common carp (Cyprinus carpio). Sci. Total Environ. 2019, 653, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Seoud, S.S.M.; Zaki, V.H.; Ahmed, G.E.; Abd El-Khalek, N.K. Studies on Amyloodinium infestation in European seabass (Dicentrarchus labrax) fishes with special reference for treatment. Int. J. Mar. Sci. 2017, 7. [Google Scholar] [CrossRef]
- Ragab, R.H.; Elgendy, M.Y.; Sabry, N.M.; Sharaf, M.S.; Attia, M.M.; Korany, R.M.; Abdelsalam, M.; Eltahan, A.S.; Eldessouki, E.A.; El-Demerdash, G.O.; et al. Mass kills in hatchery-reared European seabass (Dicentrarchus labrax) triggered by concomitant infections of Amyloodinium ocellatum and Vibrio alginolyticus. Int. J. Vet. Sci. Med. 2022, 10, 33–45. [Google Scholar] [CrossRef]
- Roque, A.; Yildiz, H.Y.; Carazo, I.; Duncan, N. Physiological stress responses of sea bass (Dicentrarchus labrax) to hydrogen peroxide (H2O2) exposure. Aquaculture 2010, 304, 104–107. [Google Scholar] [CrossRef]
- Seker, E.; Ispir, U.; Yonar, S.M.; Yonar, M.E.; Turk, C. Antioxidant responses of rainbow trout (Oncorhynchus mykiss) gills after exposure to hydrogen peroxide. Fresenius Environ. Bull. 2015, 24, 1837–1840. [Google Scholar]
- Hwang, P.-P.; Lee, T.-H.; Lin, L.-Y. Ion regulation in fish gills: Recent progress in the cellular and molecular mechanisms. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2011, 301, R28–R47. [Google Scholar] [CrossRef]
- Li, M.; Zhu, X.; Tian, J.; Liu, M.; Wang, G. Dietary flavonoids from Allium mongolicum Regel promotes growth, improves immune, antioxidant status, immune-related signaling molecules and disease resistance in juvenile northern snakehead fish (Channa argus). Aquaculture 2019, 501, 473–481. [Google Scholar] [CrossRef]
- Rodnick, K.J.; Planas, J.V. The stress and stress mitigation effects of exercise: Cardiovascular, metabolic, and skeletal muscle adjustments. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 251–294. [Google Scholar] [CrossRef]
- Gorissen, M.; Flik, G. The endocrinology of the stress response in fish: An adaptation-physiological view. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 75–111. [Google Scholar] [CrossRef]
- Spiers, J.G.; Chen, H.-J.C.; Sernia, C.; Lavidis, N.A. Activation of the hypothalamic-pituitary-adrenal stress axis induces cellular oxidative stress. Front. Neurosci. 2015, 8, 456. [Google Scholar] [CrossRef]
- Zhao, R.-Z.; Jiang, S.; Zhang, L.; Yu, Z.-B. Mitochondrial electron transport chain, ROS generation and uncoupling. Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Zafir, A.; Banu, N. Modulation of in vivo oxidative status by exogenous corticosterone and restraint stress in rats. Stress 2009, 12, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Fanouraki, E.; Mylonas, C.C.; Papandroulakis, N.; Pavlidis, M. Species specificity in the magnitude and duration of the acute stress response in Mediterranean marine fish in culture. Gen. Comp. Endocrinol. 2011, 173, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Samaras, A.; Pavlidis, M.; Lika, K.; Theodoridi, A.; Papandroulakis, N. Scale matters: Performance of European sea bass, Dicentrarchus labrax, L. (1758), reared in cages of different volumes. Aquac. Res. 2017, 48, 990–1005. [Google Scholar] [CrossRef]
- Hematyar, N.; Rustad, T.; Sampels, S.; Kastrup Dalsgaard, T. Relationship between lipid and protein oxidation in fish. Aquac. Res. 2019, 50, 1393–1403. [Google Scholar] [CrossRef]
- Irkin, L.C.; Yigit, M.; Yilmaz, S.; Maita, M. Toxicological Evaluation of Dietary Garlic (Allium sativum) Powder in European Sea Bass Dicentrarchus labrax Juveniles. Food Nutr. Sci. 2014, 5, 46429. [Google Scholar] [CrossRef]
- Yonar, M.E.; Yonar, S.M.; İspir, Ü.; Ural, M.Ş. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. achromogenes. Fish Shellfish Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative stress and antioxidant defense in fish: The implications of probiotic, prebiotic, and synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Singh, J.; Gaikwad, D.S. Phytogenic feed additives in animal nutrition. In Natural Bioactive Products in Sustainable Agriculture; Springer: Singapore, 2020; pp. 273–289. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Suharman, I.; Avillanosa, A.L.; Gonzales-Plasus, M.M. Influence of phytogenic feed additives on the health status in the gut and disease resistance of cultured fish. IOP Conf. Ser. Earth Environ. Sci. 2021, 695, 012024. [Google Scholar] [CrossRef]
- Choubey, M.; Pattanaik, A.K.; Baliyan, S.; Dutta, N.; Jadhav, S.E.; Sharma, K. Dietary supplementation of a novel phytogenic feed additive: Effects on nutrient metabolism, antioxidant status and immune response of goats. Anim. Prod. Sci. 2015, 56, 1612–1621. [Google Scholar] [CrossRef]
- Serradell, A.; Torrecillas, S.; Makol, A.; Acosta, F.; Valdenegro, V.; Montero, D. Functional additives in low fish meal and fish oil based diets for European sea bass (Dicentrarchus labrax): Effects on immune response, stress and disease resistance. Fish Shellfish Immunol. 2020, 91, 464–465. [Google Scholar] [CrossRef]
- Ali, A.; Ponnampalam, E.N.; Pushpakumara, G.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Cinnamon: A natural feed additive for poultry health and production—A review. Animals 2021, 11, 2026. [Google Scholar] [CrossRef]
- Cho, H.A.; Song, M.H.; Lee, J.H.; Oh, H.J.; Kim, Y.J.; An, J.W.; Chang, S.Y.; Go, Y.B.; Song, D.C.; Cho, S.Y.; et al. Effects of different stocking density and various phytogenic feed additives dosage levels on growing-finishing pigs. J. Anim. Sci. Technol. 2023, 65, 535–549. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef]
- Rimoldi, S.; Torrecillas, S.; Montero, D.; Gini, E.; Makol, A.; Valdenegro V., V.; Izquierdo, M.; Terova, G. Assessment of dietary supplementation with galactomannan oligosaccharides and phytogenics on gut microbiota of European sea bass (Dicentrarchus labrax) fed low fishmeal and fish oil based diet. PLoS ONE 2020, 15, e0231494. [Google Scholar] [CrossRef] [PubMed]
- Torrecillas, S.; Terova, G.; Makol, A.; Serradell, A.; Valdenegro, V.; Gini, E.; Izquierdo, M.S.; Acosta, F.; Montero, D. Dietary phytogenics and galactomannan oligosaccharides in low fish meal and fish oil-based diets for European sea bass (Dicentrarchus labrax) juveniles: Effects on gut health and implications on in vivo gut bacterial translocation. PLoS ONE 2019, 14, e0222063. [Google Scholar] [CrossRef]
- Torrecillas, S.; Terova, G.; Makol, A.; Serradell, A.; Valdenegro-Vega, V.; Izquierdo, M.; Acosta, F.; Montero, D. Dietary phytogenics and galactomannan oligosaccharides in low fish meal and fish oil-based diets for European sea bass (Dicentrarchus labrax) juveniles: Effects on gill structure and health and implications on oxidative stress status. Front. Immunol. 2021, 12, 663106. [Google Scholar] [CrossRef]
- Guerreiro, I.; Serra, C.R.; Enes, P.; Couto, A.; Salvador, A.; Costas, B.; Oliva-Teles, A. Effect of short chain fructooligosaccharides (scFOS) on immunological status and gut microbiota of gilthead sea bream (Sparus aurata) reared at two temperatures. Fish Shellfish Immunol. 2016, 49, 122–131. [Google Scholar] [CrossRef]
- Torrecillas, S.; Rivero-Ramírez, F.; Izquierdo, M.S.; Caballero, M.J.; Makol, A.; Suarez-Bregua, P.; Fernández-Montero, A.; Rotllant, J.; Montero, D. Feeding European sea bass (Dicentrarchus labrax) juveniles with a functional synbiotic additive (mannan oligosaccharides and Pediococcus acidilactici): An effective tool to reduce low fishmeal and fish oil gut health effects? Fish Shellfish Immunol. 2018, 81, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Encarnação, P. Functional feed additives in aquaculture feeds. In Aquafeed Formulation; Academic Press: Cambridge, MA, USA, 2016; pp. 217–237. [Google Scholar] [CrossRef]
- Vandeputte, M.; Dupont-Nivet, M.; Haffray, P.; Chavanne, H.; Cenadelli, S.; Parati, K.; Vidal, M.-O.; Vergnet, A.; Chatain, B. Response to domestication and selection for growth in the European sea bass (Dicentrarchus labrax) in separate and mixed tanks. Aquaculture 2009, 286, 20–27. [Google Scholar] [CrossRef]
- Kause, A.; Kiessling, A.; Martin, S.A.M.; Houlihan, D.; Ruohonen, K. Genetic improvement of feed conversion ratio via indirect selection against lipid deposition in farmed rainbow trout (Oncorhynchus mykiss Walbaum). Br. J. Nutr. 2016, 116, 1656–1665. [Google Scholar] [CrossRef]
- Vandeputte, M.; Gagnaire, P.-A.; Allal, F. The European sea bass: A key marine fish model in the wild and in aqua-culture. Anim. Genet. 2019, 50, 195–206. [Google Scholar] [CrossRef]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed. Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Montero, D.; Carvalho, M.; Terova, G.; Fontanillas, R.; Serradell, A.; Ginés, R.; Tuset, V.; Acosta, F.; Rimoldi, S.; Bajek, A.; et al. Nutritional innovations in superior European sea bass (Dicentrarchus labrax) genotypes: Implications on fish performance and feed utilization. Aquaculture 2023, 572, 739486. [Google Scholar] [CrossRef]
- Tort, L. Stress and immune modulation in fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Kousoulaki, K.; Sæther, B.-S.; Albrektsen, S.; Noble, C. Review on European sea bass (Dicentrarchus labrax, Linnaeus, 1758) nutrition and feed management: A practical guide for optimizing feed formulation and farming protocols. Aquac. Nutr. 2015, 21, 129–151. [Google Scholar] [CrossRef]
- Torrecillas, S.; Rimoldi, S.; Montero, D.; Serradell, A.; Acosta, F.; Fontanillas, R.; Allal, F.; Haffray, P.; Bajek, A.; Terova, G. Genotype x nutrition interactions in European sea bass (Dicentrarchus labrax): Effects on gut health and intestinal microbiota. Aquaculture 2023, 574, 739639. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Socorro, J.; Roo, J. Studies on the appearance of skeletal anomalies in red porgy: Effect of culture intensiveness, feeding habits and nutritional quality of live preys. J. Appl. Ichthyol. 2010, 26, 320–326. [Google Scholar] [CrossRef]
- Atalah, E.; Hernández-Cruz, C.M.; Ganuza, E.; Benítez-Santana, T.; Ganga, R.; Roo, J.; Montero, D.; Izquierdo, M.S. Importance of dietary arachidonic acid for survival, growth and stress resistance of larval European sea bass (Dicentrarchus labrax) fed high dietary docosahexaenic and eicosapentaenoic acids. Aquac. Res. 2011, 42, 1261–1268. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Azeredo, R.; Pérez-Sánchez, J.; Sitjà-Bobadilla, A.; Fouz, B.; Tort, L.; Aragão, C.; Oliva-Teles, A.; Costas, B. European Sea Bass (Dicentrarchus labrax) Immune Status and Disease Resistance Are Impaired by Arginine Dietary Supplementation. PLoS ONE 2015, 10, e0139967. [Google Scholar] [CrossRef] [PubMed]
- Román, L.; Real, F.; Padilla, D.; El Aamri, F.; Déniz, S.; Grasso, V.; Acosta, F. Cytokine expression in head-kidney leucocytes of European sea bass (Dicentrarchus labrax L.) after incubation with the probiotic Vagococcus fluvialis L-21. Fish Shellfish Immunol. 2013, 35, 1329–1332. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Corà, S.; Bernardini, G.; Gornati, R.; Saroglia, M. Acute and chronic hypoxia affects HIF-1α mRNA levels in sea bass (Dicentrarchus labrax). Aquaculture 2008, 279, 150–159. [Google Scholar] [CrossRef]
- Brandts, I.; Teles, M.; Tvarijonaviciute, A.; Pereira, M.L.; Martins, M.A.; Tort, L.; Oliveira, M. Effects of polymethylmethacrylate nanoplastics on Dicentrarchus labrax. Genomics 2018, 110, 435–441. [Google Scholar] [CrossRef]
- Basto, A.; Calduch-Giner, J.; Oliveira, B.; Petit, L.; Sá, T.; Maia, M.R.G.; Fonseca, S.C.; Matos, E.; Pérez-Sánchez, J.; Valente, L.M.P. The Use of Defatted Tenebrio molitor Larvae Meal as a Main Protein Source Is Supported in European Sea Bass (Dicentrarchus labrax) by Data on Growth Performance, Lipid Metabolism, and Flesh Quality. Front. Physiol. 2023, 12, 659567. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Ramírez, D.; Mazurais, D.; Gatesoupe, J.F.; Quazuguel, P.; Cahu, C.L.; Zambonino-Infante, J.L. Dietary probiotic live yeast modulates antioxidant enzyme activities and gene expression of sea bass (Dicentrarchus labrax) larvae. Aquaculture 2010, 300, 142–147. [Google Scholar] [CrossRef]
- Reis, M.I.R.; Nascimento, D.S.; do Vale, A.; Silva, M.T.; dos Santos, N.M.S. Molecular cloning and characterisation of sea bass (Dicentrarchus labrax L.) caspase-3 gene. Mol. Immunol. 2007, 44, 774–783. [Google Scholar] [CrossRef]
- Bodur, T.; León-Bernabeu, S.; Navarro, A.; Tort, L.; Afonso, J.M.; Montero, D. Effects of new plant based anesthetics Origanum sp. and Eucalyptus sp. oils on stress and welfare parameters in Dicentrarchus labrax and their comparison with clove oil. Aquaculture 2018, 495, 402–408. [Google Scholar] [CrossRef]
- Hoaglin, D.C.; Iglewicz, B. Fine-tuning some resistant rules for outlier labeling. J. Am. Stat. Assoc. 1987, 82, 1147–1149. [Google Scholar] [CrossRef]
- Feng, M.; Ll, Q.; Zou, Z. An outlier identification and judgment method for an improved neural-network BOF forecasting model. Steel Res. Int. 2008, 79, 323–332. [Google Scholar] [CrossRef]
- Underwood, A.J. Experiments in Ecology: Their Logical Designand Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997; 504p. [Google Scholar]
- Jensen, P. Behavior genetics and the domestication of animals. Annu. Rev. Anim. Biosci. 2014, 2, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, M.; Porte, J.D.; Auperin, B.; Dupont-Nivet, M.; Vergnet, A.; Valotaire, C.; Claireaux, G.; Prunet, P.; Chatain, B. Quantitative genetic variation for post-stress cortisol and swimming performance in growth-selected and control populations of European sea bass (Dicentrarchus labrax). Aquaculture 2016, 455, 1–7. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The Concept of Stress In Fish. In Biology of Stress in Fish; Schreck, C.B., Tort, L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–34. [Google Scholar] [CrossRef]
- Øverli, Ø.; Sørensen, C.; Kiessling, A.; Pottinger, T.G.; Gjøen, H.M. Selection for improved stress tolerance in rainbow trout (Oncorhynchus mykiss) leads to reduced feed waste. Aquaculture 2006, 261, 776–781. [Google Scholar] [CrossRef]
- Solberg, M.F.; Kvamme, B.O.; Nilsen, F.; Glover, K.A. Effects of environmental stress on mRNA expression levels of seven genes related to oxidative stress and growth in Atlantic salmon Salmo salar L. of farmed, hybrid and wild origin. BMC Res. Notes 2012, 5, 672. [Google Scholar] [CrossRef]
- Sauvage, C.; Derome, N.; Normandeau, E.; St.-Cyr, J.; Audet, C.; Bernatchez, L. Fast transcriptional responses to domestication in the brook charr Salvelinus fontinalis. Genetics 2010, 185, 105–112. [Google Scholar] [CrossRef]
- Palińska-Żarska, K.; Król, J.; Woźny, M.; Kamaszewski, M.; Szudrowicz, H.; Wiechetek, W.; Brzuzan, P.; Fopp-Bayat, D.; Żarski, D. Domestication affected stress and immune response markers in Perca fluviatilis in the early larval stage. Fish Shellfish Immunol. 2021, 114, 184–198. [Google Scholar] [CrossRef]
- Li, C.; Tang, B.; Feng, Y.; Tang, F.; Pui-Man Hoi, M.; Su, Z.; Ming-Yuen Lee, S. Pinostrobin exerts neuroprotective actions in neurotoxin-induced parkinson’s disease models through Nrf2 induction. J. Agric. Food Chem. 2018, 66, 8307–8318. [Google Scholar] [CrossRef]
- Mansour, A.T.; Espinosa, C.; García-Beltrán, J.M.; Miao, L.; Ceballos Francisco, D.C.; Alsaqufi, A.S.; Esteban, M. Dietary supplementation of drumstick tree, Moringa oleifera, improves mucosal immune response in skin and gills of seabream, Sparus aurata, and attenuates the effect of hydrogen peroxide exposure. Fish Physiol. Biochem. 2020, 46, 981–996. [Google Scholar] [CrossRef]
- Aedo, J.E.; Fuentes-Valenzuela, M.; Molina, A.; Valdés, J.A. Quantitative proteomics analysis of membrane glucocorticoid receptor activation in rainbow trout skeletal muscle. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 32, 100627. [Google Scholar] [CrossRef]
- Busiello, R.A.; Savarese, S.; Lombardi, A. Mitochondrial uncoupling proteins and energy metabolism. Front. Physiol. 2015, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Biswas, R.; Bagchi, A. NFkB pathway and inhibition: An overview. Comput. Mol. Biol. 2016, 6, 1–20. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.-H.; Chen, X.-Y.; Hu, Q.-H.; Wang, M.-X.; Jin, R.; Zhang, Q.-Y.; Wang, W.; Wang, R.; Kang, L.-L.; et al. Reactive oxygen species-induced TXNIP drives fructose-mediated hepatic inflammation and lipid accumulation through NLRP3 inflammasome activation. Antioxid. Redox Signal. 2015, 22, 848–870. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef]
- Lazado, C.C.; Voldvik, V.; Breiland, M.W.; Osório, J.; Hansen, M.H.S.; Krasnov, A. Oxidative chemical stressors alter the physiological state of the nasal olfactory mucosa of Atlantic salmon. Antioxidants 2020, 9, 1144. [Google Scholar] [CrossRef] [PubMed]
- Kierans, S.J.; Taylor, C.T. Regulation of glycolysis by the hypoxia-inducible factor (HIF): Implications for cellular physiology. J. Physiol. 2021, 599, 23–37. [Google Scholar] [CrossRef]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013, 14, 32. [Google Scholar] [CrossRef]



| Ingredients | Diet (%) | |||
|---|---|---|---|---|
| Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | |
| Fish meal 1 | 9.6 | 9.6 | 9.6 | 9.6 |
| Soya protein concentrate | 18.2 | 18.2 | 18.2 | 18.2 |
| Soya meal | 11.6 | 11.6 | 11.6 | 11.6 |
| Corn gluten meal | 24.1 | 24.1 | 24.1 | 24.1 |
| Wheat | 8.585 | 8.565 | 8.485 | 8.085 |
| Wheat gluten | 1.9 | 1.9 | 1.9 | 1.9 |
| Guar meal | 7.7 | 7.7 | 7.7 | 7.7 |
| Rapeseed extracted | 3.0 | 3.0 | 3.0 | 3.0 |
| Fish oil 2 | 6.5 | 6.5 | 6.5 | 6.5 |
| Rapeseed oil 3 | 5.2 | 5.2 | 5.2 | 5.2 |
| Vitamin and mineral premix 4 | 3.6 | 3.6 | 3.6 | 3.6 |
| Antioxidant 5 | 0.015 | 0.015 | 0.015 | 0.015 |
| Phytogenic (garlic and Labiatae plant essential oils) 6 | 0 | 0.02 | 0 | 0 |
| Phytogenic (citrus fruits and Asteraceae and Labiatae plant essential oils) 7 | 0 | 0 | 0.1 | 0 |
| Galactomannan-oligosaccharides (GMOS) 8 | 0 | 0 | 0 | 0.5 |
| Proximate composition (% of dry matter) | ||||
| Crude lipids | 19.91 | 20.44 | 20.44 | 20.47 |
| Crude protein | 49.30 | 49.27 | 49.27 | 49.76 |
| Moisture | 5.10 | 5.01 | 5.01 | 5.06 |
| Ash | 7.02 | 6.41 | 6.41 | 6.49 |
| Gene | Access. Number | Primer | Nucleotide Sequence 5′-3′ | Annealing T (°C) | Reference |
|---|---|---|---|---|---|
| nfΚβ2 | KM225790 | Fw | CTGGAGGAAACTGGCGGAGAAGC | 60 | [49] |
| Rv | CAGGTACAGGTGAGTCAGCGTCATC | ||||
| il-1b | AJ53742 | Fw | ATTACCCACCACCCACTGAC | 60 | [50] |
| Rv | TCTCTTCCACTATGCTCTCCAG | ||||
| hif-1a | DQ171936 | Fw | GACTTCAGCTGCCCTGATTC | 60 | [51] |
| Rv | GGCTGGTTTATAGCGCTGAG | ||||
| nd5 | KF857307 | Fw | CCCGATTTCTGTGCCCTACTA | 60 | [52] |
| Rv | AGGAAAGGAGTGCCTGTGA | ||||
| cyb | EF427553 | Fw | TGCCTACGCTTCCTTCGCTCGATCC | 60 | [53] |
| Rv | TAACGCCAACACCCCGCCCAAT | ||||
| cox | KF857308 | Fw | ATACTTCACATCCGCAACCATAA | 60 | [53] |
| Rv | AAGCCTCCGACTGTAAATAAGAAA | ||||
| ucp1 | MH138003 | Fw | CGATTCCAAGCCCAGACGAACCT | 60 | [53] |
| Rv | TGCCAGTGTAGCGACGAGCC | ||||
| sod | FJ860004.1 | Fw | CATGTTGGAGACCTGGGAGA | 60 | [54] |
| Rv | TGAGCATCTTGTCCGTGATGT | ||||
| cat | FJ860003.1 | Fw | TGGGACTTCTGGAGCCTGAG | 60 | [54] |
| Rv | GCAAACCTCGATCGCTGAAC | ||||
| gpx | FM013606.1 | Fw | AGTTCGTGCAGTTAATCCGGA | 60 | [54] |
| Rv | GCTTAGCTGTCAGGTCGTAAAAC | ||||
| tnf-1α | DQ200910.1 | Fw | GCCAAGCAAACAGCAGGAC | 60 | [52] |
| Rv | ACAGCGGATATGGACGGTG | ||||
| casp-9 | DQ345775 | Fw | GGCAGGACTCGACGAGATAG | 62.7 | [55] |
| Rv | CTCGCTCTGAGGAGCAAACT | ||||
| α-tub (hk) | AY326429.1 | Fw | AGGCTCATTGGCCAGATTGT | 60 | [31] |
| Rv | CAACATTCAGGGCTCCATCA | ||||
| rpl17 | AF139590 | Fw | GAGGACGTGGTGGTTCATCT | 60 | [56] |
| Rv | CTGGCTTGCCTTTCTTGACT |
| WT Genotype | GS Genotype | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | ||
| One-way ANOVA | ||||||||||
| Diet (Inside Each Genotype) | ||||||||||
| IBW (g) (300 dph) | 58 ± 9.2 | 57.8 ± 10.2 | 58.6 ± 10 | 57.5 ± 9.4 | 108.7 ± 15.4 | 106.2 ± 17.1 | 102.2 ± 17 | 102.4 ± 15.6 | ns | |
| IL (300 dph) | 17.6 ± 1 | 17.6 ± 0.9 | 17.7 ± 1 | 17.6 ± 0.87 | 20.8 ± 1.1 | 20.9 ± 1.1 | 20.5 ± 1.2 | 20.6 ± 1.3 | ns | |
| FBW (g) (372 dps) | 99.4 ± 18.3 | 95.3 ± 18.2 | 103 ± 19 | 99.8 ± 18.1 | 192.8 ± 31.7 | 189.7 ± 34 | 176.1 ± 33.2 | 180.4 ± 30.3 | ns | |
| FL (cm) (372 dph) | 21 ± 2.1 | 20.8 ± 1.2 | 21 ± 2.1 | 21.1 ± 1.2 | 25.6 ± 1.1 | 25.5 ± 1.3 | 25.1 ± 1.3 | 24.8 ± 1.4 | ns | |
| One-way ANOVA | ||||||||||
| Diet | Genotype | |||||||||
| 1 SGR (%/day) | 0.75 ± 0.03 | 0.70 ± 0.06 | 0.78 ± 0.04 | 0.76 ± 0.01 | 0.80 ± 0.01 | 0.83 ± 0.01 | 0.81 ± 0.01 | 0.80 ± 0.01 | ns | ns |
| 2 FCR | 1.84 a ± 0.16 | 1.99 a ± 0.29 | 1.78 a ± 0.17 | 1.70 a ± 0.12 | 1.48 b ± 0.01 | 1.46 b ± 0.01 | 1.58 b ± 0.01 | 1.58 b ± 0.01 | ns | F = 8.335, p-val = 0.0119 |
| 3 FI (g feed/ 100 g BW/day) | 0.48 a ± 0.02 | 0.48 a ± 0.02 | 0.47 a ± 0.02 | 0.46 a ± 0.01 | 0.27 b ± 0.00 | 0.28 b ± 0.00 | 0.27 b ± 0.00 | 0.29 b ± 0.00 | ns | F = 364.1, p-val = 2.03 × 10−11 |
| WT Genotype | GS Genotype | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | Control | PHYTO0.02 | PHYTO0.1 | GMOS0.5 | |
| Time | ||||||||
| 0 h | 3.49 a1 ± 1.10 | 3.77 a1 ± 0.62 | 3.61 a1 ± 0.92 | 3.83 a1 ± 1.15 | 0.92 b1 ± 0.23 | 1.96 b1 ± 0.89 | 1.95 b1 ± 0.21 | 1.97 b1 ± 0.85 |
| 2 h | 7.22 a2 ± 2.95 | 5.80 a2 ± 1.62 | 5.51 a2 ± 1.29 | 6.20 a2 ± 1.13 | 3.43 b2 ± 0.53 | 3.43 b2 ± 0.53 | 4.23 b2 ± 1.43 | 3.81 b2 ± 0.24 |
| 24 h | 3.26 a1 ± 0.31 | 3.54 a1 ± 1.35 | 3.57 a1 ± 1.73 | 4.10 a1 ± 1.32 | 2.14 b1 ± 0.41 | 1.61 b1 ± 0.47 | 1.58 b1 ± 0.13 | 1.79 b1 ± 0.68 |
| Three-way ANOVA | ||||||||
| Diet | Genotype | Time | D × G | D × T | G × T | D × G × T | ||
| Plasmatic cortisol | ns | F = 0.41; p-val = 2 × 10−16 | F = 55.023; p-val = 3.16 × 10−16 | ns | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serradell, A.; Montero, D.; Terova, G.; Rimoldi, S.; Makol, A.; Acosta, F.; Bajek, A.; Haffray, P.; Allal, F.; Torrecillas, S. Functional Additives in a Selected European Sea Bass (Dicentrarchus labrax) Genotype: Effects on the Stress Response and Gill Antioxidant Response to Hydrogen Peroxide (H2O2) Treatment. Animals 2023, 13, 2265. https://doi.org/10.3390/ani13142265
Serradell A, Montero D, Terova G, Rimoldi S, Makol A, Acosta F, Bajek A, Haffray P, Allal F, Torrecillas S. Functional Additives in a Selected European Sea Bass (Dicentrarchus labrax) Genotype: Effects on the Stress Response and Gill Antioxidant Response to Hydrogen Peroxide (H2O2) Treatment. Animals. 2023; 13(14):2265. https://doi.org/10.3390/ani13142265
Chicago/Turabian StyleSerradell, Antonio, Daniel Montero, Genciana Terova, Simona Rimoldi, Alex Makol, Félix Acosta, Aline Bajek, Pierrick Haffray, François Allal, and Silvia Torrecillas. 2023. "Functional Additives in a Selected European Sea Bass (Dicentrarchus labrax) Genotype: Effects on the Stress Response and Gill Antioxidant Response to Hydrogen Peroxide (H2O2) Treatment" Animals 13, no. 14: 2265. https://doi.org/10.3390/ani13142265
APA StyleSerradell, A., Montero, D., Terova, G., Rimoldi, S., Makol, A., Acosta, F., Bajek, A., Haffray, P., Allal, F., & Torrecillas, S. (2023). Functional Additives in a Selected European Sea Bass (Dicentrarchus labrax) Genotype: Effects on the Stress Response and Gill Antioxidant Response to Hydrogen Peroxide (H2O2) Treatment. Animals, 13(14), 2265. https://doi.org/10.3390/ani13142265