
Drone Observation for the Quantitative Study of Complex Multilevel Societies
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Challenges of Studying Multilevel Societies
3. Using Drones to Understand Group Structure and Measuring Social Relationships in a Multilevel Society
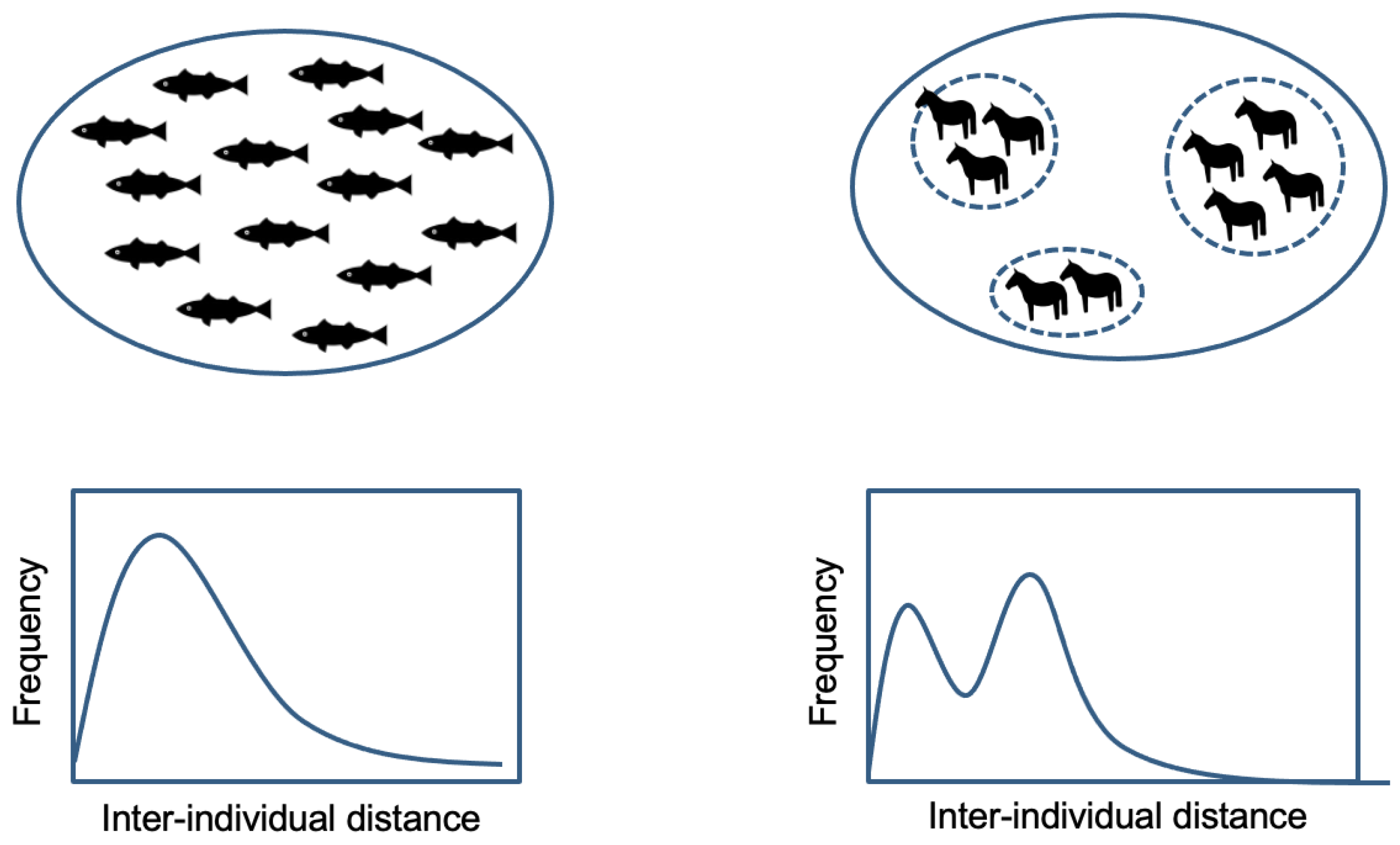
3.1. The Necessity of Quantitative Assessment of the Multilevel Social Group Structure
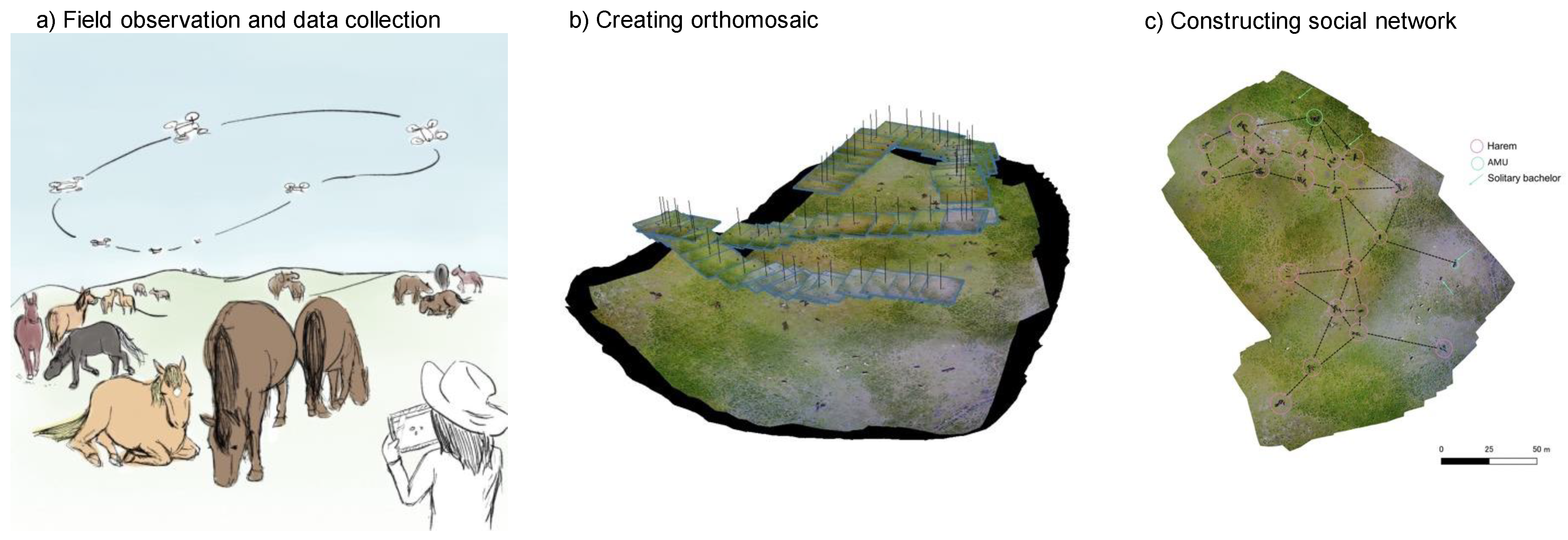
3.2. How to Apply Drone Methods
4. Collective Behavior in a Multilevel Society
4.1. Previous Studies of Collective Behavior in a Multilevel Society
4.2. How to Apply the Drone Method
5. Possibilities for Future Studies
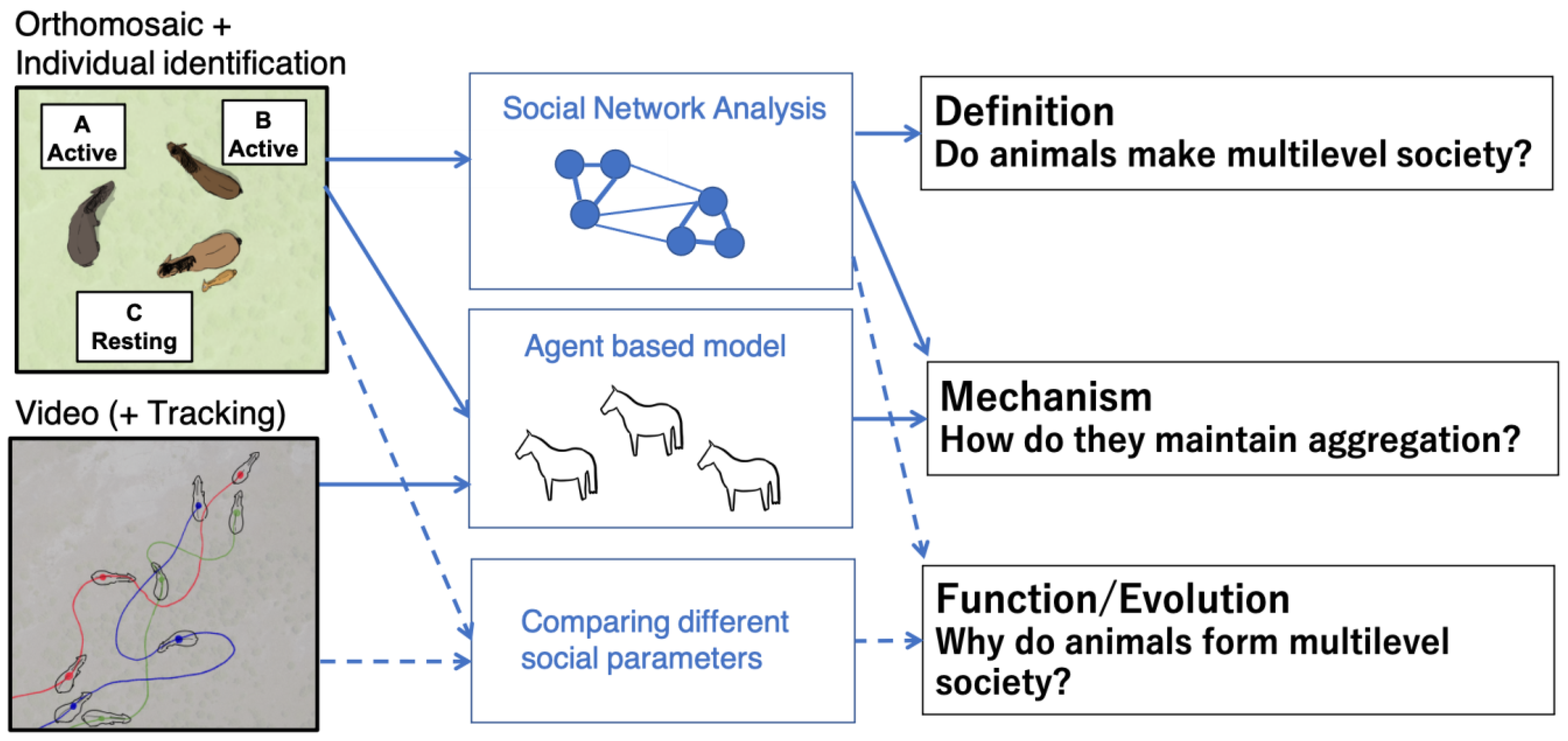
5.1. What Can Drones Contribute to Future Studies?
5.2. Challenges in Drone Observation and Their Possible Solutions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughey, L.F.; Hein, A.M.; Strandburg-Peshkin, A.; Jensen, F.H. Challenges and Solutions for Studying Collective Animal Behaviour in the Wild. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170005. [Google Scholar] [CrossRef] [PubMed]
- Kubo, D. History and Technological Developmeny of Unmanned Aircraft System (Drone). J. Soc. Instrum. Control Eng. 2017, 56, 12–56. [Google Scholar]
- Inzerillo, L.; Acuto, F.; Di Mino, G.; Uddin, M.Z. Super-Resolution Images Methodology Applied to UAV Datasets to Road Pavement Monitoring. Drones 2022, 6, 171. [Google Scholar] [CrossRef]
- Pfeifer, C.; Barbosa, A.; Mustafa, O.; Peter, H.U.; Rümmler, M.C.; Brenning, A. Using Fixed-Wing Uav for Detecting and Mapping the Distribution and Abundance of Penguins on the South Shetlands Islands, Antarctica. Drones 2019, 3, 39. [Google Scholar] [CrossRef]
- Schad, L.; Fischer, J. Opportunities and Risks in the Use of Drones for Studying Animal Behaviour. Methods Ecol. Evol. 2022, 1–9. [Google Scholar] [CrossRef]
- Inoue, S.; Yamamoto, S.; Ringhofer, M.; Mendonça, R.S.; Pereira, C.; Hirata, S. Spatial Positioning of Individuals in a Group of Feral Horses: A Case Study Using Drone Technology. Mammal Res. 2019, 64, 249–259. [Google Scholar] [CrossRef]
- Inoue, S.; Yamamoto, S.; Ringhofer, M.; Mendonça, R.S.; Hirata, S. Lateral Position Preference in Grazing Feral Horses. Ethology 2019, 126, 111–119. [Google Scholar] [CrossRef]
- Go, C.K.; Ringhofer, M.; Lao, B.; Kubo, T.; Yamamoto, S.; Ikeda, K. A Mathematical Model of Herding in Horse-Harem Group. J. Ethol. 2020, 38, 343–353. [Google Scholar] [CrossRef]
- Mendonça, R.S.; Pinto, P.; Inoue, S.; Ringhofer, M.; Godinho, R.; Hirata, S. Social Determinants of Affiliation and Cohesion in a Population of Feral Horses. Appl. Anim. Behav. Sci. 2021, 245, 105496. [Google Scholar] [CrossRef]
- Torney, C.J.; Lamont, M.; Debell, L.; Angohiatok, R.J.; Leclerc, L.-M.M.; Berdahl, A.M. Inferring the Rules of Social Interaction in Migrating Caribou. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170385. [Google Scholar] [CrossRef]
- Fudala, K.; Bialik, R.J. The Use of Drone-Based Aerial Photogrammetry in Population Monitoring of Southern Giant Petrels in ASMA 1, King George Island, Maritime Antarctica. Glob. Ecol. Conserv. 2022, 33, e01990. [Google Scholar] [CrossRef]
- Burke, C.; Rashman, M.F.; Longmore, S.N.; McAree, O.; Glover-Kapfer, P.; Ancrenaz, M.; Wich, S.A. Successful Observation of Orangutans in the Wild with Thermal-Equipped Drones. J. Unmanned Veh. Syst. 2019, 7, 235–257. [Google Scholar] [CrossRef]
- Morrison, R.E.; Eckardt, W.; Stoinski, T.S.; Brent, L.J.N. Comparing Measures of Social Complexity: Larger Mountain Gorilla Groups Do Not Have a Greater Diversity of Relationships. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201026. [Google Scholar] [CrossRef]
- Nakamura, M. What Does the Social Complexity Indicate?—Posing Questions about the Relationship between Society and Individuals. Primate Res. 2010, 26, 131–142. [Google Scholar] [CrossRef]
- Ripperger, S.P.; Carter, G.G.; Duda, N.; Koelpin, A.; Cassens, B.; Kapitza, R.; Josic, D.; Berrío-Martínez, J.; Page, R.A.; Mayer, F. Vampire Bats That Cooperate in the Lab Maintain Their Social Networks in the Wild. Curr. Biol. 2019, 29, 4139–4144. [Google Scholar] [CrossRef] [PubMed]
- Seyfarth, R.M.; Cheney, D.L. Grooming, Alliances and Reciprocal Altruism in Vervet Monkeys. Nature 1984, 308, 541–543. [Google Scholar] [CrossRef]
- Ripperger, S.P.; Carter, G.G. Social Foraging in Vampire Bats Is Predicted by Long-Term Cooperative Relationships. PLoS Biol. 2021, 19, e3001366. [Google Scholar] [CrossRef]
- Fudala, K.; Bialik, R. Breeding Colony Dynamics of Southern Elephant. Remote Sens. 2020, 12, 2964. [Google Scholar] [CrossRef]
- Grueter, C.C.; Qi, X.; Zinner, D.; Bergman, T.; Li, M.; Xiang, Z.; Zhu, P.; Migliano, A.B.; Miller, A.; Krützen, M.; et al. Multilevel Organisation of Animal Sociality. Trends Ecol. Evol. 2020, 35, 834–847. [Google Scholar] [CrossRef]
- Grueter, C.C.; Qi, X.; Li, B.; Li, M. Multilevel Societies. Curr. Biol. 2017, 27, 984–986. [Google Scholar] [CrossRef]
- Hill, K.R.; Wood, B.M.; Baggio, J.; Hurtado, A.M.; Boyd, R.T. Hunter-Gatherer Inter-Band Interaction Rates: Implications for Cumulative Culture. PLoS ONE 2014, 9, e102806. [Google Scholar] [CrossRef] [PubMed]
- Swedell, L.; Plummer, T. A Papionin Multilevel Society as a Model for Hominin Social Evolution. Int. J. Primatol. 2012, 33, 1165–1193. [Google Scholar] [CrossRef]
- Dyble, M.; Thompson, J.; Smith, D.; Salali, G.D.; Chaudhary, N.; Page, A.E.; Vinicuis, L.; Mace, R.; Migliano, A.B. Networks of Food Sharing Reveal the Functional Significance of Multilevel Sociality in Two Hunter-Gatherer Groups. Curr. Biol. 2016, 26, 2017–2021. [Google Scholar] [CrossRef] [PubMed]
- Grueter, C.C.; Matsuda, I.; Zhang, P.; Zinner, D. Multilevel Societies in Primates and Other Mammals: Introduction to the Special Issue. Int. J. Primatol. 2012, 33, 993–1001. [Google Scholar] [CrossRef]
- Snyder-Mackler, N.; Beehner, J.C.; Bergman, T.J. Defining Higher Levels in the Multilevel Societies of Geladas (Theropithecus Gelada). Int. J. Primatol. 2012, 33, 1054–1068. [Google Scholar] [CrossRef]
- Papageorgiou, D.; Christensen, C.; Gall, G.E.C.; Klarevas-Irby, J.A.; Nyaguthii, B.; Couzin, I.D.; Farine, D.R. The Multilevel Society of a Small-Brained Bird. Curr. Biol. 2019, 29, R1120–R1121. [Google Scholar] [CrossRef]
- Camerlenghi, E.; McQueen, A.; Delhey, K.; Cook, C.N.; Kingma, S.A.; Farine, D.R.; Peters, A. Cooperative Breeding and the Emergence of Multilevel Societies in Birds. Ecol. Lett. 2022, 25, 766–777. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Qi, X.; MacIntosh, A.J.J.; Watanabe, K. A Proximity-Based Social Network of a Group of Sichuan Snub-Nosed Monkeys (Rhinopithecus Roxellana). Int. J. Primatol. 2012, 33, 1081–1095. [Google Scholar] [CrossRef]
- Tong, W.; Shapiro, B.; Rubenstein, D.I. Genetic Relatedness in Two-Tiered Plains Zebra Societies Suggests That Females Choose to Associate with Kin. Behaviour 2015, 152, 2059–2078. [Google Scholar] [CrossRef]
- Bowler, M.; Knogge, C.; Heymann, E.W.; Zinner, D. Multilevel Societies in New World Primates? Flexibility May Characterize the Organization of Peruvian Red Uakaris (Cacajao Calvus Ucayalii). Int. J. Primatol. 2012, 33, 1110–1124. [Google Scholar] [CrossRef]
- Dal Pesco, F.; Trede, F.; Zinner, D.; Fischer, J. Kin Bias and Male Pair-Bond Status Shape Male-Male Relationships in a Multilevel Primate Society. Behav. Ecol. Sociobiol. 2021, 75, 24. [Google Scholar] [CrossRef]
- Forcina, G.; Vallet, D.; Le Gouar, P.J.; Bernardo-Madrid, R.; Illera, G.; Molina-Vacas, G.; Dréano, S.; Revilla, E.; Rodríguez-Teijeiro, J.D.; Ménard, N.; et al. From Groups to Communities in Western Lowland Gorillas. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182019. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, D.I.; Hack, M. Ecology and Social Structure of the Gobi Khulan Equus Hemionus Subsp. in the Gobi B National Park, Mongolia. Biol. Conserv. 2001, 1, 51–61. [Google Scholar] [CrossRef]
- Qi, X.G.; Garber, P.A.; Ji, W.; Huang, Z.P.; Huang, K.; Zhang, P.; Guo, S.T.; Wang, X.W.; He, G.; Zhang, P.; et al. Satellite Telemetry and Social Modeling Offer New Insights into the Origin of Primate Multilevel Societies. Nat. Commun. 2014, 5, 5296. [Google Scholar] [CrossRef] [PubMed]
- Fischhoff, I.R.; Sundaresan, S.R.; Cordingley, J.; Larkin, H.M.; Sellier, M.-J.; Rubenstein, D.I. Social Relationships and Reproductive State Influence Leadership Roles in Movements of Plains Zebra, Equus Burchellii. Anim. Behav. 2007, 73, 825–831. [Google Scholar] [CrossRef]
- Montanari, D.; O’Hearn, W.J.; Hambuckers, J.; Fischer, J.; Zinner, D. Coordination during Group Departures and Progressions in the Tolerant Multi-Level Society of Wild Guinea Baboons (Papio Papio). Sci. Rep. 2021, 11, 21938. [Google Scholar] [CrossRef]
- Papageorgiou, D.; Farine, D.R. Shared Decision-Making Allows Subordinates to Lead When Dominants Monopolize Resources. Sci. Adv. 2020, 6, 5881–5906. [Google Scholar] [CrossRef]
- Papageorgiou, D.; Farine, D.R. Group Size and Composition Influence Collective Movement in a Highly Social Terrestrial Bird. eLife 2020, 9, e59902. [Google Scholar] [CrossRef]
- Biquand, S.; Boug, A.; Biquand-Guyot, V.; Gauthier, J.-P. Management of Commensal Baboons in Saudi Arabia. Rev. d’Ecologie (Terre Vie) 1994, 49, 213–222. [Google Scholar] [CrossRef]
- Stead, S.M.; Teichroeb, J.A. A Multi-Level Society Comprised of One-Male and Multi-Male Core Units in an African Colobine (Colobus Angolensis Ruwenzorii). bioRxiv 2019, 14, 641746. [Google Scholar] [CrossRef]
- Chang, Z.; Yang, B.; Vigilant, L.; Liu, Z.; Ren, B.; Yang, J.; Xiang, Z.; Garber, P.A.; Li, M. Evidence of Male-Biased Dispersal in the Endangered Sichuan Snub-Nosed Monkey (Rhinopithexus Roxellana). Am. J. Primatol. 2014, 76, 72–83. [Google Scholar] [CrossRef]
- Maeda, T.; Ochi, S.; Ringhofer, M.; Sosa, S.; Sueur, C.; Hirata, S.; Yamamoto, S. Aerial Drone Observations Identified a Multilevel Society in Feral Horses. Sci. Rep. 2021, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Oda, K. Commentary: Structure from Motion (SfM) (1) The Overview of SfM and Bundle Adjustment. J. Jpn. Soc. Photogramm. 2016, 55, 206–209. [Google Scholar]
- Nakano, K. Commentary: Structure from Motion (SfM) (3)Software for Three-Dementional Shape Recovery Using SfM. J. Jpn. Soc. Photogramm. 2016, 55, 321–324. [Google Scholar] [CrossRef]
- Fuse, T. Commentary: Structure from Motion (SfM) (2) SfM and Multi-View Stereo. J. Jpn. Soc. Photogramm. 2016, 55, 259–262. [Google Scholar] [CrossRef]
- Gonçalves, J.; Henriques, R.; Alves, P.; Sousa-Silva, R.; Monteiro, A.T.; Lomba, Â.; Marcos, B.; Honrado, J. Evaluating an Unmanned Aerial Vehicle-Based Approach for Assessing Habitat Extent and Condition in Fine-Scale Early Successional Mountain Mosaics. Appl. Veg. Sci. 2016, 19, 132–146. [Google Scholar] [CrossRef]
- Han, Y.G.; Jung, S.H.; Kwon, O. How to Utilize Vegetation Survey Using Drone Image and Image Analysis Software. J. Ecol. Environ. 2017, 41, 18. [Google Scholar] [CrossRef]
- de Castro, A.I.; Shi, Y.; Maja, J.M.; Peña, J.M. Uavs for Vegetation Monitoring: Overview and Recent Scientific Contributions. Remote Sens. 2021, 13, 2139. [Google Scholar] [CrossRef]
- Vieira, G.; Mora, C.; Pina, P.; Ramalho, R.; Fernandes, R. UAV-Based Very High Resolution Point Cloud, Digital Surface Model and Orthomosaic of the Chã Das Caldeiras Lava Fields (Fogo, Cabo Verde). Earth Syst. Sci. Data 2021, 13, 3179–3201. [Google Scholar] [CrossRef]
- Opitz, R.; Herrmann, J. Recent Trends and Long-Standing Problems in Archaeological Remote Sensing. J. Comput. Appl. Archaeol. 2018, 1, 19–41. [Google Scholar] [CrossRef]
- Goebel, M.E.; Perryman, W.L.; Hinke, J.T.; Krause, D.J.; Hann, N.A.; Gardner, S.; LeRoi, D.J. A Small Unmanned Aerial System for Estimating Abundance and Size of Antarctic Predators. Polar Biol. 2015, 38, 619–630. [Google Scholar] [CrossRef]
- Maeda, T.; Sueur, C.; Hirata, S.; Yamamoto, S. Behavioural Synchronization in a Multilevel Society of Feral Horses. PLoS ONE 2021, 16, e0258944. [Google Scholar] [CrossRef] [PubMed]
- Couzin, I.D.; Krause, J. Self-Organization and Collective Behavior in Vertebrates. Adv. Study Behav. 2003, 32, 1–75. [Google Scholar] [CrossRef]
- Romenskyy, M.; Herbert-Read, J.E.; Ward, A.J.W.; Sumpter, D.J.T. Body Size Affects the Strength of Social Interactions and Spatial Organization of a Schooling Fish (Pseudomugil Signifer). R. Soc. Open Sci. 2017, 4, 161056. [Google Scholar] [CrossRef] [PubMed]
- Schellinck, J.; White, T. A Review of Attraction and Repulsion Models of Aggregation: Methods, Findings and a Discussion of Model Validation. Ecol. Model. 2011, 222, 1897–1911. [Google Scholar] [CrossRef]
- Zienkiewicz, A.K.; Ladu, F.; Barton, D.A.W.; Porfiri, M.; Bernardo, M. Di Data-Driven Modelling of Social Forces and Collective Behaviour in Zebrafish. J. Theor. Biol. 2018, 443, 39–51. [Google Scholar] [CrossRef]
- Ballerini, M.; Cabibbo, N.; Candelier, R.; Cavagna, A.; Cisbani, E.; Giardina, I.; Orlandi, A.; Parisi, G.; Procaccini, A.; Viale, M.; et al. Empirical Investigation of Starling Flocks: A Benchmark Study in Collective Animal Behaviour. Anim. Behav. 2008, 76, 201–215. [Google Scholar] [CrossRef]
- Kovalev, A.; Filippov, A.E.; Gorb, S.N. Numerical Model of the Spatio-Temporal Dynamics in a Water Strider Group. Sci. Rep. 2021, 11, 18047. [Google Scholar] [CrossRef]
- Séguret, A.; Collignon, B.; Halloy, J. Strain Differences in the Collective Behaviour of Zebrafish (Danio Rerio) in Heterogeneous Environment. R. Soc. Open Sci. 2016, 3, 160451. [Google Scholar] [CrossRef]
- Ballerini, M.; Cabibbo, N.; Candelier, R.; Cavagna, A.; Cisbani, E.; Giardina, I.; Lecomte, V.; Orlandi, A.; Parisi, G.; Procaccini, A.; et al. Interaction Ruling Animal Collective Behavior Depends on Topological Rather than Metric Distance: Evidence from a Field Study. Proc. Natl. Acad. Sci. USA 2008, 105, 1232–1237. [Google Scholar] [CrossRef]
- Couzin, I.D. Synchronization: The Key to Effective Communication in Animal Collectives. Trends Cogn. Sci. 2018, 22, 844–846. [Google Scholar] [CrossRef] [PubMed]
- Graving, J.M.; Chae, D.; Naik, H.; Li, L.; Koger, B.; Costelloe, B.R.; Couzin, I.D. Deepposekit, a Software Toolkit for Fast and Robust Animal Pose Estimation Using Deep Learning. eLife 2019, 8, e47994. [Google Scholar] [CrossRef] [PubMed]
- Ringhofer, M.; Go, C.K.; Inoue, S.; Mendonça, R.S.; Hirata, S.; Kubo, T.; Ikeda, K.; Yamamoto, S. Herding Mechanisms to Maintain the Cohesion of a Harem Group: Two Interaction Phases during Herding. J. Ethol. 2019, 38, 71–77. [Google Scholar] [CrossRef]
- Torney, C.J.; Dobson, A.P.; Borner, F.; Lloyd-Jones, D.J.; Moyer, D.; Maliti, H.T.; Mwita, M.; Fredrick, H.; Borner, M.; Hopcraft, J.G.C. Assessing Rotation-Invariant Feature Classification for Automated Wildebeest Population Counts. PLoS ONE 2016, 11, e0156342. [Google Scholar] [CrossRef] [PubMed]
- Chabot, D.; Bird, D.M. Wildlife Research and Management Methods in the 21st Century: Where Do Unmanned Aircraft Fit In? J. Unmanned Veh. Syst. 2015, 3, 137–155. [Google Scholar] [CrossRef]
- Wang, C.; Pan, R.; Wang, X.; Qi, X.; Zhao, H.; Guo, S.; Ren, Y.; Fu, W.; Zhu, Z.; Li, B. Decision-Making Process during Collective Movement Initiation in Golden Snub-Nosed Monkeys (Rhinopithecus Roxellana). Sci. Rep. 2020, 10, 480. [Google Scholar] [CrossRef] [PubMed]
- Derex, M.; Boyd, R. Partial Connectivity Increases Cultural Accumulation within Groups. Proc. Natl. Acad. Sci. USA 2016, 113, 2982–2987. [Google Scholar] [CrossRef]
- Migliano, A.B.; Battiston, F.; Viguier, S.; Page, A.E.; Dyble, M.; Schlaepfer, R.; Smith, D.; Astete, L.; Ngales, M.; Gomez-Gardenes, J.; et al. Hunter-Gatherer Multilevel Sociality Accelerates Cumulative Cultural Evolution. Sci. Adv. 2020, 6, eaax5913. [Google Scholar] [CrossRef]
- Ozogány, K.; Vicsek, T. Modeling the Emergence of Modular Leadership Hierarchy During the Collective Motion of Herds Made of Harems. J. Stat. Phys. 2014, 158, 628–646. [Google Scholar] [CrossRef]
- Grueter, C.C. Home Range Overlap as a Driver of Intelligence in Primates. Am. J. Primatol. 2015, 77, 418–424. [Google Scholar] [CrossRef]
- Ringhofer, M.; Trösch, M.; Lansade, L.; Yamamoto, S. Horses with Sustained Attention Follow the Pointing of a Human Who Knows Where Food Is Hidden. Sci. Rep. 2021, 11, 16184. [Google Scholar] [CrossRef] [PubMed]
- Ringhofer, M.; Yamamoto, S. Domestic Horses Send Signals to Humans When They Face with an Unsolvable Task. Anim. Cogn. 2017, 20, 397–405. [Google Scholar] [CrossRef]
- Proops, L.; McComb, K.; Reby, D. Cross-Modal Individual Recognition in Domestic Horses (Equus Caballus). Proc. Natl. Acad. Sci. USA 2009, 106, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Proops, L.; Mccomb, K. Cross-Modal Individual Recognition in Domestic Horses (Equus Caballus) Extends to Familiar Humans. Proc. R. Soc. B Biol. Sci. 2012, 279, 3131–3138. [Google Scholar] [CrossRef] [PubMed]
- Grueter, C.C.; Van Schaik, C.P. Evolutionary Determinants of Modular Societies in Colobines. Behav. Ecol. 2010, 21, 63–71. [Google Scholar] [CrossRef]
- Tavares, S.B.; Samarra, F.I.P.; Miller, P.J.O. A Multilevel Society of Herring-Eating Killer Whales Indicates Adaptation to Prey Characteristics. Behav. Ecol. 2017, 28, 500–514. [Google Scholar] [CrossRef]
- Witczuk, J.; Pagacz, S.; Zmarz, A.; Cypel, M. Exploring the Feasibility of Unmanned Aerial Vehicles and Thermal Imaging for Ungulate Surveys in Forests—Preliminary Results. Int. J. Remote Sens. 2018, 39, 5504–5521. [Google Scholar] [CrossRef]
- Brunton, E.A.; Leon, J.X.; Burnett, S.E. Evaluating the Efficacy and Optimal Deployment of Thermal Infrared and True-Colour Imaging When Using Drones for Monitoring Kangaroos. Drones 2020, 4, 20. [Google Scholar] [CrossRef]
- DJI Official Website. Available online: https://www.dji.com/jp (accessed on 13 November 2022).
- Sueur, C.; Petit, O.; Deneubourg, J.L. Selective Mimetism at Departure in Collective Movements of Macaca Tonkeana: An Experimental and Theoretical Approach. Anim. Behav. 2009, 78, 1087–1095. [Google Scholar] [CrossRef]
- Desai, B.; Patel, A.; Patel, V.; Shah, S.; Raval, M.S.; Ghosal, R. Identification of Free-Ranging Mugger Crocodiles by Applying Deep Learning Methods on UAV Imagery. Ecol. Inform. 2022, 72, 101874. [Google Scholar] [CrossRef]
- Andrew, W.; Greatwood, C.; Burghardt, T. Aerial Animal Biometrics: Individual Friesian Cattle Recovery and Visual Identification via an Autonomous UAV with Onboard Deep Inference. In Proceedings of the 2019 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Macau, China, 3–8 November 2019; pp. 237–243. [Google Scholar] [CrossRef]
- Landeo-Yauri, S.S.; Ramos, E.A.; Castelblanco-Martínez, D.N.; Niño-Torres, C.A.; Searle, L. Using Small Drones to Photo-Identify Antillean Manatees: A Novel Method for Monitoring an Endangered Marine Mammal in the Caribbean Sea. Endanger. Species Res. 2020, 41, 79–90. [Google Scholar] [CrossRef]
- Pomeroy, P.; O’connor, L.; Davies, P. Assessing Use of and Reaction to Unmanned Aerial Systems in Gray and Harbor Seals during Breeding and Molt in the Uk. J. Unmanned Veh. Syst. 2015, 3, 102–113. [Google Scholar] [CrossRef]
- Koski, W.R.; Gamage, G.; Davis, A.R.; Mathews, T.; Leblanc, B.; Ferguson, S.H. Evaluation of Uas for Photographic Re-Identification of Bowhead Whales, Balaena Mysticetus. J. Unmanned Veh. Syst. 2015, 3, 22–29. [Google Scholar] [CrossRef]
- Schneider, S.; Taylor, G.W.; Linquist, S.; Kremer, S.C. Past, Present and Future Approaches Using Computer Vision for Animal Re-identification from Camera Trap Data. Methods Ecol. Evol. 2019, 10, 461–470. [Google Scholar] [CrossRef]
- Vidal, M.; Wolf, N.; Rosenberg, B.; Harris, B.P.; Mathis, A. Perspectives on Individual Animal Identification from Biology and Computer Vision. Integr. Comp. Biol. 2021, 61, 900–916. [Google Scholar] [CrossRef] [PubMed]
- Valle, R.G.; Scarton, F. Drones Improve Effectiveness and Reduce Disturbance of Censusing Common Redshanks Tringa Totanus Breeding on Salt Marshes. Ardea 2020, 107, 275. [Google Scholar] [CrossRef]
- Scholten, B.D.; Beard, A.R.; Choi, H.; Baker, D.M.; Caulfield, M.E.; Proppe, D.S. Short-Term Exposure to Unmanned Aerial Vehicles Does Not Alter Stress Responses in Breeding Tree Swallows. Conserv. Physiol. 2020, 8, coaa080. [Google Scholar] [CrossRef] [PubMed]
- Ditmer, M.A.; Vincent, J.B.; Werden, L.K.; Tanner, J.C.; Laske, T.G.; Iaizzo, P.A.; Garshelis, D.L.; Fieberg, J.R. Bears Show a Physiological but Limited Behavioral Response to Unmanned Aerial Vehicles. Curr. Biol. 2015, 25, 2278–2283. [Google Scholar] [CrossRef]
- Ditmer, M.A.; Werden, L.K.; Tanner, J.C.; Vincent, J.B.; Callahan, P.; Iaizzo, P.A.; Laske, T.G.; Garshelis, D.L. Bears Habituate to the Repeated Exposure of a Novel Stimulus, Unmanned Aircraft Systems. Conserv. Physiol. 2019, 7, coy067. [Google Scholar] [CrossRef]
- Brisson-Curadeau, É.; Bird, D.; Burke, C.; Fifield, D.A.; Pace, P.; Sherley, R.B.; Elliott, K.H. Seabird Species Vary in Behavioural Response to Drone Census. Sci. Rep. 2017, 7, 17884. [Google Scholar] [CrossRef]
- Bennitt, E.; Bartlam-Brooks, H.L.A.; Hubel, T.Y.; Wilson, A.M. Terrestrial Mammalian Wildlife Responses to Unmanned Aerial Systems Approaches. Sci. Rep. 2019, 9, 2142. [Google Scholar] [CrossRef] [PubMed]
- Palomino-González, A.; Kovacs, K.M.; Lydersen, C.; Ims, R.A.; Lowther, A.D. Drones and Marine Mammals in Svalbard, Norway. Mar. Mammal Sci. 2021, 37, 1212–1229. [Google Scholar] [CrossRef]
- Weimerskirch, H.; Prudor, A.; Schull, Q. Flights of Drones over Sub-Antarctic Seabirds Show Species- and Status-Specific Behavioural and Physiological Responses. Polar Biol. 2018, 41, 259–266. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Jenni-Eiermann, S.; Strebel, N.; Sattler, T.; Negro, J.J.; Tablado, Z. Unmanned Aircraft Systems as a New Source of Disturbance for Wildlife: A Systematic Review. PLoS ONE 2017, 12, e0178448. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, T.; Yamamoto, S. Drone Observation for the Quantitative Study of Complex Multilevel Societies. Animals 2023, 13, 1911. https://doi.org/10.3390/ani13121911
Maeda T, Yamamoto S. Drone Observation for the Quantitative Study of Complex Multilevel Societies. Animals. 2023; 13(12):1911. https://doi.org/10.3390/ani13121911
Chicago/Turabian StyleMaeda, Tamao, and Shinya Yamamoto. 2023. "Drone Observation for the Quantitative Study of Complex Multilevel Societies" Animals 13, no. 12: 1911. https://doi.org/10.3390/ani13121911
APA StyleMaeda, T., & Yamamoto, S. (2023). Drone Observation for the Quantitative Study of Complex Multilevel Societies. Animals, 13(12), 1911. https://doi.org/10.3390/ani13121911