
Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
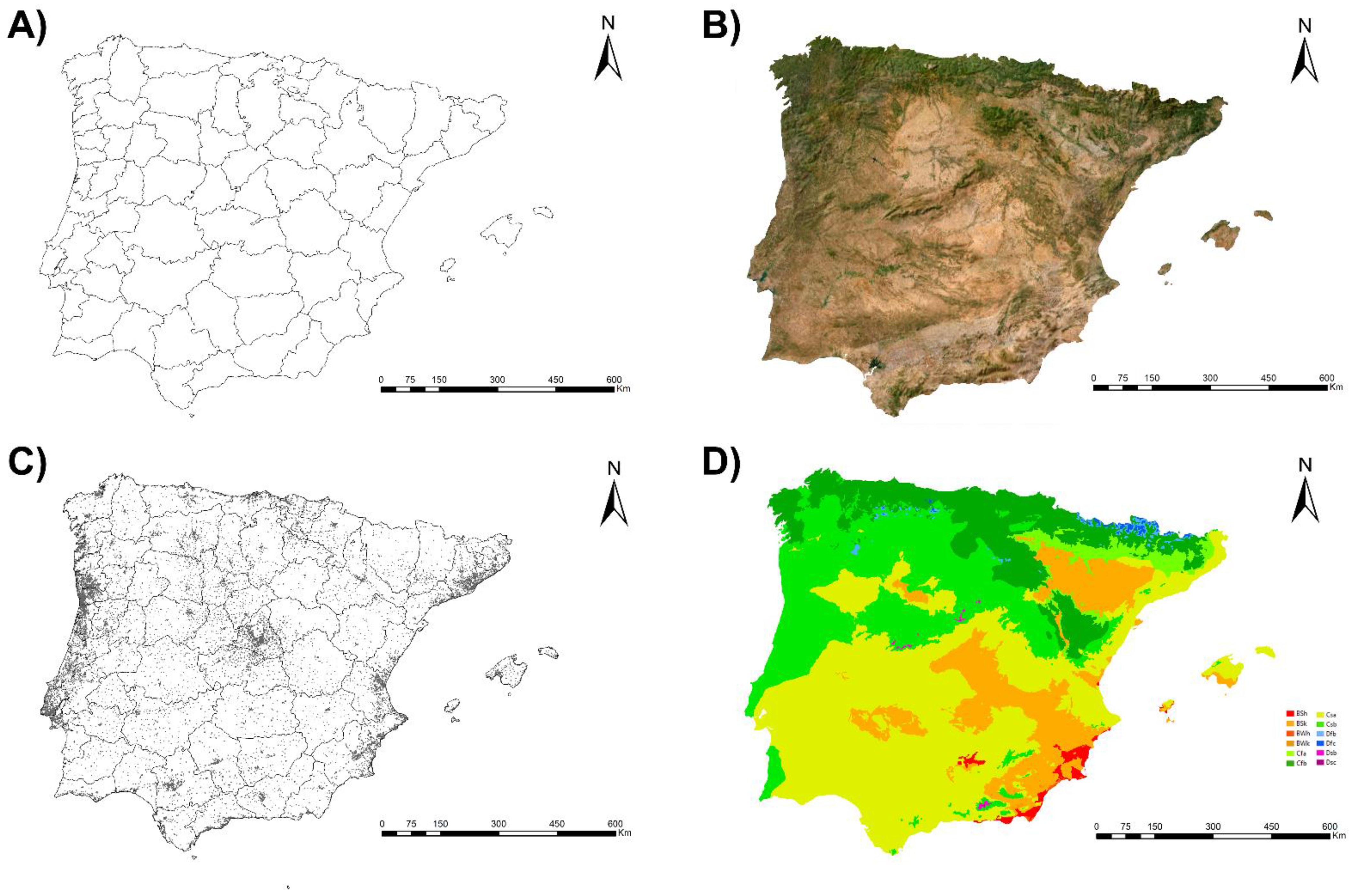
2.1. Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain)
2.2. Cx. pipiens Habitat Suitability Modelling
2.2.1. Cx. pipiens Data Collection
2.2.2. Environmental and Bioclimatic Data
2.2.3. Modeling Approaches
2.3. Dirofilaria Spp. Generations
2.4. Dirofilaria Spp. Risk Map and Its Validation
2.5. Forward Projection and Rank Change Analysis
3. Results
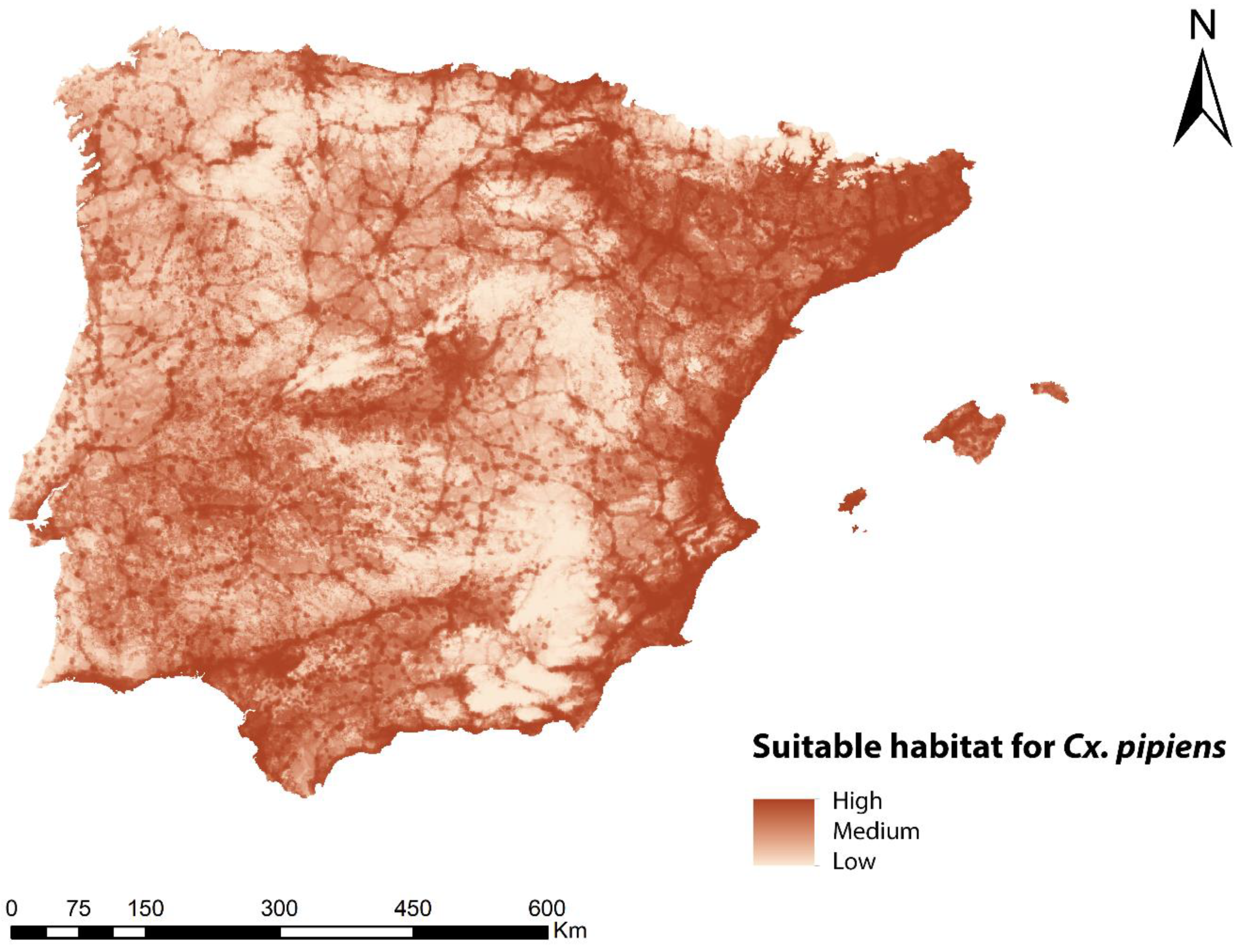
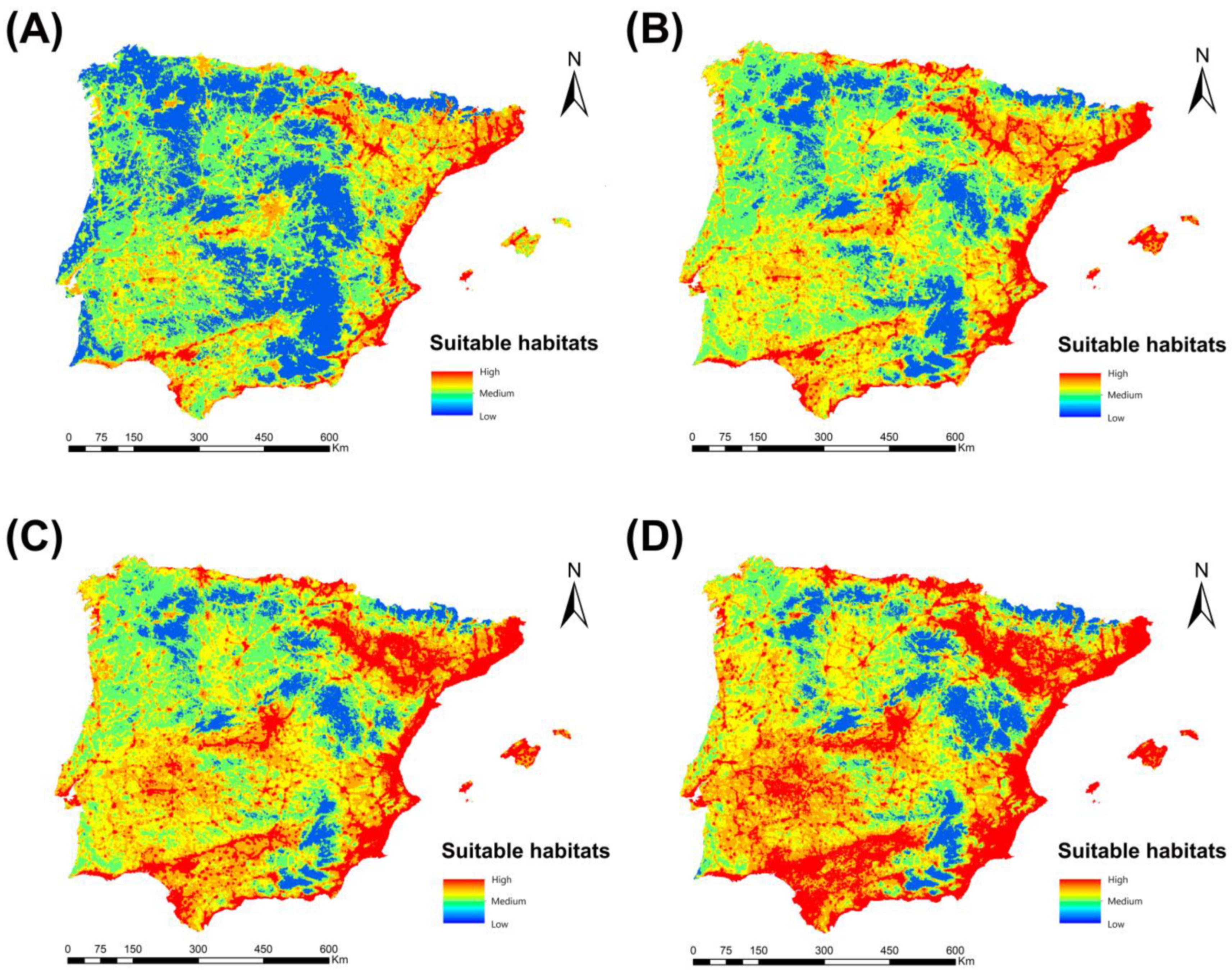
3.1. Habitat Suitability Model for Cx. pipiens
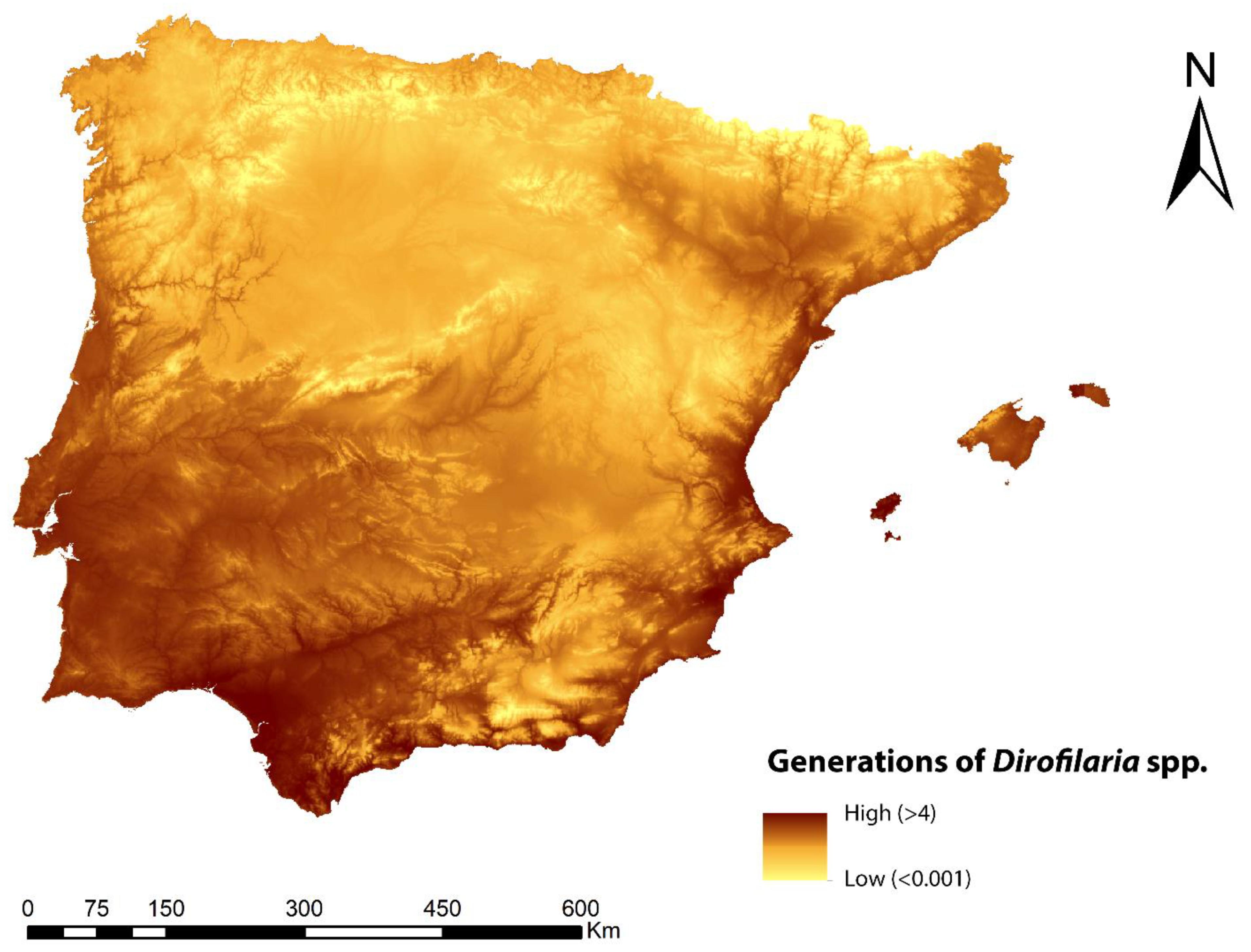
3.2. Number of Generations of Dirofilaria Spp.
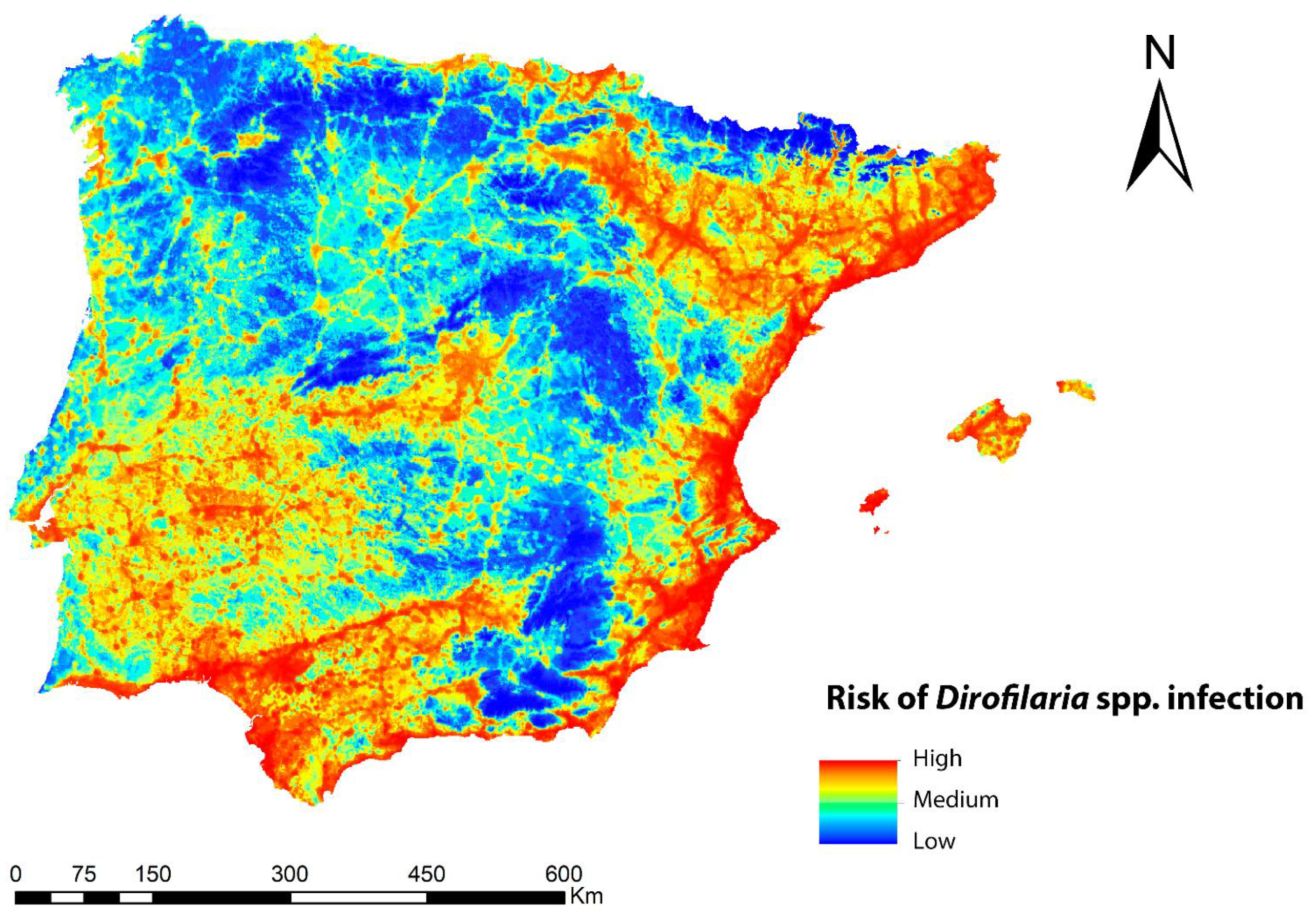
3.3. Potential Risk of Transmission of Dirofilaria spp.
3.4. Validation of the Dirofilaria spp. Transmission Risk Model
3.5. Forward Projection of Culex pipiens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Intern. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Gangoso, L.; Aragonés, D.; Martínez-de la Puente, J.; Lucientes, J.; Delacour-Estrella, S.; Estrada Peña, R.; Montalvo, T.; Bueno-Marí, R.; Bravo-Barriga, D.; Frontera, E.; et al. Determinants of the current and future distribution of the West Nile virus mosquito vector Culex pipiens in Spain. Env. Res. 2020, 188, 109837. [Google Scholar] [CrossRef] [PubMed]
- Simón, F.; Siles-Lucas, M.; Morchón, R.; González-Miguel, J.; Mellado, I.; Carretón, E.; Montoya-Alonso, J.A. Human and animal dirofilariasis: The emergence of a zoonotic mosaic. Clin. Microbiol. Rev. 2012, 25, 507–544. [Google Scholar] [CrossRef] [PubMed]
- Capelli, G.; Genchi, C.; Baneth, G.; Bourdeau, P.; Brianti, E.; Cardoso, L.; Danesi, P.; Fuehrer, H.P.; Giannelli, A.; Ionică, A.M.; et al. Recent advances on Dirofilaria repens in dogs and humans in Europe. Parasit. Vectors 2018, 1, 663. [Google Scholar] [CrossRef]
- Morchón, R.; Montoya-Alonso, J.A.; Rodríguez-Escolar, I.; Carretón, E. What Has Happened to Heartworm Disease in Europe in the Last 10 Years? Pathogens 2022, 11, 1042. [Google Scholar] [CrossRef]
- Fuehrer, H.P.; Auer, H.; Leschnik, M.; Silbermayr, K.; Duscher, G.; Joachim, A. Dirofilaria in humans, dogs, and vectors in Austria (1978–2014)—From imported pathogens to the endemicity of Dirofilaria repens. PLoS Negl. Trop. Dis. 2016, 10, e0004547. [Google Scholar] [CrossRef]
- Simón, L.; Afonin, A.; López-Díez, L.I.; González-Miguel, J.; Morchón, R.; Carretón, E.; Montoya-Alonso, J.A.; Kartashev, V.; Simón, F. Geo-environmental model for the prediction of potential transmission risk of Dirofilaria in an area with dry climate and extensive irrigated crops. The case of Spain. Vet. Parasitol. 2014, 200, 257–264. [Google Scholar] [CrossRef]
- Simón, F.; Diosdado, A.; Siles-Lucas, M.; Kartashev, V.; González-Miguel, J. Human dirofilariosis in the 21st century: A scoping review of clinical cases reported in the literature. Transbound. Emerg. Dis. 2022, 69, 2424–2439. [Google Scholar] [CrossRef]
- Cancrini, G.; Scaramozzino, P.; Gabrielli, S.; Paolo, M.D.; Toma, L.; Romi, R. Aedes albopictus and Culex pipiens implicated as natural vectors of Dirofilaria repens in central Italy. J. Med. Entomol. 2007, 44, 1064–1066. [Google Scholar] [CrossRef]
- Genchi, C.; Rinaldi, L.; Mortarino, M.; Genchi, M.; Cringoli, G. Climate and Dirofilaria infection in Europe. Vet. Parasitol. 2009, 163, 286–292. [Google Scholar] [CrossRef]
- Morchón, R.; Carretón, E.; González-Miguel, J.; Mellado-Hernández, I. Heartworm Disease (Dirofilaria immitis) and Their Vectors in Europe—New Distribution Trends. Front. Physiol. 2012, 3, 196. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Dantas-Torres, F.; Brianti, E.; Traversa, D.; Petrić, D.; Genchi, C.; Capelli, G. Vector-borne helminths of dogs and humans in Europe. Parasit. Vectors 2013, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Alonso, J.A.; Morchón, R.; García-Rodriguez, S.N.; Falcón-Cordón, Y.; Costa-Rodríguez, N.; Matos, J.I.; Escolar, I.R.; Carretón, E. Expansion of Canine Heartworm in Spain. Animals 2022, 12, 1268. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Alonso, J.A.; García Rodríguez, S.N.; Carretón, E.; Rodríguez Escolar, I.; Costa-Rodríguez, N.; Matos, J.I.; Morchón, R. Seroprevalence of Feline Heartworm in Spain: Completing the Epidemiological Puzzle of a Neglected Disease in the Cat. Front. Vet. Sci. 2022, 9, 900371. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Mendão, C.; Madeira de Carvalho, L. Prevalence of Dirofilaria immitis, Ehrlichia canis, Borrelia burgdorferi sensu lato, Anaplasma spp. and Leishmania infantum in apparently healthy and CVBD-suspect dogs in Portugal-a national serological study. Parasit. Vectors 2012, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Landum, M.; Ferreira, C.C.; Calado, M.; Alho, A.M.; Maurício, I.L.; Meireles, J.S.; de Carvalho, L.M.; Cunha, C.; Belo, S. Detection of Wolbachia in Dirofilaria infected dogs in Portugal. Vet. Parasitol. 2014, 204, 407–410. [Google Scholar] [CrossRef]
- Vieira, A.L.; Vieira, M.J.; Oliveira, J.M.; Simoes, A.R.; Diez-Banos, P.; Gestal, J. Prevalence of canine heartworm (Dirofilaria immitis) disease in dogs of central Portugal. Parasite 2014, 21, 5. [Google Scholar] [CrossRef]
- Maia, C.; Coimbra, M.; Ramos, C.; Cristóvão, J.M.; Cardoso, L.; Campino, L. Serological investigation of Leishmania infantum, Dirofilaria immitis and Angiostrongylus vasorum in dogs from southern Portugal. Parasit. Vectors 2015, 8, 152. [Google Scholar] [CrossRef]
- Alho, A.M.; Landum, M.; Ferreira, C.; Meireles, J.; Goncalves, L.; de Carvalho, L.M.; Belo, S. Prevalence and seasonal variations of canine dirofilariosis in Portugal. Vet. Parasitol. 2014, 206, 99–105. [Google Scholar] [CrossRef]
- Ferreira, C.; Afonso, A.; Calado, M.; Maurício, I.; Alho, A.M.; Meireles, J.; Madeira de Carvalho, L.; Belo, S. Molecular characterization of Dirofilaria spp. circulating in Portugal. Parasit. Vectors. 2017, 10, 250. [Google Scholar] [CrossRef]
- Perez-Sanchez, R.; Gomez-Bautista, M.; Grandes, A.E. Canine filariasis in Salamanca (northwest Spain). Ann. Trop. Med. Parasitol. 1989, 83, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Vazquez, F.A.; Valcarcel, F.; Guerrero, J.; Gómez-Bautista, M. Prevalence of canine dirofilariasis in four geographical areas of Spain. Med. Vet. 1990, 7, 297–305. [Google Scholar]
- Cancrini, G.; Allende, E.; Favia, G.; Bornay, F.; Antón, F.; Simón, F. Canine dirofilariosis in two cities of southeastern Spain. Vet. Parasitol. 2000, 92, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Lorentz, S.; Cardoso, L.; Otranto, D.; Naucke, T.J. Detection of Dirofilaria repens microfilariae in a dog from Portugal. Parasitol. Res. 2016, 115, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Baptista-Fernandes, T.; Rodrigues, M.; Domingues, D.; Monteiro, L.; Paixão, P.; Pereira, P.; Tavares, R.; Rodrigues, P.; Maurício, I.; Belo, S.; et al. Dirofilariasis by Dirofilaria repens: An imported case and a brief review. Parasitol. Int. 2015, 64, 261–263. [Google Scholar] [CrossRef]
- Rodríguez-Calzadilla, M.; Ruíz-Benítez, M.W.; de-Francisco-Ramírez, J.L.; Redondo-Campos, A.R.; Fernández-Repeto-Nuche, E.; Gárate, T.; Morchón, R. Human dirofilariasis in the eyelid caused by Dirofilaria repens: An imported case. Dirofilariasis palpebral causada por Dirofilaria repens: Un caso importado. Arch. Soc. Esp. Oftalmol. 2017, 92, 439–441. [Google Scholar] [CrossRef]
- Laynez-Roldán, P.; Martínez-de la Puente, J.; Montalvo, T.; Mas, J.; Muñoz, J.; Figuerola, J.; Rodríguez-Valero, N. Two cases of subcutaneous dirofilariasis in Barcelona, Spain. Parasitol. Res. 2018, 117, 3679–3681. [Google Scholar] [CrossRef]
- Morchón, R.; Bargues, M.D.; Latorre, J.M.; Melero-Alcíbar, R.; Pou-Barreto, C.; Mas-Coma, S.; Simón, F. Haplotype H1 of Culex pipiens implicated as natural vector of Dirofilaria immitis in an endemic area of Western Spain. Vector Borne Zoonotic Dis. 2007, 7, 653–658. [Google Scholar] [CrossRef]
- Ferreira, C.A.; de Pinho Mixão, V.; Novo, M.T.; Calado, M.M.; Gonçalves, L.A.; Belo, S.M.; de Almeida, A.P. First molecular identification of mosquito vectors of Dirofilaria immitis in continental Portugal. Parasit. Vectors 2015, 8, 139. [Google Scholar] [CrossRef]
- Bravo-Barriga, D.; Parreira, R.; Almeida, A.P.; Calado, M.; Blanco-Ciudad, J.; Serrano-Aguilera, F.J.; Pérez-Martín, J.E.; Sánchez-Peinado, J.; Pinto, J.; Reina, D.; et al. Culex pipiens as a potential vector for transmission of Dirofilaria immitis and other unclassified Filarioidea in Southwest Spain. Vet. Parasitol. 2016, 223, 173–180. [Google Scholar] [CrossRef]
- Omar, K.; Thabet, H.S.; TagEldin, R.A.; Asadu, C.C.; Chukwuekezie, O.C.; Ochu, J.C.; Dogunro, F.A.; Nwangwu, U.C.; Onwude, O.C.; Ezihe, E.K.; et al. Ecological niche modeling for predicting the potential geographical distribution of Aedes species (Diptera: Culicidae): A case study of Enugu State, Nigeria. Parasite Epidemiol. Control. 2021, 15, e00225. [Google Scholar] [CrossRef]
- Fleitas, P.E.; Kehl, S.D.; Lopez, W.; Travacio, M.; Nieves, E.; Gil, J.F.; Cimiro, R.O.; Krolewiecki, A.J. Mapping the global distribution of Strongyloides stercoralis and hookworms by ecological niche modeling. Parasit. Vectors 2022, 15, 197. [Google Scholar] [CrossRef]
- Escobar, L.E. Ecological niche modeling: An introduction for veterinarians and epidemiologists. Front. Vet. Sci. 2020, 7, 519059. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Wintle, B.A.; Elith, J.; Potts, J.M. Fauna habitat modelling and mapping: A review and case study in the Lower Hunter Central Coast region of NSW. Austral Eco. 2005, 30, 719–738. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rangel, T.F. Hutchinson’s Duality: The Once and Future Niche. Proc. Natl. Acad. Sci. USA 2009, 106, 19651–19658. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Graham, C.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.; Lehmann, A. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.T.; Soberón, J.; Overton, J.M.; Aragón, P.; Lobo, J.M. Use of niche models in invasive species risk assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- Fath, B. Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Diver. Dis. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Hernández-Lambraño, R.E.; González-Moreno, P.; Sánchez-Agudo, J.Á. Towards the Top: Niche Expansion of Taraxacum Officinale and Ulex Europaeus in Mountain Regions of South America. Austral Ecol. 2017, 42, 577–589. [Google Scholar] [CrossRef]
- Battini, N.; Farías, N.; Giachetti, C.B.; Schwindt, E.; Bortolus, A. Staying ahead of invaders: Using species distribution modeling to predict alien species’ potential niche shifts. Mar. Ecol. Prog. Ser. 2019, 612, 127–140. [Google Scholar] [CrossRef]
- Gholamrezaei, M.; Mohebali, M.; Hanafi-Bojd, A.A.; Sedaghat, M.M.; Shirzadi, M.R. Ecological Niche Modeling of main reservoir hosts of zoonotic cutaneous leishmaniasis in Iran. Acta Trop. 2016, 160, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, H.; Ali, A.; Fatima, S.H.; Zaidi, F.; Khisroon, M.; Rasheed, S.B.; Ullah, I.; Ullah, S.; Shakir, M. Spatial modeling of dengue prevalence and kriging prediction of dengue outbreak in Khyber Pakhtunkhwa (Pakistan) using presence only data. Stoch. Environ. Res. Risk Assess. 2020, 34, 1023–1036. [Google Scholar] [CrossRef]
- Boorgula, G.D.; Peterson, A.T.; Foley, D.H.; Ganta, R.R.; Raghavan, R.K. Assessing the current and future potential geographic distribution of the American dog tick, Dermacentor variabilis (Say) (Acari: Ixodidae) in North America. PLoS ONE 2020, 15, e0237191. [Google Scholar] [CrossRef]
- Cunze, S.; Kochmann, J.; Klimpel, S. Global occurrence data improve potential distribution models for Aedes japonicus japonicus in non-native regions. Pest Manag. Sci. 2020, 76, 1814–1822. [Google Scholar] [CrossRef]
- Hanafi-Bojd, A.A.; Vatandoost, H.; Yaghoobi-Ershadi, M.R. Climate change and the risk of malaria transmission in Iran. J. Med. Entomol. 2020, 57, 50–64. [Google Scholar] [CrossRef]
- Rochat, E.; Vuilleumier, S.; Aeby, S.; Greub, G.; Joost, S. Nested species distribution models of chlamydiales in Ixodes ricinus (Tick) hosts in Switzerland. Appl. Environ. Microbiol. 2020, 87, e01237-e20. [Google Scholar] [CrossRef]
- Kuhls, K.; Moskalenko, O.; Sukiasyan, A.; Manukyan, D.; Melik-Andreasyan, G.; Atshemyan, L.; Apresyan, H.; Strelkova, M.; Jaeschke, A.; Wieland, R.; et al. Microsatellite based molecular epidemiology of Leishmania infantum from re-emerging foci of visceral leishmaniasis in Armenia and pilot risk assessment by ecological niche modeling. PLoS Negl. Trop. Dis. 2021, 15, e0009288. [Google Scholar] [CrossRef]
- Charrahy, Z.; Yaghoobi-Ershadi, M.R.; Shirzadi, M.R.; Akhavan, A.A.; Rassi, Y.; Hosseini, S.Z.; Webb, N.J.; Haque, U.; Bozorg Omid, F.; Hanafi-Bojd, A.A. Climate change and its effect on the vulnerability to zoonotic cutaneous leishmaniasis in Iran. Transbound. Emerg. Dis. 2022, 69, 1506–1520. [Google Scholar] [CrossRef]
- Meshgi, B.; Majidi-Rad, M.; Hanafi-Bojd, A.A.; Fathi, S. Ecological niche modeling for predicting the habitat suitability of fascioliasis based on maximum entropy model in southern Caspian Sea littoral, Iran. Acta Trop. 2019, 198, 105079. [Google Scholar] [CrossRef]
- Di, X.; Li, S.; Ma, B.; Di, X.; Li, Y.; An, B.; Jiang, W. How climate, landscape, and economic changes increase the exposure of Echinococcus spp. BMC Public Health 2022, 22, 2315. [Google Scholar] [CrossRef] [PubMed]
- Cenni, L.; Simoncini, A.; Massetti, L.; Rizzoli, A.; Hauffe, H.C.; Massolo, A. Current and future distribution of a parasite with complex life cycle under global change scenarios: Echinococcus multilocularis in Europe. Glob. Chang. Biol. 2023, 29, 2436–2449. [Google Scholar] [CrossRef] [PubMed]
- Phang, W.K.; Hamid, M.H.A.; Jelip, J.; Mudin, R.N.; Chuang, T.W.; Lau, Y.L.; Fong, M.Y. Predicting Plasmodium knowlesi transmission risk across Peninsular Malaysia using machine learning-based ecological niche modeling approaches. Front. Microbiol. 2023, 14, 357. [Google Scholar] [CrossRef] [PubMed]
- Genchi, C.; Rinaldi, L.; Cascone, C.; Mortarino, M.; Cringoli, G. Is heartworm disease really spreading in Europe? Vet. Parasitol. 2005, 133, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Genchi, C.; Mortarino, M.; Rinaldi, L.; Cringoli, G.; Traldi, G.; Genchi, M. Changing climate and changing vector-borne disease distribution: The example of Dirofilaria in Europe. Vet. Parasitol. 2011, 176, 295–299. [Google Scholar] [CrossRef]
- Rinaldi, L.; Musella, V.; Biggeri, A.; Cringoli, G. New insights into the application of geographical information systems and remote sensing in veterinary parasitology. Geospat. Health 2006, 1, 33–47. [Google Scholar] [CrossRef]
- Medlock, J.M.; Barrass, I.; Kerrod, E.; Taylor, M.A.; Leach, S. Analysis of climatic predictions for extrinsic incubation of Dirofilaria in the United Kingdom. Vector Borne Zoonotic Dis. 2007, 7, 4–14. [Google Scholar] [CrossRef]
- Mortarino, M.; Musella, V.; Costa, V.; Genchi, C.; Cringoli, G.; Rinaldi, L. GIS modeling for canine dirofilariosis risk assessment in central Italy. Geospat. Health 2008, 2, 253–261. [Google Scholar] [CrossRef]
- Sassnau, R.; Czajka, C.; Kronefeld, M.; Werner, D.; Genchi, C.; Tannich, E.; Kampen, H. Dirofilaria repens and Dirofilaria immitis DNA findings in mosquitoes in Germany: Temperature data allow autochthonous extrinsic development. Parasitol. Res. 2014, 113, 3057–3061. [Google Scholar] [CrossRef]
- Kartashev, V.; Afonin, A.; González-Miguel, J.; Sepúlveda, R.; Simón, L.; Morchón, R.; Simón, F. Regional warming and emerging vector-borne zoonotic dirofilariosis in the Russian Federation, Ukraine, and other post-Soviet states from 1981 to 2011 and projection by 2030. BioMed Res. Intern. 2014, 2014, 858936. [Google Scholar] [CrossRef]
- Ciuca, L.; Musella, V.; Miron, L.D.; Maurelli, M.P.; Cringoli, G.; Bosco, A.; Rinaldi, L. Geographic distribution of canine heartworm (Dirofilaria immitis) infection in stray dogs of eastern Romania. Geospat. Health 2016, 11, 499. [Google Scholar] [CrossRef]
- Farkas, R.; Mag, V.; Gyurkovszky, M.; Takács, N.; Vörös, K.; Solymosi, N. The current situation of canine dirofilariosis in Hungary. Parasitol. Res. 2020, 119, 129–135. [Google Scholar] [CrossRef]
- Montoya-Alonso, J.A.; Carretón, E.; Simón, L.; González-Miguel, J.; García-Guasch, L.; Morchón, R.; Simón, F. Prevalence of Dirofilaria immitis in dogs from Barcelona: Validation of a geospatial prediction model. Vet. Parasitol. 2015, 212, 456–459. [Google Scholar] [CrossRef]
- Climate Shifts. In Worldmaps of Köppen-Geiger Climate Classification. 2019. Available online: http://koeppen-geiger.vuwien.ac.at/shifts.html (accessed on 12 April 2023).
- Global Biodiversity Information Facility. 2016. Available online: www.gbif.org (accessed on 23 March 2023).
- World Clim. 2023. Available online: www.worldclim.org (accessed on 11 February 2023).
- Ripley, B.D. The R project in statistical computing. MSOR Connections. The Newsletter of the LTSN Maths, Stats OR Network 1.1. MSOR Connect. 2001, 1, 23–25. [Google Scholar] [CrossRef]
- Socioeconomic Data and Applications Center. 2023. Available online: https://sedac.ciesin.columbia.edu (accessed on 22 May 2023).
- Corine Land Cover. Copernicus Global Land Service. Available online: https://land.copernicus.eu/pan-european/corine-land-cover/clc2018 (accessed on 12 April 2023).
- EarthEnv. 2023. Available online: http://www.earthenv.org/landcover (accessed on 12 April 2023).
- American Museum of Natural History. 2023. Available online: https://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 22 May 2023).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. Kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed]
- Climatologies at High Resolution for the Earth’s Land Surface Areas. 2023. Available online: https://chelsa-climate.org/ (accessed on 23 March 2023).
- Karger, D.N.; Lange, S.; Hari, C.; Reyer, C.P.; Zimmermann, N.E. CHELSA-W5E5 v1. 0: W5E5 v1. 0 Downscaled with CHELSA v2. 0.; ISIMIP: Potsdam, Germany, 2021. [Google Scholar]
- Alho, A.M.; Meireles, J.; Schnyder, M.; Cardoso, L.; Belo, S.; Deplazes, P.; de Carvalho, L.M. Dirofilaria immitis and Angiostrongylus vasorum: The current situation of two major canine heartworms in Portugal. Vet. Parasitol. 2018, 252, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Kuhl Brodt, T.; Jones, C.G.; Sellar, A.; Storkey, D.; Blockley, E.; Stringer, M.; Hill, R.; Graham, T.; Ridley, J.; Blaker, A.; et al. The low-resolution version of HadGEM3 GC3.1: Development and evaluation for global climate. J. Adv. Model. Earth Syst. 2018, 10, 2865–2888. [Google Scholar] [CrossRef]
- Andrews, M.B.; Ridley, J.K.; Wood, R.A.; Andrews, T.; Blockley, E.W.; Booth, B.; Burke, E.; Dittus, A.J.; Florek, P.; Gray, L.J.; et al. Historical simulations with HadGEM3-GC3.1 for CMIP6. J. Adv. Model. Earth Syst. 2020, 12, e2019MS001995. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; y Araújo, M.B. BIOMOD: Una plataforma para la predicción por conjuntos de la distribución de especies. Ecografía 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Guerrero, J.; Ducos de la Hitte, J.; Genchi, C.; Rojo, F.; Gomez-Bautista, M.; Carvalho Valera, M.; Labarthe, N.; Bordini, E.; Gonzales, G.; Mancebo, O.; et al. Update on the distribution of Dirofilaria immitis in dogs from Southern Europe and Latin America. In Proceedings of the American Heartworm Symposium, Austin, TX, USA, 27–29 March 1992; pp. 31–37. [Google Scholar]
- Morchón, R.; Moya, I.; González-Miguel, J.; Montoya, M.N.; Simón, F. Zoonotic Dirofilaria immitis infections in a province of Northern Spain. Epidemiol. Infect. 2010, 138, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Alonso, J.A.; Morchón, R.; Falcón-Cordón, Y.; Falcón-Cordón, S.; Simón, F.; Carretón, E. Prevalence of heartworm in dogs and cats of Madrid, Spain. Parasit. Vectors 2017, 10, 354. [Google Scholar] [CrossRef]
- Diosdado, A.; Gómez, P.J.; González-Miguel, J.; Simón, F.; Morchón, R. Current status of canine dirofilariosis in an endemic area of western Spain. J. Helminthol. 2018, 92, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Pérez Pérez, P.; Rodríguez-Escolar, I.; Carretón, E.; Sánchez Agudo, J.Á.; Lorenzo-Morales, J.; Montoya-Alonso, J.A.; Morchón, R. Serological Survey of Canine Vector-Borne Infections in North-Center Spain. Front. Vet. Sci. 2021, 8, 784331. [Google Scholar] [CrossRef] [PubMed]
- Hongoh, V.; Berrang-Ford, L.; Scott, M.E.; Lindsay, L.R. Expanding geographical distribution of the mosquito, Culex pipiens, in Canada under climate change. Appl. Geogr. 2012, 33, 53–62. [Google Scholar] [CrossRef]
- Githeko, A.K.; Lindsay, S.W.; Confalonieri, U.E.; Patz, J.A. Climate change and vector-borne diseases: A regional analysis. Bull. World Health Organ. 2000, 78, 1136–1147. [Google Scholar]
- Semenza, J.C.; Menne, B. Climate change and infectious diseases in Europe. Lancet Infect. Dis. 2009, 9, 365–375. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Percent Contribution |
|---|---|
| Human footprint | 56.7% |
| BIO8 (mean temperature of the wettest quarter) | 27.2% |
| BIO4 (temperature seasonality) | 3.6% |
| Rivers | 3.3% |
| Herbaceous density | 2.1% |
| BIO9 (mean temperature of the driest quarter) | 1.9% |
| BIO15 (precipitation seasonality) | 1.5% |
| BIO1 (annual mean temperature) | 1.2% |
| Shrub density | 0.7% |
| Water bodies | 0.6% |
| Irrigated crops | 0.6% |
| BIO3 (isothermality) | 0.4% |
| BIO12 (annual precipitation) | 0.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Escolar, I.; Hernández-Lambraño, R.E.; Sánchez-Agudo, J.Á.; Collado, M.; Pérez-Pérez, P.; Morchón, R. Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios. Animals 2023, 13, 1764. https://doi.org/10.3390/ani13111764
Rodríguez-Escolar I, Hernández-Lambraño RE, Sánchez-Agudo JÁ, Collado M, Pérez-Pérez P, Morchón R. Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios. Animals. 2023; 13(11):1764. https://doi.org/10.3390/ani13111764
Chicago/Turabian StyleRodríguez-Escolar, Iván, Ricardo E. Hernández-Lambraño, José Ángel Sánchez-Agudo, Manuel Collado, Patricia Pérez-Pérez, and Rodrigo Morchón. 2023. "Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios" Animals 13, no. 11: 1764. https://doi.org/10.3390/ani13111764
APA StyleRodríguez-Escolar, I., Hernández-Lambraño, R. E., Sánchez-Agudo, J. Á., Collado, M., Pérez-Pérez, P., & Morchón, R. (2023). Current Risk of Dirofilariosis Transmission in the Iberian Peninsula (Spain and Portugal) and the Balearic Islands (Spain) and Its Future Projection under Climate Change Scenarios. Animals, 13(11), 1764. https://doi.org/10.3390/ani13111764