
Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Design of Experiment and Housing
2.2. Chlorella Vulgaris Biomass
2.3. Diets and Feeding
2.4. Broiler Performance
2.5. Immune Responses
2.6. Intestinal Histomorphology
2.7. Statistical Analysis
3. Results
3.1. Production Performances
3.2. Immune Responses
3.3. Histomorphology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Świątkiewicz, S.; Arczewska-Włosek, A.; Józefiak, D. Application of microalgae biomass in poultry nutrition. World’s Poult. Sci. J. 2015, 71, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Arif, M.; Khafaga, A.F.; Taha, A.E. The application of the microalgae Chlorella spp. As a supplement in broiler feed. World’s Poult. Sci. J. 2019, 75, 305–318. [Google Scholar] [CrossRef]
- Coudert, E.; Baéza, E.; Berri, C. Use of algae in poultry production: A review. World’s Poult. Sci. J. 2020, 76, 767–786. [Google Scholar] [CrossRef]
- An, B.-K.; Kim, K.-E.; Jeon, J.-Y.; Lee, K.W. Effect of dried Chlorella vulgaris and Chlorella growth factor on growth performance, meat qualities and humoral immune responses in broiler chickens. SpringerPlus 2016, 5, 718. [Google Scholar] [CrossRef] [Green Version]
- Andrade, L.M. Chlorella and spirulina microalgae as sources of functional foods, nutraceuticals, and food supplements; an overview. MOJ Food Process. Technol. 2018, 6, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Pignolet, O.; Jubeau, S.; Vaca-Garcia, C.; Michaud, P. Highly valuable microalgae: Biochemical and topological aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Lipstein, B.; Hurwitz, S. The Nutritional Value of Sewage-Grown Samples of Chlorella and Micractinium in Broiler Diets. Poult. Sci. 1983, 62, 1254–1260. [Google Scholar] [CrossRef]
- Enalgae. Available online: https://edepot.wur.nl/347712 (accessed on 20 February 2022).
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Kotrbáček, V.; Doubek, J.; Doucha, J. The chlorococcalean alga Chlorella in animal nutrition: A review. J. Appl. Phycol. 2015, 27, 2173–2180. [Google Scholar] [CrossRef]
- Bastiaens, L.; Van Roy, S.; Thomassen, G.; Elst, K. Biorefinery of Algae: Technical and Economic Considerations; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 9780081010273. [Google Scholar]
- Kar, S.K. FeedOmics, an Approach to Evaluate the Functional Properties of Protein Containing Feed Ingredients; Wageningen University: Wageningen, The Netherlands, 2017; p. 4442154. [Google Scholar]
- Yan, L.; Lim, S.U.; Kim, I.H. Effect of fermented chlorella supplementation on growth performance, nutrient digestibility, blood characteristics, fecal microbial and fecal noxious gas content in growing pigs. Asian-Australas. J. Anim. Sci. 2012, 25, 1742–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [Green Version]
- Mirzaie, S.; Sharifi, S.D.; Zirak-Khattab, F. The effect of a Chlorella by-product dietary supplement on immune response, antioxidant status, and intestinal mucosal morphology of broiler chickens. J. Appl. Phycol. 2020, 32, 1771–1777. [Google Scholar] [CrossRef]
- Sugiharto, S. Chlorella vulgaris and Spirulina platensis: Their Nutrient Contents and Bioactive Compounds for Improving Poultry Productivity. War. Indones. Bull. Anim. Vet. Sci. 2020, 30, 123–138. [Google Scholar] [CrossRef]
- Alfaia, C.M.; Pestana, J.M.; Rodrigues, M.; Coelho, D.; Aires, M.J.; Ribeiro, D.M.; Major, V.T.; Martins, C.F.; Santos, H.; Lopes, P.A.; et al. Influence of dietary Chlorella vulgaris and carbohydrate-active enzymes on growth performance, meat quality and lipid composition of broiler chickens. Poult. Sci. 2021, 100, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Salim, H.M.; Akter, N.; Kim, D.W.; Kim, J.H.; Bang, H.T.; Kim, M.J.; Na, J.C.; Hwangbo, J.; Choi, H.C.; et al. Effect of various forms of dietary Chlorella supplementation on growth performance, immune characteristics, and intestinal microflora population of broiler chickens. J. Appl. Poult. Res. 2013, 22, 100–108. [Google Scholar] [CrossRef]
- Zheng, L.; Oh, S.T.; Jeon, J.Y.; Moon, B.H.; Kwon, H.S.; Lim, S.U.; An, B.K.; Kang, C.W. The Dietary Effects of Fermented Chlorella vulgaris (CBT®) on Production Performance, Liver Lipids and Intestinal Microflora in Laying Hens. Asian-Australas. J. Anim. Sci. 2011, 25, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Mirzaie, S.; Tabarsa, M.; Safavi, M. Effects of extracted polysaccharides from a Chlorella vulgaris biomass on expression of interferon-γ and interleukin-2 in chicken peripheral blood mononuclear cells. J. Appl. Phycol. 2020, 33, 409–418. [Google Scholar] [CrossRef]
- CVB. Tabellenboek Veevoeding 2018. Voedernormen Pluimvee en Voederwaarden Voedermiddelen Voor Pluimvee [Livestock Feed Table 2018. Tables Values Poultry Nutrition and Feed Values Raw Materials for Poultry]. 2018. Available online: https://www.cvbdiervoeding.nl/pagina/10081/downloads.aspx (accessed on 20 February 2022).
- Mesa, D.; Muniz, E.; Souza, A.; Geffroy, B. Broiler-Housing Conditions Affect the Performance. Braz. J. Poult. Sci. 2017, 19, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, I.; Choct, M. Feed intake pattern of broiler chickens under intermittent lighting: Do birds eat in the dark? Anim. Nutr. 2019, 5, 174–178. [Google Scholar] [CrossRef]
- Szőllősi, L.; Béres, E.; Szűcs, I. Effects of modern technology on broiler chicken performance and economic indicators—A Hungarian case study. Ital. J. Anim. Sci. 2021, 20, 188–194. [Google Scholar] [CrossRef]
- Le Floch, J.-P.; Escuyer, P.; Baudin, E.; Baudon, D.; Perlemuter, L. Blood Glucose Area Under the Curve: Methodological Aspects. Diabetes Care 1990, 13, 172–175. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 20 February 2022).
- Choi, H.; Jung, S.K.; Kim, J.S.; Kim, K.W.; Oh, K.B.; Lee, P.Y.; Byun, S.J. Effects of dietary recombinant chlorella supplementation on growth performance, meat quality, blood characteristics, excreta microflora, and nutrient digestibility in broilers. Poult. Sci. 2017, 96, 710–716. [Google Scholar] [CrossRef] [PubMed]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of Dietary Microalgae on Growth Performance, Profiles of Amino and Fatty Acids, Antioxidant Status, and Meat Quality of Broiler Chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, M.; Zaghari, M.; Moravej, H. A survey on Chlorella vulgaris effect’s on performance and cellular immunity in broilers. Int. J. Agric. Sci. Res. 2012, 3, 9–15. [Google Scholar]
- An, H.; Rim, H.; Jeong, H.; Hong, S.; Um, J.; Kim, H. Hot water extracts of Chlorella vulgaris improve immune function in protein-deficient weanling mice and immune cells. Immunopharmacol. Immunotoxicol. 2010, 32, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Hulst, M.; Van der Weide, R.; Hoekman, A.; Van Krimpen, M. Transcriptional response of cultured porcine intestinal epithelial cells to micro algae extracts in the presence and absence of enterotoxigenic Escherichia coli. Genes Nutr. 2019, 14, 1–15. [Google Scholar] [CrossRef]
- Ao, Z.; Kocher, A.; Choct, M. Effects of Dietary Additives and Early Feeding on Performance, Gut Development and Immune Status of Broiler Chickens Challenged with Clostridium perfringens. Asian-Australas. J. Anim. Sci. 2012, 25, 541. [Google Scholar] [CrossRef] [Green Version]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721.e70. [Google Scholar]
- Asasi, K.; Mohammadi, A.; Boroomand, Z.; Hosseinian, S.A.; Nazifi, S. Changes of several acute phase factors in broiler chickens in response to infectious bronchitis virus infection. Poult. Sci. 2013, 92, 1989. [Google Scholar] [CrossRef]
- Kuwahara, N.; Yamaguchi, M.; Tanaka, A.; Ohta, S.; Uno, T.; Uchida, Y.; Manabe, R.; Jinno, M.; Hirai, K.; Miyata, Y.; et al. Inhibitory Effects of Chlorella Extract on Airway Hyperresponsiveness and Airway Remodeling in a Murine Model of Asthma. Showa Univ. J. Med. Sci. 2019, 31, 1–12. [Google Scholar] [CrossRef]
- Ewart, H.S.; Bloch, O.; Girouard, G.S.; Kralovec, J.; Barrow, C.J.; Ben-Yehudah, G.; Suárez, E.R.; Rapoport, M.J. Stimulation of Cytokine Production in Human Peripheral Blood Mononuclear Cells by an Aqueous Chlorella Extract. Planta Med. 2007, 73, 762–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.I. Astaxanthin as a peroxisome proliferator-activated receptor (PPAR) modulator: Its therapeutic implications. Mar. Drugs 2019, 17, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Ciliberti, M.G.; Albenzio, M.; Francavilla, M.; Neglia, G.; Esposito, L.; Caroprese, M. Extracts from microalga chlorella sorokiniana exert an anti-proliferative effect and modulate cytokines in sheep peripheral blood mononuclear cells. Animals 2019, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, Y.-C.; Prakash, E.; Huang, C.-F.; Lien, T.-W.; Chen, X.; Su, I.-J.; Chao, Y.-S.; Hsieh, H.-P.; Hsu, J.T.-A. Bioassay-guided purification and identification of PPARα/γ agonists fromChlorella sorokiniana. Phyther. Res. 2008, 22, 605–613. [Google Scholar] [CrossRef]
- Li, Y.; Lammi, C.; Boschin, G.; Arnoldi, A.; Aiello, G. Recent Advances in Microalgae Peptides: Cardiovascular Health Benefits and Analysis. J. Agric. Food Chem. 2019, 67, 11825–11838. [Google Scholar] [CrossRef]
- Buzala, M.; Janicki, B. Review: Effects of different growth rates in broiler breeder and layer hens on some productive traits. Poult. Sci. 2016, 95, 2151–2159. [Google Scholar] [CrossRef]
- Yu, K.; Choi, I.; Yun, C.-H. Immunosecurity: Immunomodulants enhance immune responses in chickens. Anim. Biosci. 2021, 34, 321–337. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Klasing, K.C. Nutrition and the immune system. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Park, S.B.; Kim, C.H. Effects of dietary supplementation with a chlorella by-product on the growth performance, immune response, intestinal microflora and intestinal mucosal morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101, 208–214. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Measured Parameters | CON | CV | p-Value 2 | |
|---|---|---|---|---|
| Performance parameters 3 of broilers during day 0–14 | ||||
| Growth performance | BW start (g) on day 0 | 46.9 ± 1.0 | 47.1 ± 0.6 | 0.70 |
| BW end (g) on day 14 | 481.4 ± 34.6 | 482.9 ± 39.2 | 0.95 | |
| dBWG (g/d) | 31.0 ± 2.5 | 31.1 ± 2.8 | 0.96 | |
| FCR (g/g) | 1.21 ± 0.01 | 1.21 ± 0.02 | 0.76 | |
| dFI (g/d) | 41.6 ± 3.2 | 41.8 ± 3.8 | 0.91 | |
| Performance parameters 3 of broilers during day 14–28 | ||||
| BW start (g) on day 14 | 481.4 ± 34.6 | 482.9 ± 39.2 | 0.95 | |
| BW end (g) on day 28 | 1738.6 ± 85.4 | 1787.3 ± 73.9 | 0.36 | |
| dBWG (g/d) | 90.1 ± 5.1 | 93.3 ± 3.3 | 0.27 | |
| FCR (g/g) | 1.38 ± 0.02 | 1.36 ± 0.02 | 0.30 | |
| dFI (g/d) | 123.9 ± 6.2 | 127.2 ± 4.4 | 0.35 | |
| Performance parameters 3 of broilers during day 28–35 | ||||
| BW start (g) on day 28 | 1738.6 ± 85.4 | 1787.3 ± 73.9 | 0.36 | |
| BW end (g) on day 35 | 2509.5 ± 75.9 | 2606.3 ± 63.2 | 0.05 | |
| dBWG (g/d) | 110.1 ± 7.9 | 117.0 ± 6.2 | 0.16 | |
| FCR (g/g) | 1.69 ± 0.10 | 1.65 ± 0.08 | 0.52 | |
| dFI (g/d) | 185.2 ± 8.1 | 192.3 ± 5.3 | 0.13 | |
| Performance parameters 3 of broilers during the whole experimental period (day 0–35) | ||||
| BW start (g) on day 0 | 46.9 ± 1.0 | 47.1 ± 0.6 | 0.70 | |
| BW end (g) on day 35 | 2509.5 ± 75.9 | 2606.3± 63.2 | 0.05 | |
| dBWG (g/d) | 70.4 ± 2.2 | 73.1 ± 1.8 | 0.05 | |
| FCR (g/g) | 1.47 ± 0.02 | 1.45 ± 0.02 | 0.23 | |
| dFI (g/d) | 103.2 ± 4.0 | 106.1 ± 3.4 | 0.25 | |
| FCRcorrected 4 (g/g) | 1.39 ± 0.02 | 1.42 ± 0.02 | 0.02 | |
| Area under the curve (AUC) of haptoglobin and interleukin-13 (IL-13) 5 during grower phase | ||||
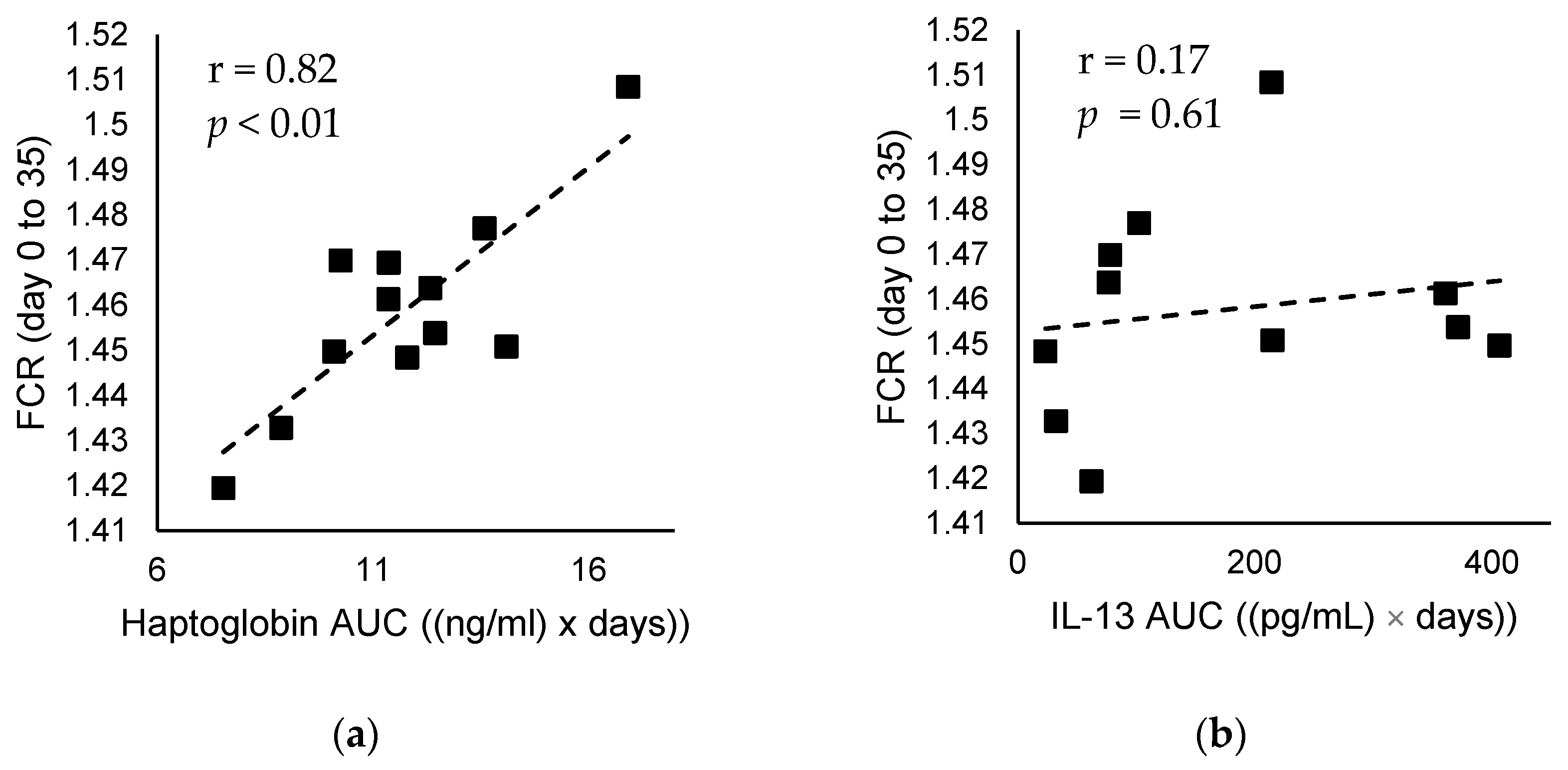
| Immune response | Haptoglobin AUC ((ng/mL) × days) | 13.1 ± 2.8 | 10.4 ± 2.5 | 0.02 |
| IL-13 AUC ((pg/mL) × days) 6 | 8.4 ± 2.3 | 5.1 ± 1.9 | <0.01 | |
| Haptoglobin and IL-13 concentrations 7 on day 14 | ||||
| Haptoglobin (ng/mL) | 2.3 ± 0.8 | 1.8 ± 0.6 | 0.09 8 | |
| IL-13 (pg/mL) | 61.8 ± 63.7 | 8.5 ± 5.2 | <0.01 8 | |
| Haptoglobin and IL-13 concentrations on day 21 | ||||
| Haptoglobin (ng/mL) | 1.4 ± 0.3 | 1.2 ± 0.3 | 0.06 | |
| IL-13 (pg/mL) | 18.2 ± 16.8 | 18.6 ± 19.0 | 0.41 8 | |
| Jejunal morphology 7 on day 14 | ||||
| Intestinal morphology | Villi length (μm) | 679.0 ± 70.5 | 642.4 ± 97.5 | 0.32 |
| Crypt depth (μm) | 174.7 ± 17.7 | 186.3 ± 28.4 | 0.27 | |
| V:C 9 ratio | 4.0 ± 0.4 | 3.6 ± 0.7 | 0.15 | |
| Villi size area (μm2) | 95,878 ± 14,203 | 88,178 ± 13,977 | 0.21 | |
| Goblet cell surface area (% of villi) | 2.0 ± 0.6 | 2.1 ± 0.8 | 0.85 | |
| Goblet cells per villi | 197.3 ± 78.1 | 178.3 ± 66.5 | 0.54 | |
| Goblet cell size (μm2) | 10.6 ± 2.6 | 10.1 ± 1.1 | 0.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roques, S.; Koopmans, S.-J.; Mens, A.; van Harn, J.; van Krimpen, M.; Kar, S.K. Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens. Animals 2022, 12, 1114. https://doi.org/10.3390/ani12091114
Roques S, Koopmans S-J, Mens A, van Harn J, van Krimpen M, Kar SK. Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens. Animals. 2022; 12(9):1114. https://doi.org/10.3390/ani12091114
Chicago/Turabian StyleRoques, Simon, Sietse-Jan Koopmans, Annemarie Mens, Jan van Harn, Marinus van Krimpen, and Soumya Kanti Kar. 2022. "Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens" Animals 12, no. 9: 1114. https://doi.org/10.3390/ani12091114
APA StyleRoques, S., Koopmans, S.-J., Mens, A., van Harn, J., van Krimpen, M., & Kar, S. K. (2022). Effect of Feeding 0.8% Dried Powdered Chlorella vulgaris Biomass on Growth Performance, Immune Response, and Intestinal Morphology during Grower Phase in Broiler Chickens. Animals, 12(9), 1114. https://doi.org/10.3390/ani12091114