Using the Ratio of Urine Testosterone to Estrone-3-Glucuronide to Identify the Sex of Chinese Giant Salamanders (Andrias davidianus)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Husbandry, Management, and Sexing of the Animals
2.3. Experimental Reagents and Consumables
2.4. Experimental Apparatus
2.5. Experimental Methods
2.5.1. Urine Collection
2.5.2. Steps for Determining the Concentration of T and E1G in Urine
2.6. Statistical Analysis
3. Results
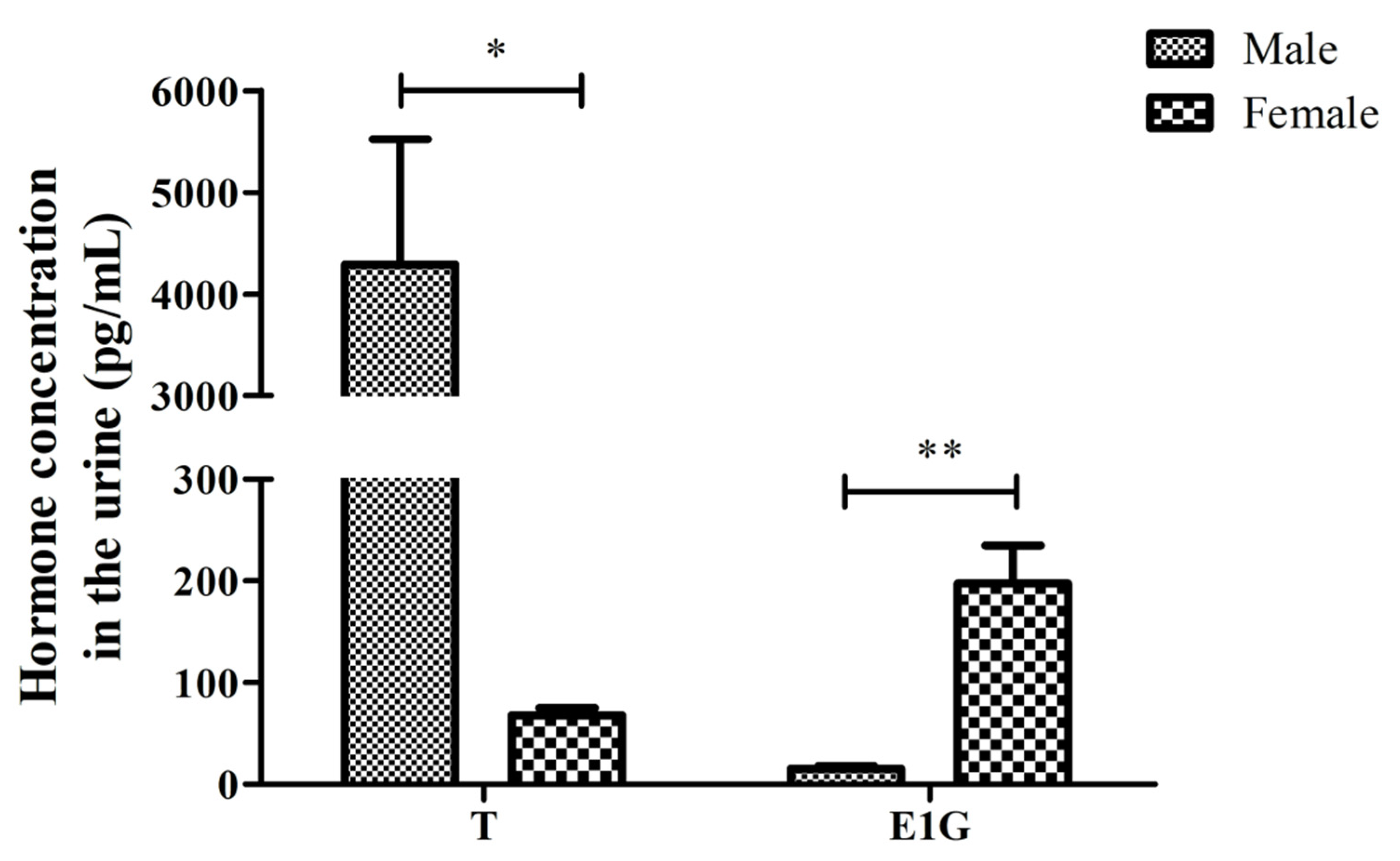
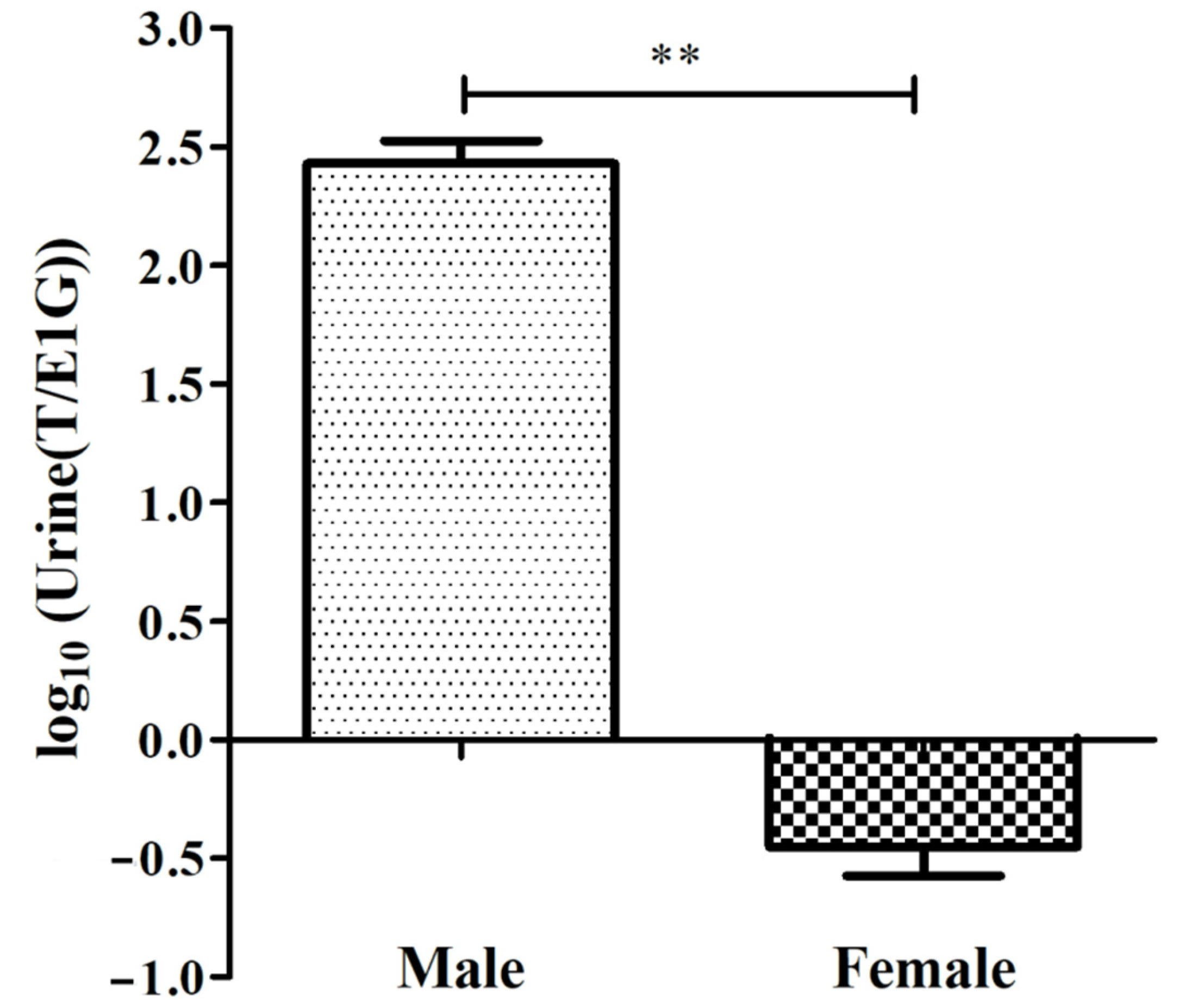
3.1. Determining the Concentration of T and E1G in the Urine of CGSs during the Breeding Season
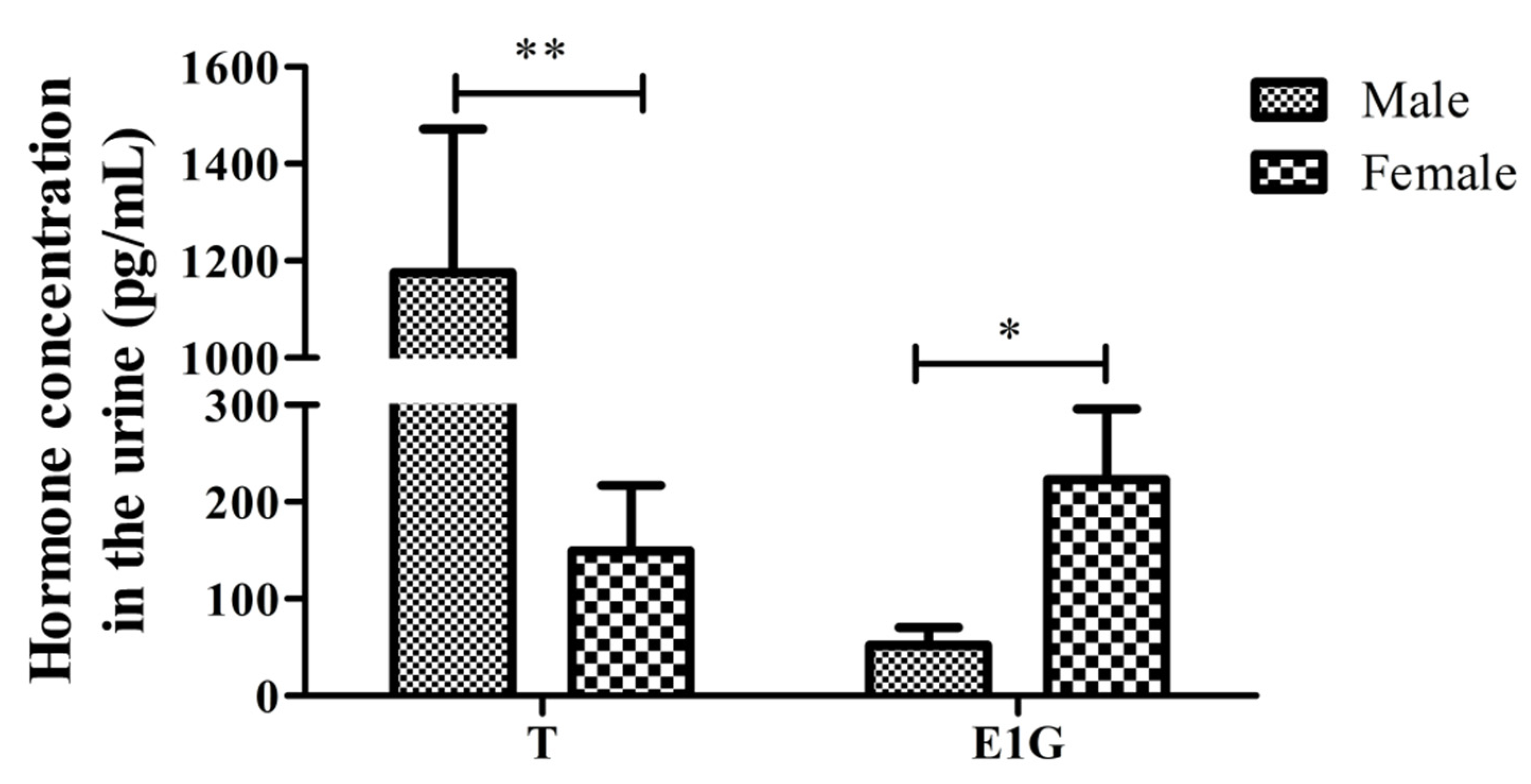
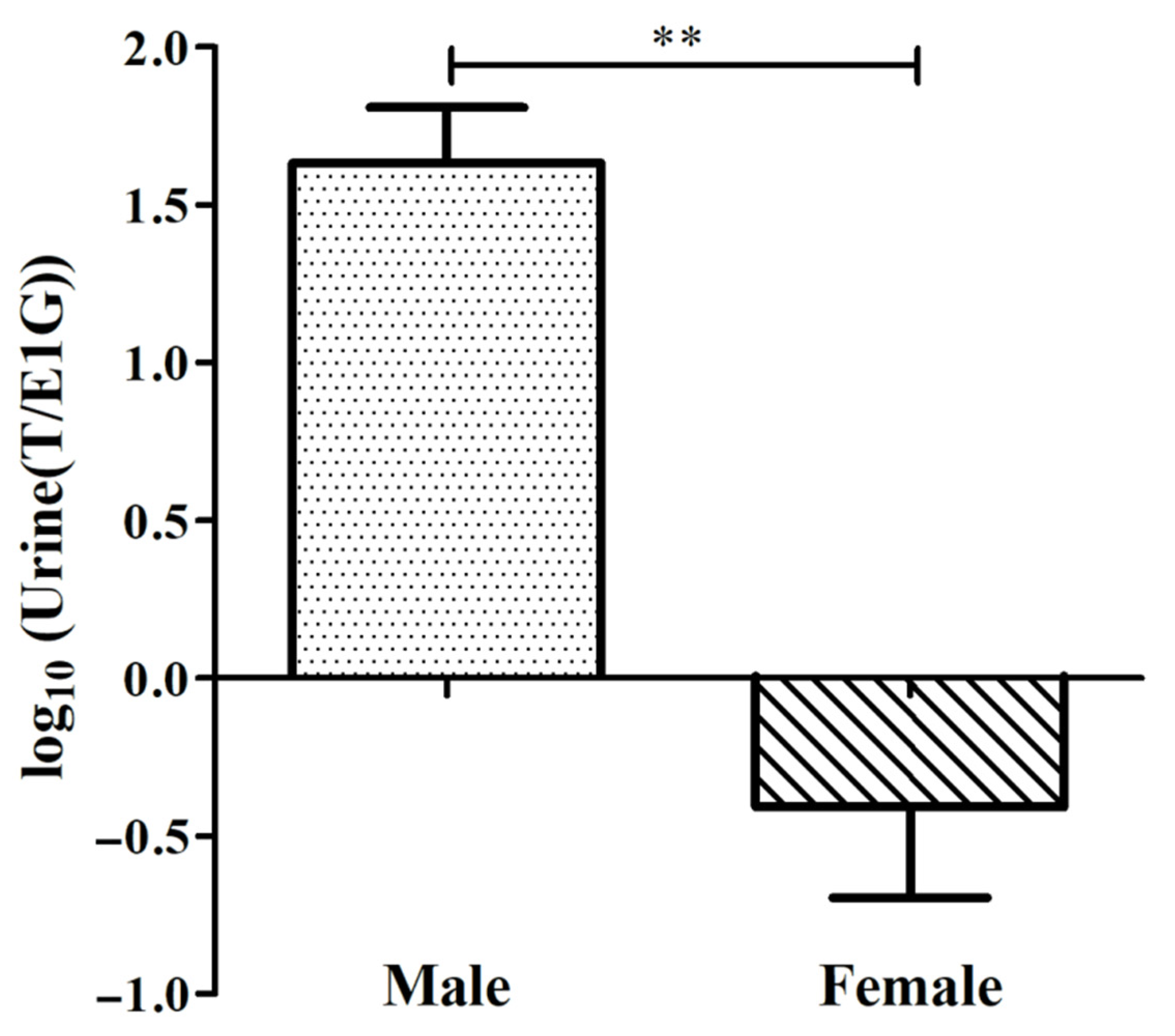
3.2. Verification of the Determination of CGSs’ Sex Using the T/E1G Ratio during the Non-Breeding Season
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Browne, R.K.; Wang, Z.H.; Okada, S.; Hime, P.; McMillan, A.; Wu, M.Y.; Diaz, R.; McGinnity, D.; Briggler, J.T. The giant salamanders (Cryptobranchidae): Part B. Biogeography, ecology and reproduction. Amphib. Reptile Conserv. 2014, 5, 30–50. [Google Scholar]
- Liang, G.; Geng, B.R.; Zhao, E.M. Andrias Davidianus. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Available online: https://www.iucnredlist.org/species/1272/3375181 (accessed on 10 March 2022).
- Gao, K.Q.; Shubin, N.H. Earliest known crown-group salamanders. Nature 2003, 422, 424–428. [Google Scholar] [CrossRef]
- Pyron, R.A.; Wiens, J.J. A Large-Scale Phylogeny of Amphibia Including Over 2800 Species, and a Revised Classification of Extant Frogs, Salamanders, and Caecilians. Mol. Phylogenet. Evol. 2011, 61, 543–583. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.H.; Tong, F.; Song, Y.J.; Wang, H.; Du, M.L.; Ji, H.B. Observation of the breeding behavior of the Chinese giant salamander (Andrias davidianus) using a digital monitoring system. Animals 2018, 8, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijspeert, A.J.; Crespi, A.; Ryczko, D.; Cabelguen, J.M. From swimming to walking with a salamander robot driven by a spinal cord model. Science 2007, 315, 1416–1420. [Google Scholar] [CrossRef] [Green Version]
- CITES. Convention on International Trade in Endangered Species of Wild Fauna and Flora. Appendices I, II and III. 2014. Available online: http://cites.org/eng/app/appendices.php (accessed on 11 March 2022).
- IUCN. IUCN Red List of Threatened Species v. 2012. Available online: http://www.iucnredlist.org (accessed on 10 March 2022).
- Zhang, P.; Chen, Y.Q.; Liu, Y.F.; Zhou, H.; Qu, L.H. The Complete Mitochondrial Genome of the Chinese giant salamander, Andrias Davidianus (Amphibia: Caudata). Gene 2003, 311, 93–98. [Google Scholar] [CrossRef]
- Meng, Y.; Ma, J.; Jiang, N.; Zeng, L.B.; Xiao, H.B. Pathological and microbiological findings from mortality of the Chinese giant salamander (Andrias Davidianus). Arch. Virol. 2014, 159, 1403–1412. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, Y.D.; Zhou, Y.; Liu, W.Z.; Ma, J.; Meng, Y.; Xie, C.X.; Zeng, L.B. Characterization of Chinese giant salamander iridovirus tissue tropism and inflammatory response after infection. Dis. Aquat. Organisms. 2015, 114, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, A.A.; Turvey, S.T.; Zhou, F.; Meredith, H.M.R.; Guan, W.; Liu, X.L.; Sun, C.M.; Wang, Z.Q.; Wu, M.Y. Development of the Chinese giant salamander Andrias davidianus farming industry in Shaanxi Province, China: Conservation threats and opportunities. Oryx 2015, 50, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.W.; Fu, J.; Upton, D.E.; Lema, T.D.; Zhao, E.M. Genetic variability among endangered Chinese giant salamanders, Andrias davidianus. Mol. Ecol. 2010, 9, 1539–1547. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Zhang, K.J.; Wang, Z.H.; Ding, Y.Z.; Wu, W.; Huang, S. The decline of the Chinese giant salamander Andrias davidianus and implications for its conservation. Oryx 2004, 38, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.J.; Zhang, L.; Zhao, H.; Zhao, Q.; Deng, J.; Kong, F.; Jiang, W.; Zhang, H.X.; Liu, H.; Kouba, A. Abiotic and biotic influences on the movement of reintroduced Chinese giant salamanders (Andrias davidianus) in two montane rivers. Animals 2021, 11, 1480. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Willers, R.W. Status and trends of amphibian declines and extinctions world-wide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Jiang, W.; Wang, Q.J.; Zhao, H.; Zhang, H.X.; Marcec, R.M.; Willard, S.T.; Kouba, A.J. Reintroduction and post-release survival of a living fossil: The Chinese giant salamander. PLoS ONE 2016, 11, e0156715. [Google Scholar] [CrossRef]
- Li, P.Q.; Zhu, B.C.; Wang, Y.F.; Xiang, X.J. Sex identification of Chinese giant salamanders (Andrias davidianus) by Doppler B-ultrasoud method. J. Biol. 2010, 27, 94–96. (In Chinese) [Google Scholar]
- Zang, K.J.; Wang, X.M.; Wu, W.; Wang, Z.H.; Huang, S. Advances in conservation biology of Chinese giant salamander. Biodivers. Sci. 2002, 3, 291–297. (In Chinese) [Google Scholar]
- Bauer, B.; Palme, R.; Machatschke, I.H.; Dittami, J.; Huber, S. Noninvasive measurement of adrenocortical and gonadal activity in male and female guinea pigs (Cavia aperea f. porcellus). Gen. Comp. Endocrinol. 2008, 156, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Wasser, S.K.; Wildt, D.E.; Graham, L.H. Comparative aspects of steroid hormone metabolism and ovarian activity in felids, measured noninvasively in feces. Biol. Reprod. 1994, 51, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, J.F.; Bancroft, K.; Kincy, V. Pregnancy and ovulation detection in bison (Bison bison) assessed by means of urinary and fecal steroids. J. Wildlife Dis. 1992, 28, 590–597. [Google Scholar] [CrossRef]
- Cry, N.E.; Romero, L.M. Fecal glucocorticoid metabolites of experimentally stressed captive and free-living starlings: Implications for conservation research. Gen. Comp. Endocrinol. 2008, 158, 20–28. [Google Scholar] [CrossRef]
- Goymann, W. Noninvasive monitoring of hormones in bird droppings: Physiological validation, sampling, extraction, sex differences, and the influence of diet on hormone metabolite levels. Ann. N. Y. Acad. Sci. 2005, 1046, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.M.; Molinia, F.C.; Bishop, P.J.; Cree, A. Urinary hormone analysis assists reproductive monitoring and sex identification of bell frogs (Litoria raniformis). Theriogenology 2009, 72, 663–671. [Google Scholar] [CrossRef]
- Narayan, E.J.; Molinia, F.C.; Christi, K.S.; Morley, C.G.; Cockrem, J.F. Annual cycles of urinary reproductive steroid concentrations in wild and captive endangered Fijian ground frogs (Platymantis vitiana). Gen. Comp. Endocrinol. 2009, 166, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Narayan, E.J.; Molinia, F.C.; Kindermann, C.; Cockrem, J.F.; Hero, J. Urinary corticosterone responses to capture and toe-clipping in the cane toad (Rhinella marina) indicate that toe-clipping is a stressor for amphibians. Gen. Comp. Endocrinol. 2011, 174, 238–245. [Google Scholar] [CrossRef]
- Szymanski, D.C.; Gist, D.H.; Roth, T.L. Anuran gender identification by fecal steroid analysis. Zoo Biol. 2006, 25, 35–46. [Google Scholar] [CrossRef]
- Guntrum, E.B.; Haley, A.M.; Margulis, S.W. Characterization of cycling in a Hoolock Gibbon (Hoolock leuconedys). Folia Primatol. 2021, 92, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Margulis, S.W.; Hálfdanardóttir, M.R. Hormones and color change in female White-Cheeked Gibbons, Nomascus leucogenys. Int. J. Primatol. 2021, 42, 201–219. [Google Scholar] [CrossRef]
- Sattler, R.; Bishop, A.; Woodie, K.; Polasek, L. Characterizing estrus by trans-abdominal ultrasounds, fecal estrone-3-glucuronide, and vaginal cytology in the Steller sea lion (Eumetopias jubatus). Theriogenology 2018, 120, 25–32. [Google Scholar] [CrossRef]
- Li, P.Q.; Xiang, X.J.; Zhu, B.C. Preliminary study on the reproductive hormones of Chinese giant salamander Andrias davidianus. J. Biol. 2008, 03, 30–32. (In Chinese) [Google Scholar]
- Jiang, W.; Wang, Q.J.; Zhao, H.; Zhang, H.X.; Andy, K.; Zhang, L.; Ruth, M. Effectiveness of MS-222 as anesthetic agents for Andrias davidianus. Freshwater Fisheries 2014, 44, 94–97. (In Chinese) [Google Scholar]
- Klein, S.S.; Bogart, K. Achieving sex equity in education: A comparison at Pre-and Post-Secondary levels. Equity Excell. Educ. 1987, 23, 114–122. [Google Scholar] [CrossRef]
- Nakamura, M. The mechanism of sex determination in vertebrates-are sex steroids the key-factor? J. Exp. Zool. A Ecol. Genet. Physiol. 2010, 313, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M. Is a sex-determining gene(s) necessary for sex-determination in amphibians? Steroid hormones may be the key factor. Sex. Dev. 2013, 7, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, L.F.; Cooke, D.G.; Brown, S. The Use of Estrone-3-Glucuronide and Pregnanediol-3-Glucuronide Excretion Rates to Navigate the Continuum of Ovarian Activity. Front. Public. Health 2018, 31, 153. [Google Scholar] [CrossRef]
- Zou, Z.Y.; Wang, H.W.; Ou, D.S.; He, W. Seasonal changes of sexual hormone level in Andrias davidianus urinarys and its relationship with breeding. J. Hydroecol. 2012, 33, 128–131. (In Chinese) [Google Scholar]
- Kalz, B.; Jewgenow, K.; Fickel, J. Structure of an otter (Lutra lutra) population in Germany-results of DNA and hormone analyses from faecal samples. Mamm. Biol. 2006, 71, 321–335. [Google Scholar] [CrossRef]
- Prado-Ortiz, L.E.; Valdespino, C.; Romano, M.; González-Romero, A. Quantification of immunoreactive testosterone and estradiol-17ß metabolites to identify the sex of Neotropical otters (Lontra longicaudis annectens) in the field. Anim. Reprod. Sci. 2020, 222, 106607. [Google Scholar] [CrossRef]
- Lynch, K.S.; Wilczynski, W. Gonadal steroids vary with reproductive stage in a tropically breeding anuran. Gen. Comp. Endocrinol. 2005, 143, 51–56. [Google Scholar] [CrossRef]
- Medina, M.F.; Ines, R.; Crespo, C.A.; Gonzalez-Calvar, S.; Fenandez, S.L. Changes in serum sex steroid levels throught the reproductive cycle of Bufo arenarum females. Gen. Comp. Endocrinol. 2004, 136, 143–151. [Google Scholar] [CrossRef]
- Mendoca, M.T.; Licht, P.; Ryan, M.J.; Barnes, R. Changes in hormone levels in relation to breeding behavior in male bullfrogs (Rana catesbeiana) at the individual and population levels. Gen. Comp. Endocrinol. 1985, 58, 270–279. [Google Scholar] [CrossRef]
- Rastogi, R.K.; Iela, L.; Delrio, G.; Bagnara, J.T. Reproduction in the Mexican leaf frog, Pachymedusa dacnicolor II. The Male. Gen. Comp. Endocrinol. 1986, 62, 23–35. [Google Scholar] [CrossRef]
- Dehnhard, M.; Naidenko, S.; Frank, A.; Braun, B.; Goritz, F.; Jewgenow, K. Non-invasive monitoring of hormones: A tool to improve reproduction in captive breeding of the Eurasian lynx hormone monitoring in breeding programmes of mammals. Reprod. Dom. Anim. 2008, 43, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Pribbenow, S.; Wachter, B.; Ludwig, C.; Weigold, A.; Dehnhard, M. Validation of an enzyme-immunoassay for the non-invasive monitoring of faecal testosterone metabolites in male cheetahs (Acinonyx jubatus). Gen. Compar. Endocrinol. 2016, 228, 40–47. [Google Scholar] [CrossRef]
- Anderson, L.L. Pigs. In Reproduction in Farm Animals; Hafez, E.S.E., Ed.; Lea & Febiger: Philadelphia, PA, USA, 1992; pp. 343–360. [Google Scholar]
- Hafez, E.S.E. (Ed.) Horses. In Reproduction in Farm Animals; Lea & Febiger: Philadelphia, PA, USA, 1992; pp. 361–384. [Google Scholar]
- Lin, Y.; Gan, C.; Wang, Y. Evaluation of the Effect of releasing Giant Salamanders into the Shuaishui River. J. Hydroecol. 2017, 38, 88–96. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | No. | T | E1G | T/E1G | log10(T/E1G) | ||
|---|---|---|---|---|---|---|---|
| OD | Conc. (pg/mL) | OD | Conc. (pg/mL) | ||||
| Male | M1 | 1.179 | 3052.5 | 1.580 | 10.1 | 302.736 | 2.481 |
| M2 | 1.178 | 3060.3 | 1.555 | 17.5 | 174.675 | 2.242 | |
| M3 | 0.875 | 6754.5 | 1.552 | 18.4 | 367.470 | 2.565 | |
| Female | F1 | 2.151 | 82.1 | 1.159 | 135.5 | 0.606 | −0.218 |
| F2 | 2.180 | 62.8 | 0.866 | 264.2 | 0.238 | −0.623 | |
| F3 | 2.186 | 59.0 | 1.011 | 193.2 | 0.306 | −0.514 | |
| Sex | No. | T (pg/mL) | E1G (pg/mL) | ||||
|---|---|---|---|---|---|---|---|
| OD | Ind. Conc. | Aver. Conc. | OD | Ind. Conc. | Aver. Conc. | ||
| Male | SA1 | 2.217 | 138.6 | 1175.5 ± 936.9 | 1.608 | 0.8 | 52.1 ± 102.9 |
| SA2 | 2.089 | 438.6 | 1.576 | 38.2 | |||
| SA3 | 1.986 | 790.6 | 1.607 | 2.6 | |||
| SA4 | 2.146 | 285.7 | 1.607 | 1.5 | |||
| SA5 | 1.792 | 1771.6 | 1.541 | 71.7 | |||
| SA6 | 1.623 | 3066.4 | 1.560 | 54.1 | |||
| SA7 | 1.832 | 1527.6 | 1.490 | 118.8 | |||
| A1 | 1.750 | 2049.9 | 1.421 | 182.7 | |||
| A2 | 2.083 | 454.9 | 1.585 | 28.9 | |||
| A3 | 1.887 | 1231.2 | 1.592 | 21.3 | |||
| Female | SA8 | 2.275 | 51.7 | 149.4 ± 203.2 | 1.494 | 115.6 | 222.9 ± 218.7 |
| SA9 | 2.241 | 98.7 | 1.465 | 141.7 | |||
| SA10 | 2.292 | 32.1 | 1.573 | 40.7 | |||
| SA11 | 2.026 | 639.8 | 1.487 | 122.0 | |||
| SA12 | 2.141 | 299.0 | 1.487 | 121.6 | |||
| A4 | 2.266 | 63.0 | 1.305 | 294.2 | |||
| A5 | 2.340 | 1.8 | 1.344 | 255.2 | |||
| A6 | 2.280 | 45.2 | 0.930 | 769.6 | |||
| A7 | 2.231 | 113.0 | 1.461 | 145.8 | |||
| Sex | No. | T/E1G | log10(T/E1G) |
|---|---|---|---|
| Male | SA05 | 178.493 | 2.252 |
| SA06 | 11.477 | 1.060 | |
| A07 | 309.311 | 2.490 | |
| SA08 | 186.089 | 2.270 | |
| SA09 | 24.707 | 1.393 | |
| SA10 | 56.727 | 1.754 | |
| SA12 | 12.856 | 1.109 | |
| A01 | 11.223 | 1.050 | |
| A02 | 15.734 | 1.197 | |
| A03 | 57.703 | 1.761 | |
| Female | SA01 | 0.442 | −0.355 |
| SA02 | 0.697 | −0.157 | |
| SA03 | 0.788 | −0.103 | |
| SA04 | 5.243 | 0.720 | |
| SA11 | 2.460 | 0.391 | |
| A04 | 0.214 | −0.670 | |
| A05 | 0.007 | −2.155 | |
| A06 | 0.059 | −1.229 | |
| A07 | 0.775 | −0.111 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Huang, J.; Zhao, H.; Deng, J.; Kong, F.; Zhang, H.; Wang, Q. Using the Ratio of Urine Testosterone to Estrone-3-Glucuronide to Identify the Sex of Chinese Giant Salamanders (Andrias davidianus). Animals 2022, 12, 1112. https://doi.org/10.3390/ani12091112
Zhang J, Huang J, Zhao H, Deng J, Kong F, Zhang H, Wang Q. Using the Ratio of Urine Testosterone to Estrone-3-Glucuronide to Identify the Sex of Chinese Giant Salamanders (Andrias davidianus). Animals. 2022; 12(9):1112. https://doi.org/10.3390/ani12091112
Chicago/Turabian StyleZhang, Jianlu, Jiqin Huang, Hu Zhao, Jie Deng, Fei Kong, Hongxing Zhang, and Qijun Wang. 2022. "Using the Ratio of Urine Testosterone to Estrone-3-Glucuronide to Identify the Sex of Chinese Giant Salamanders (Andrias davidianus)" Animals 12, no. 9: 1112. https://doi.org/10.3390/ani12091112
APA StyleZhang, J., Huang, J., Zhao, H., Deng, J., Kong, F., Zhang, H., & Wang, Q. (2022). Using the Ratio of Urine Testosterone to Estrone-3-Glucuronide to Identify the Sex of Chinese Giant Salamanders (Andrias davidianus). Animals, 12(9), 1112. https://doi.org/10.3390/ani12091112