
Dynamic of Hepatitis E Virus (HEV) Shedding in Pigs

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samplings
2.2. RNA Extraction and Quantitative Real-Time Reverse Transcription PCR
2.3. Statistical Analysis
2.4. Sequencing
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Smith, D.B.; Simmonds, P. Classification and Genomic Diversity of Enterically Transmitted Hepatitis Viruses. Cold Spring Harb. Perspect. Med. 2018, 8, a031880. [Google Scholar] [CrossRef]
- Kamar, N.; Pischke, S. Acute and Persistent Hepatitis E Virus Genotype 3 and 4 Infection: Clinical Features, Pathogenesis, and Treatment. Cold Spring Harb. Perspect. Med. 2019, 9, a031872. [Google Scholar] [CrossRef]
- Aspinall, E.J.; Couturier, E.; Faber, M.; Said, B.; Ijaz, S.; Tavoschi, L.; Takkinen, J.; Adlhoch, C.; on behalf of the country experts. Hepatitis E virus infection in Europe: Surveillance and descriptive epidemiology of confirmed cases, 2005 to 2015. Euro. Surveill. 2017, 22, 30561. [Google Scholar] [CrossRef] [Green Version]
- Pavio, N.; Doceul, V.; Bagdassarian, E.; Johne, R. Recent knowledge on hepatitis E virus in Suidae reservoirs and transmission routes to human. Vet. Res. 2017, 48, 78. [Google Scholar] [CrossRef] [Green Version]
- Primadharsini, P.P.; Nagashima, S.; Okamoto, H. Genetic Variability and Evolution of Hepatitis E Virus. Viruses 2019, 11, 456. [Google Scholar] [CrossRef] [Green Version]
- Treagus, S.; Wright, C.; Baker-Austin, C.; Longdon, B.; Lowther, J. The Foodborne Transmission of Hepatitis E Virus to Humans. Food Environ. Virol. 2021, 13, 127–145. [Google Scholar] [CrossRef]
- Salines, M.; Andraud, M.; Rose, N. From the epidemiology of hepatitis E virus (HEV) within the swine reservoir to public health risk mitigation strategies: A comprehensive review. Vet. Res. 2017, 48, 31. [Google Scholar] [CrossRef] [Green Version]
- Salines, M.; Rose, N.; Andraud, M. Tackling hepatitis E virus spread and persistence on farrow-to-finish pig farms: Insights from a stochastic individual-based multi-pathogen model. Epidemics 2020, 30, 100369. [Google Scholar] [CrossRef]
- Andraud, M.; Casas, M.; Pavio, N.; Rose, N. Early-life hepatitis e infection in pigs: The importance of maternally-derived antibodies. PLoS ONE 2014, 9, e105527. [Google Scholar] [CrossRef]
- Kasorndorkbua, C.; Thacker, B.J.; Halbur, P.G.; Guenette, D.K.; Buitenwerf, R.M.; Royer, R.L.; Meng, X.J. Experimental infection of pregnant gilts with swine hepatitis E virus. Can. J. Vet. Res. 2003, 67, 303–306. [Google Scholar]
- Casas, M.; Cortes, R.; Pina, S.; Peralta, B.; Allepuz, A.; Cortey, M.; Casal, J.; Martin, M. Longitudinal study of hepatitis E virus infection in Spanish farrow-to-finish swine herds. Vet. Microbiol. 2011, 148, 27–34. [Google Scholar] [CrossRef]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’Amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; et al. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef]
- Feurer, C.; Le Roux, A.; Rossel, R.; Barnaud, E.; Dumarest, M.; Garry, P.; Pavio, N. High load of hepatitis E viral RNA in pork livers but absence in pork muscle at French slaughterhouses. Int. J. Food Microbiol. 2018, 264, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Di Bartolo, I.; Diez-Valcarce, M.; Vasickova, P.; Kralik, P.; Hernandez, M.; Angeloni, G.; Ostanello, F.; Bouwknegt, M.; Rodriguez-Lazaro, D.; Pavlik, I.; et al. Hepatitis E virus in pork production chain in Czech Republic, Italy, and Spain, 2010. Emerg. Infect. Dis. 2012, 18, 1282–1289. [Google Scholar] [CrossRef]
- Rose, N.; Pavio, N. Epidemiology of hepatitis E virus in pigs: The way forward to limit consumer exposure (in french). Journées Rech. Porc. 2014, 46, 159–168. [Google Scholar]
- Satou, K.; Nishiura, H. Transmission dynamics of hepatitis E among swine: Potential impact upon human infection. BMC Vet. Res. 2007, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kamili, S.; Krawczynski, K. Quantitative detection of hepatitis E virus RNA and dynamics of viral replication in experimental infection. J. Viral. Hepat. 2006, 13, 835–839. [Google Scholar] [CrossRef]
- Salines, M.; Demange, A.; Stephant, G.; Renson, P.; Bourry, O.; Andraud, M.; Rose, N.; Pavio, N. Persistent viremia and presence of hepatitis E virus RNA in pig muscle meat after experimental co-infection with porcine reproductive and respiratory syndrome virus. Int. J. Food Microbiol. 2019, 292, 144–149. [Google Scholar] [CrossRef]
- De Sabato, L.; Amoroso, M.G.; Ianiro, G.; Esposito, C.; De Grossi, L.; Fusco, G.; Barone, A.; Martini, E.; Ostanello, F.; Di Bartolo, I. Detection of Hepatitis E Virus in Livers and Muscle Tissues of Wild Boars in Italy. Food Environ. Virol. 2020, 12, 1–8. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Huang, F.F.; Haqshenas, G.; Guenette, D.K.; Halbur, P.G.; Schommer, S.K.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Detection by reverse transcription-PCR and genetic characterization of field isolates of swine hepatitis E virus from pigs in different geographic regions of the United States. J. Clin. Microbiol. 2002, 40, 1326–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Deus, N.; Casas, M.; Peralta, B.; Nofrarias, M.; Pina, S.; Martin, M.; Segales, J. Hepatitis E virus infection dynamics and organic distribution in naturally infected pigs in a farrow-to-finish farm. Vet. Microbiol. 2008, 132, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Withenshaw, S.M.; Grierson, S.S.; Smith, R.P. Study of Animal Mixing and the Dynamics of Hepatitis E Virus Infection on a Farrow-to-Finish Pig Farm. Animals 2022, 12, 272. [Google Scholar] [CrossRef]
- Kantala, T.; Oristo, S.; Heinonen, M.; von Bonsdorff, C.H.; Maunula, L. A longitudinal study revealing hepatitis E virus infection and transmission at a swine test station. Res. Vet. Sci. 2013, 95, 1255–1261. [Google Scholar] [CrossRef]
- Kanai, Y.; Tsujikawa, M.; Yunoki, M.; Nishiyama, S.; Ikuta, K.; Hagiwara, K. Long-term shedding of hepatitis E virus in the feces of pigs infected naturally, born to sows with and without maternal antibodies. J. Med. Virol. 2010, 82, 69–76. [Google Scholar] [CrossRef]
- Andraud, M.; Dumarest, M.; Cariolet, R.; Aylaj, B.; Barnaud, E.; Eono, F.; Pavio, N.; Rose, N. Direct contact and environmental contaminations are responsible for HEV transmission in pigs. Vet. Res. 2013, 44, 102. [Google Scholar] [CrossRef] [Green Version]
- Walachowski, S.; Dorenlor, V.; Lefevre, J.; Lunazzi, A.; Eono, F.; Merbah, T.; Eveno, E.; Pavio, N.; Rose, N. Risk factors associated with the presence of hepatitis E virus in livers and seroprevalence in slaughter-age pigs: A retrospective study of 90 swine farms in France. Epidemiol. Infect. 2014, 142, 1934–1944. [Google Scholar] [CrossRef]
- Ianiro, G.; Chelli, E.; De Sabato, L.; Monini, M.; Ostanello, F.; Di Bartolo, I. Long-term surveillance for hepatitis E virus in an Italian two-site farrow-to-finish swine farm. Zoonoses Public Health 2021, 68, 474–482. [Google Scholar] [CrossRef]
- Wang, H.; Karlsson, M.; Lindberg, M.; Nystrom, K.; Norder, H. Hepatitis E virus strains infecting Swedish domestic pigs are unique for each pig farm and remain in the farm for at least 2 years. Transbound. Emerg. Dis. 2019, 66, 1314–1323. [Google Scholar] [CrossRef]
- Meester, M.; Tobias, T.J.; Bouwknegt, M.; Kusters, N.E.; Stegeman, J.A.; van der Poel, W.H.M. Infection dynamics and persistence of hepatitis E virus on pig farms—A review. Porc. Health Manag. 2021, 7, 16. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
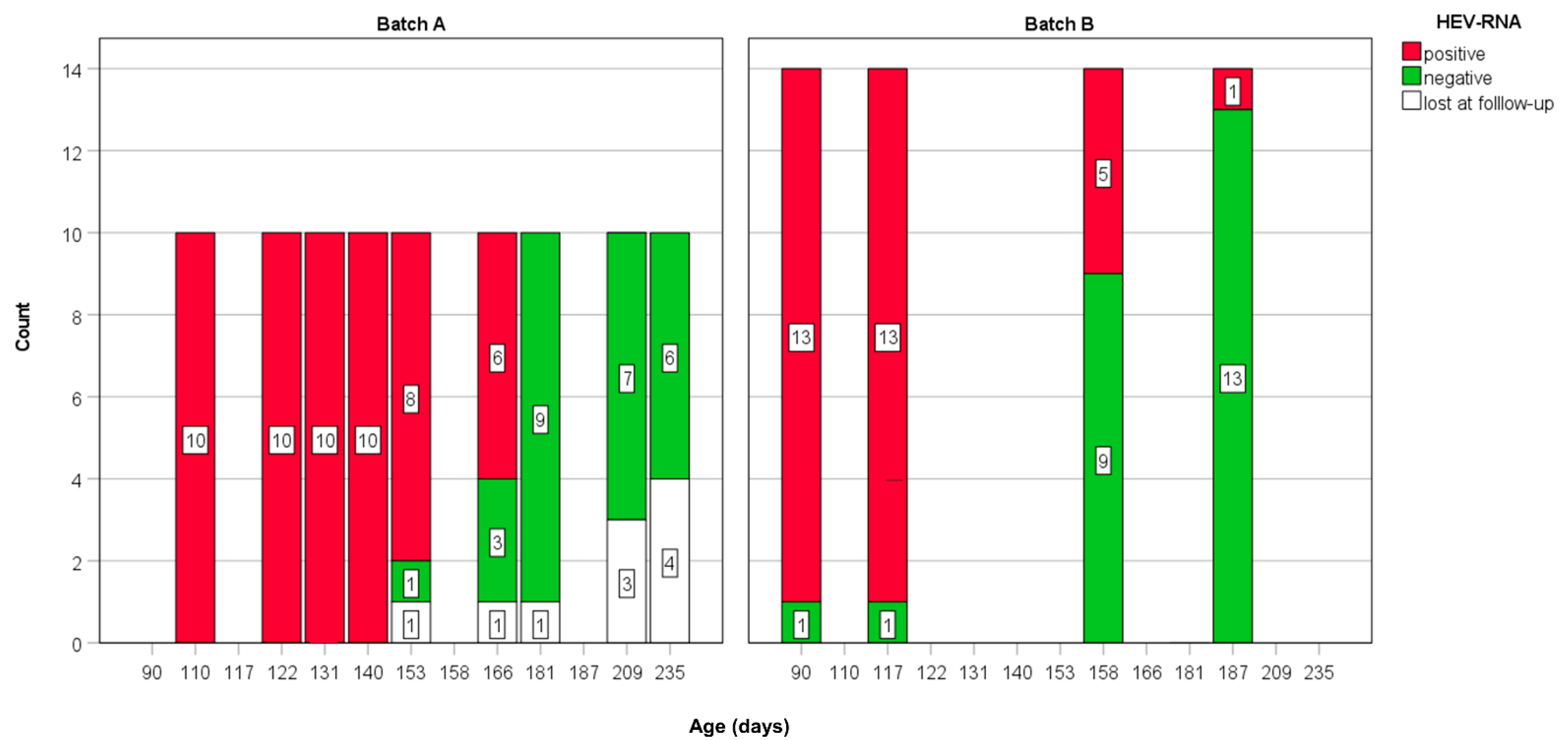
| Batch | Sampling (T) | Age (Days) | No. of Days from the First Sampling (T1) | HEV-RNA | ||
|---|---|---|---|---|---|---|
| Total Tested | Positive | Prevalence % | ||||
| A | 1 | 110 | 0 | 10 | 10 | 100 |
| 2 | 122 | 12 | 10 | 10 | 100 | |
| 3 | 131 | 21 | 10 | 10 | 100 | |
| 4 | 140 | 30 | 10 | 10 | 100 | |
| 5 | 153 | 43 | 9 | 8 | 88.9 | |
| 6 | 166 | 56 | 9 | 6 | 66.7 | |
| 7 | 181 | 71 | 9 | 0 | 0 | |
| 8 | 209 | 99 | 7 | 0 | 0 | |
| 9 | 235 | 125 | 6 | 0 | 0 | |
| B | 1 | 90 | 0 | 13 | 13 | 100 |
| 2 | 117 | 27 | 13 | 13 | 100 | |
| 3 | 158 | 68 | 13 | 5 | 38.5 | |
| 4 | 187 | 97 | 13 | 1 | 7.7 | |
| Age of Animals (Days) | |||||
|---|---|---|---|---|---|
| Batch | Mean | Std. Dev. | Minimum | Maximum | Median |
| A | 143.29 | 23.33 | 110 | 166 | 140.0 |
| B | 138.00 | 37.60 | 90 | 187 | 137.5 |
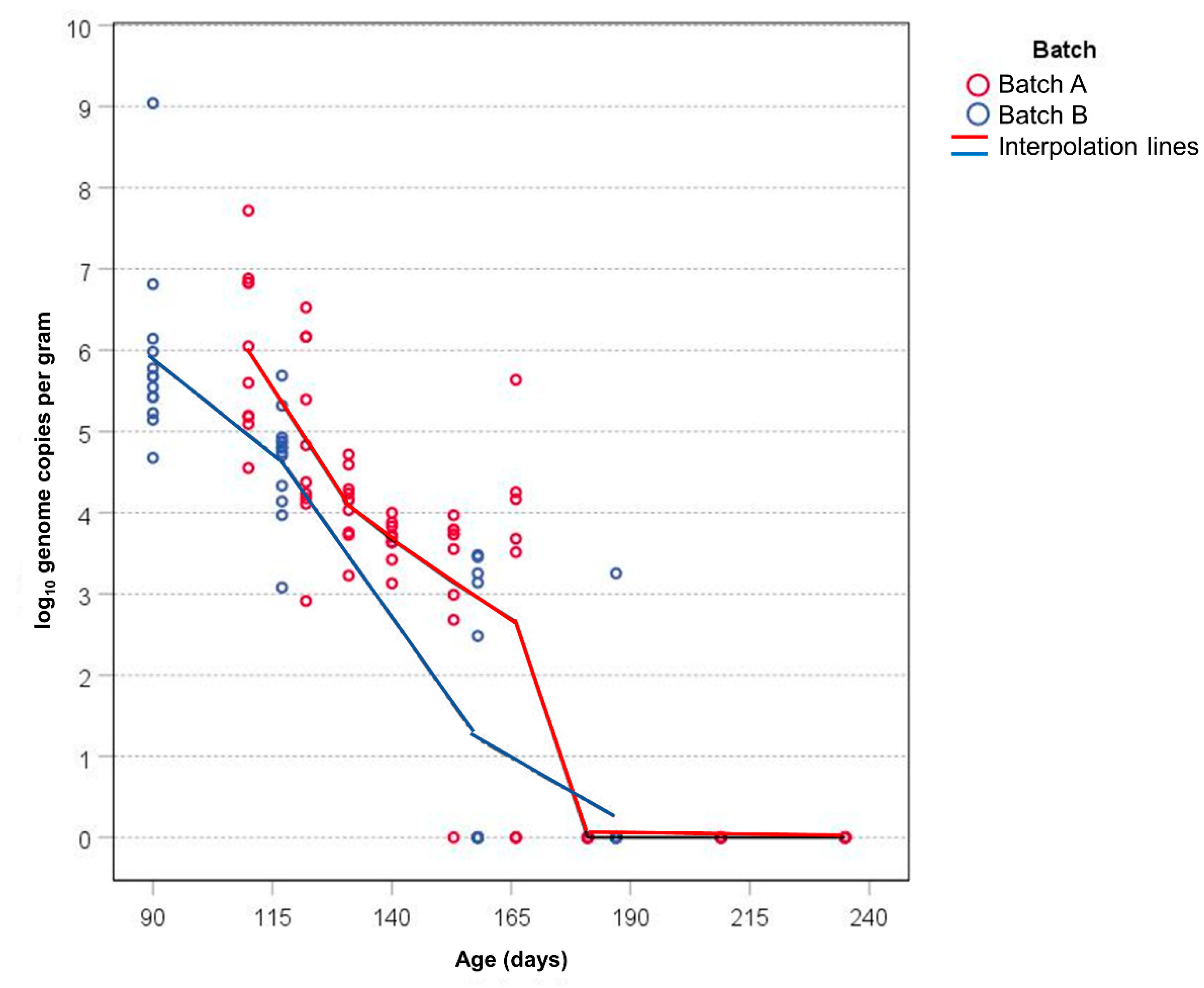
| Animals | GC/g 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Batch | Sampling (T) | Age (Days) | No. Tested | HEV Positive | Mean 1 | Std. Deviation | Minimum | Maximum | Median |
| A | 1 | 110 | 10 | 10 | 5.99 | 1.03 | 4.55 | 7.72 | 5.82 |
| 2 | 122 | 10 | 10 | 4.89 | 1.15 | 2.91 | 6.53 | 4.60 | |
| 3 | 131 | 10 | 10 | 4.09 | 0.44 | 3.23 | 4.72 | 4.16 | |
| 4 | 140 | 10 | 10 | 3.67 | 0.25 | 3.13 | 4.00 | 3.71 | |
| 5 | 153 | 9 | 8 | 3.14 | 1.25 | 0.00 | 3.97 | 3.73 | |
| 6 | 166 | 8 | 6 | 2.66 | 2.29 | 0.00 | 5.64 | 3.59 | |
| 7 | 181 | 9 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 8 | 209 | 7 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 9 | 235 | 6 | 0 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| B | 1 | 90 | 13 | 13 | 5.89 | 1.08 | 4.67 | 9.04 | 5.67 |
| 2 | 117 | 13 | 13 | 4.63 | 0.65 | 3.08 | 5.69 | 4.80 | |
| 3 | 158 | 13 | 5 | 1.22 | 1.62 | 0.00 | 3.48 | 0.00 | |
| 4 | 187 | 13 | 1 | 0.25 | 0.90 | 0.00 | 3.25 | 0.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ianiro, G.; Monini, M.; De Sabato, L.; Chelli, E.; Cerini, N.; Ostanello, F.; Di Bartolo, I. Dynamic of Hepatitis E Virus (HEV) Shedding in Pigs. Animals 2022, 12, 1063. https://doi.org/10.3390/ani12091063
Ianiro G, Monini M, De Sabato L, Chelli E, Cerini N, Ostanello F, Di Bartolo I. Dynamic of Hepatitis E Virus (HEV) Shedding in Pigs. Animals. 2022; 12(9):1063. https://doi.org/10.3390/ani12091063
Chicago/Turabian StyleIaniro, Giovanni, Marina Monini, Luca De Sabato, Eleonora Chelli, Natalino Cerini, Fabio Ostanello, and Ilaria Di Bartolo. 2022. "Dynamic of Hepatitis E Virus (HEV) Shedding in Pigs" Animals 12, no. 9: 1063. https://doi.org/10.3390/ani12091063
APA StyleIaniro, G., Monini, M., De Sabato, L., Chelli, E., Cerini, N., Ostanello, F., & Di Bartolo, I. (2022). Dynamic of Hepatitis E Virus (HEV) Shedding in Pigs. Animals, 12(9), 1063. https://doi.org/10.3390/ani12091063