
Behavioral Assessment of Six Reptile Species during a Temporary Zoo Closure and Reopening



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing
2.2. Behavioral Observations
2.3. Data Analysis
3. Results
3.1. Catalina Island Rattlesnake
3.2. European Glass Lizard
3.3. Beaded Lizard
3.4. Sonoran Spiny-Tailed Iguana
3.5. Arrau Turtle
3.6. Dwarf Caiman
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conde, D.A.; Colchero, F.; Gusset, M.; Pearce-Kelly, P.; Byers, O.; Flesness, N.; Browne, R.K.; Jones, O.R. Zoos through the lens of the IUCN Red List: A global metapopulation approach to support conservation breeding programs. PLoS ONE 2013, 8, e80311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar]
- Lambert, H.; Carder, G.; D’Cruze, N. Given the cold shoulder: A review of the scientific literature for evidence of reptile sentience. Animals 2019, 9, 821. [Google Scholar] [CrossRef] [Green Version]
- Avila-Villegas, H.; Martins, M.; Arnaud, G. Feeding ecology of the endemic rattleless rattlesnake, Crotalus catalinensis, of Santa Catalina Island, Gulf of California, Mexico. Copeia 2007, 1, 80–84. [Google Scholar] [CrossRef]
- Reynoso, V.H.; Vázquez-Cruz, M.; Rivera-Arroyo, R.C.; Zarza-Franco, E.; Grant, T.D. Ctenosaura macrolopha. In The IUCN Red List of Threatened Species 2020: E.T174476A1414477; International Union for Conservation of Nature and Natural Resources, 2020; Available online: https://repository.sandiegozoo.org/handle/20.500.12634/601 (accessed on 10 April 2020).
- Magnusson, W.E.; Campos, Z.M.S. Cuvier’s smooth-fronted caiman Paleosuchus palpebrosus. In Crocodiles. Status Survey and Conservation Action Plan, 3rd ed.; Manolis, S.C., Stevenson, C., Eds.; Crocodile Specialist Group: Darwin, Australia, 2010; pp. 40–42. [Google Scholar]
- Arnaud, G.; Sandoval, S.; Escobar-Flores, J.G.; Gomez-Muñoz, V.M.; Burguete, J.L. Thermal ecology of the rattlesnake Crotalus catalinensis from Santa Catalina island, gulf of California. Acta Univ. 2018, 28, 39–46. [Google Scholar] [CrossRef]
- Rifai, L.; Baker, M.A.; Shafei, D.A.; Disi, A.; Mahasneh, A.; Amr, Z. Pseudopus apodus (PALLAS, 1775) from Jordan, with notes on its ecology. Herpetozoa 2005, 18, 133–140. [Google Scholar]
- Beck, D.D.; Lowe, C.H. Ecology of the Beaded Lizard, Heloderma horridum, in a tropical dry forest in Jalisco, México. J. Herpetol. 1991, 25, 395–406. [Google Scholar] [CrossRef]
- Ferrara, C.R.; Vogt, R.C.; Sousa-Lima, R.S.; Tardio, B.M.R.; Bernardes, V.C.D. Sound communication and social behavior in an amazonian river turtle (podocnemis expansa). Herpetologica 2014, 70, 149–156. [Google Scholar] [CrossRef]
- Oliveira, S.R.; Vaz-Silva, W.; de Souza, M.B.; de Andrade, S.P.; de Morais, A.R. A caecilian (Siphonops sp.) as prey of a dwarf caiman Paleosuchus palpebrosus: An observation from Central Brazil. Herpetol. Notes 2019, 12, 71–73. [Google Scholar]
- Sheetz, K.; Lemos-Espinal, J.A.; Smith, G.R. Ctenosaura Macrolopha (mainland spinytail iguana) Diet; Clutch size. Herpetol. Rev. 2007, 38, 77–78. [Google Scholar]
- Lara, N.R.F.; Marques, T.S.; Montelo, K.M.; de Ataídes, Á.G.; Verdade, L.M.; Malvásio, A.; de Camargo, P.B. A trophic study of the sympatric Amazonian freshwater turtle Podocnemis unifilis and Podocnemis expansa (Testudines, Podocnemidae) using carbon and nitrogren stable isotope analyses. Can. J. Zool. 2012, 90, 1394–1401. [Google Scholar] [CrossRef]
- Nagloo, N.; Collin, S.P.; Hemmi, J.M.; Hart, N.S. Spatial resolving power and spectral sensitivity of the saltwater crocodile, Crocodylus porosus, and the freshwater crocodile, Crocodylus johnstoni. J. Exp. Biol. 2016, 219, 1394–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, R.T.; Parker, M.R. Social behavior and pheromonal communication in reptiles. J. Comp. Physiol. 2010, 196, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Weldon, P.J.; Ferguson, M.W. Chemoreception in crocodilians: Anatomy, natural history, and empirical results. Brain Behav. Evol. 1993, 41, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.E.; Christensen-Dalsgaard, J.; Bierman, H. Coupled ears in lizards and crocodilians. Biol. Cybern. 2016, 110, 291–302. [Google Scholar] [CrossRef]
- Leitch, D.B.; Catania, K.C. Structure, innervation and response properties of integumentary sensory organs in crocodilians. J. Exp. Biol. 2012, 215, 4217–4230. [Google Scholar] [CrossRef] [Green Version]
- Young, B.A. Snake bioacoustics: Toward a richer understanding of the behavioral ecology of snakes. Q. Rev. Biol. 2003, 78, 303–325. [Google Scholar] [CrossRef]
- Davey, G. Visitor behavior in zoos: A review. Anthrozoös 2006, 19, 143–157. [Google Scholar] [CrossRef]
- Kratochvil, H.; Schwammer, H. Reducing acoustic disturbances by aquarium visitors. Zoo Biol. 1997, 16, 349–353. [Google Scholar] [CrossRef]
- Leong, H.; Ros, A.F.H.; Oliveira, R.F. Effects of putative stressors in public aquaria on locomotor activity, metabolic rate and cortisol levels in the Mozambique tilapia Oreochromis mossambicus. J. Fish Biol. 2009, 74, 1549–1561. [Google Scholar] [CrossRef]
- Kight, C.R.; Swaddle, J.P. How and why environmental noise impacts animals: An integrative, mechanistic review. Ecol. Lett. 2011, 14, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.R.; Lukas, K.E. Zoo visitor behavior at an African ape exhibit. Visit. Stud. Today 2005, 8, 4–12. [Google Scholar]
- Sherwen, S.L.; Hemsworth, P.H. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, A.; Terry, M.; Freeman, H.; Alba, A.C.; Soltis, J.; Leeds, A. Evaluating the effect of visitor presence on Nile crocodile (Crocodylus niloticus) behavior. J. Zool. Bot. Gard. 2021, 2, 115–129. [Google Scholar] [CrossRef]
- Carter, K.C.; Keane, I.A.; Clifforde, L.M.; Rowden, L.J.; Fieschi-Méric, L.; Michaels, C.J. The Effect of Visitors on Zoo Reptile Behaviour during the COVID-19 Pandemic. J. Zool. Bot. Gard. 2021, 2, 664–676. [Google Scholar] [CrossRef]
- Boultwood, J.; O’Brien, M.; Rose, P. Bold frogs or shy toads? How did the COVID-19 closure of zoological organisations affect amphibian activity? Animals 2021, 11, 1982. [Google Scholar] [CrossRef] [PubMed]
- Kagan, R.; Carter, S.; Allard, S. A universal animal welfare framework for zoos. J. Appl. Anim. Welf. Sci. 2015, 18, S1–S10. [Google Scholar] [CrossRef] [Green Version]
- Burghardt, G.M.; Rand, A.S. Iguanas of the World: Their Behavior, Ecology, and Conservation; Noyes Publications: Park Ridge, NJ, USA, 1982. [Google Scholar]
- Vieira, L.G.; Santos, A.L.; Moura, L.R.; Orpinelli, S.R.; Pereira, K.F.; Lima, F.C. Morphology, development and heterochrony of the carapace of Giant Amazon River Turtle Podocnemis expansa (Testudines, Podocnemidae). Pesqui. Vet. Bras. 2016, 36, 436–446. [Google Scholar] [CrossRef] [Green Version]
- Martin, S. Global diversity of crocodiles (Crocodilia, Reptilia) in freshwater. Hydrobiologia 2008, 595, 587–591. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Zoo, L.P. ZooMonitor (version 3.2). In Mobile Application Software; 2020; Available online: https://zoomonitor.org (accessed on 10 April 2020).
- Hallgren, K.A. Computing Inter-Rater Reliability for Observational Data: An Overview and Turtorial. Tutor. Quant. Methods Psychol. 2012, 8, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, P.; Evans, C.; Coffin, R.; Miller, R.; Nash, S. Using student-centred research to evidence-base exhibition of reptiles and amphibians: Three species-specific case studies. J. Zoo Aquar. Res. 2014, 2, 25–32. [Google Scholar]
- Spain, M.S.; Fuller, G.; Allard, S.M. Effects of habitat modifications on behavioral indicators of welfare for Madagascar giant hognose snakes (Leioheterodon madagascariensis). Anim. Behav. Cogn. 2020, 7, 70–81. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral diversity as a potential indicator of positive animal welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef]
- Di Bitetti, M.S. The Distribution of Grooming among Female Primates: Testing Hypotheses with the Shannon-Wiener Diversity Index. Behaviour 2000, 137, 1517–1540. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Harvey, E. Enclosure use as a measure of behavioural welfare in three zoo-housed African wild dogs Lycaon pictus. J. Zoo Aquar. Res. 2021, 9, 88–93. [Google Scholar]
- Miller, L.J.; Piscane, C.B.; Vicino, G.A. Relationship between behavioural diversity and faecal glucocorticoid metabolites: A case study with cheetahs (Acinonyx jubatus). Anim. Welf. 2016, 25, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Razal, C.B.; Bryant, J.; Miller, L.J. Monitoring the behavioral and adrenal activity of giraffe (Giraffa camelopardalis) to assess welfare during seasonal housing changes. Anim. Behav. Cogn. 2017, 4, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Dickens, M. A statistical formula to quantify the “ spread-of-participation” in group discussion. Speech Monogr. 1955, 22, 28–30. [Google Scholar] [CrossRef]
- Brereton, J.E. Current directions in animal enclosure use studies. J. Zoo Aquar. Res. 2020, 8, 1–9. [Google Scholar]
- Hedeen, S.E. Utilization of space by captive groups of lowland gorillas (Gorilla g. gorilla). Ohio J. Sci. 1982, 82, 27–30. [Google Scholar]
- Plowman, A.B. A note on a modification of the spread of participation index allowing for unequal zones. Appl. Anim. Behav. Sci. 2003, 83, 331–336. [Google Scholar] [CrossRef]
- Prystupczuk, L.; Leynaud, G.C.; Lábaque, M.C. Evaluating behavior and enclosure use of zoo-housed broad-snouted caiman (Caiman latirostris): A case study in the zoo of Córdoba (Argentina). Herpetol. Conserv. Biol. 2019, 14, 601–614. [Google Scholar]
- Berry, K.J.; Mielke, P.W., Jr. Exact and Monte Carlo resampling procedures for the Wilcoxon-Mann-Whitney and Kruskal-Wallis tests. Percept. Mot. Ski. 2000, 91, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Colegrave, N.J.; Engel, J.; Plowman, A.B. Randomisation tests. In Zoo Research Guidelines: Statistics for Typical Zoo Datasets; Plowman, A.B., Ed.; BIAZA: London, UK, 2006; pp. 7–16. [Google Scholar]
- Leeds, A.; Lukas, K.E. Monitoring the social behavior of a bachelor mandrill (Mandrillus sphinx) dyad participating in touchscreen-mediated cognitive testing. Zoo Biol. 2019, 3, 397–402. [Google Scholar] [CrossRef]
- Plowman, A.B. BIAZA statistics guidelines: Toward a common application of statistical tests for zoo research. Zoo Biol. 2008, 27, 226–233. [Google Scholar] [CrossRef]
- Cronin, K.A.; Ross, S.R. Technical contribution: A cautionary note on the use of behavioural diversity (H-index) in animal welfare science. Anim. Welf. 2019, 28, 157–164. [Google Scholar] [CrossRef]
- Hall, K.; Bryant, J.; Staley, M.; Whitham, J.C.; Miller, L.J. Behavioural diversity as a potential welfare indicator for professionally managed chimpanzees (Pan troglodytes): Exploing variations in calculating diversity using species-specific behaviours. Anim. Welf. 2021, 30, 381–392. [Google Scholar] [CrossRef]
- Brereton, J.E.; Fernandez, E.J. Which index should I use? A comparison of indices for enclosure use studies. Anim. Behav. Cogn. 2022, 9, 119–132. [Google Scholar] [CrossRef]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing reptile welfare using behavioural criteria. Practice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Hoehfurtner, T.; Wilkinson, A.; Nagabaskaran, G.; Burman, O.H.P. Does the provision of environmental enrichment affect the behaviour and welfare of captive snakes? Appl. Anim. Behav. Sci. 2021, 239, 105324. [Google Scholar] [CrossRef]
- Jones, M.; Gartland, K.N.; Fuller, G. Effects of visitor presence and crowd size on zoo-housed red kangaroos (Macropus rufus) During and After a COVID-19 closure. Anim. Behav. Cogn. 2021, 8, 521–537. [Google Scholar] [CrossRef]
- Mitchell, G.; Herring, F.; Obradovich, S.; Tromborg, C.; Dowd, B.; Neville, L.E.; Field, L. Effect of visitors and cage changes on the behaviors of mangabeys. Zoo Biol. 1991, 10, 417–423. [Google Scholar] [CrossRef]
- Kuhar, C.W. Group differences in captive gorillas’ reacion to large crowds. Appl. Anim. Behav. Sci. 2008, 110, 377–385. [Google Scholar] [CrossRef]
- Queiroz, M.B.; Young, R.J. The different physical and behavioural characteristics of zoo mammals that influence their response to visitors. Animals 2018, 8, 139. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Habitat | Activity Cycle | Diet | Stratum Use |
|---|---|---|---|---|
| Catalina Island Rattlesnake (Crotalus catalinensis) | Open [4] | Nocturnal [7] | Carnivore [4] | Terrestrial [4] |
| European Glass Lizard (Pseudopus apodus) | Closed [8] | Diurnal [8] | Carnivore [8] | Terrestrial [8] (semi-fossorial) |
| Beaded Lizard (Heloderma horridum) | Closed [9] | Diurnal [9] | Carnivore [9] | Terrestrial [9] (arboreal) |
| Sonoran Spiny-tailed Iguana (Ctenosaura macrolopha) | Open [5] | Diurnal [30] | Omnivore [12] | Terrestrial [5] (arboreal) |
| Arrau Turtle (Podocnemis expansa) | Closed [13] | Diurnal [31] | Omnivore [13] | Aquatic [13] |
| Dwarf Caiman (Paleosuchus palpebrosus) | Closed [6] | Nocturnal [32] | Carnivore [11] | Aquatic [6] |
| Enclosure | Species | Inhabitants Sex | Ages at Start of Study | Area of Enclosure | Visual of the Enclosure |
|---|---|---|---|---|---|
| A | Catalina Island Rattlesnake | Male Female | 6 years 4 years | 2.3 m2 |  |
| B | European Glass Lizard | Male Female Female Female | 22 years 22 years 22 years 25 years | 2.2 m2 |  |
| C | Beaded Lizard | Male Female Unknown | 26 years 9 years 25 years | 4.6 m2 |  |
| Sonoran Spiny-tailed Iguana | Male | 10 years | |||
| D | Arrau Turtle | Male Female | 39 years 39 years | 17.3 m2 |  |
| Dwarf Caiman | Male Female | 34 years 28 years |
| Time Period | Enclosures | Condition of the Zoo | Approximate Number of Hours Observed | Total Number of Days Analyzed | Comment |
|---|---|---|---|---|---|
| 20 April 2020 to 14 May 2020 | A, B, C | Closed | 9 h | A—18 B—18 C—18 | |
| 12 May 2020 to 5 June 2020 | D | Closed | 8 h | D—16 | An enclosure modification on 11 May 2020, caused a shift in the observational period. |
| 15 June 2020 to 9 July 2020 | All Enclosures | Open | 9 h | A—18 B—17 C—18 D—18 |
| Behavior | Definition |
|---|---|
| Startle/Freeze * | Sudden cessation of movement accompanied by muscle tension while remaining frozen in place |
| Interact with Transparent Boundaries (ITB) * | Repetitive movement directed along glass boundaries of the enclosure; must complete the same circuit at least three times to count as ITB |
| Social * | Any interaction involving two or more animals |
| Move | Whole-body movement that causes a change in location, either via walking, undulating, swimming, or other means |
| Eat | Consumption of food or water |
| Investigate | Movement of the head and neck region to touch or nearly touch features in the immediate surroundings; includes tongue-flicking for snakes and lizards |
| Inactive | The individual is stationary |
| Other | Any behavior not previously described that involves whole-body movement |
| Not Visible | The individual or the individual’s behavior cannot be seen |
| Exposure | Definition |
| Hiding | More than half of the body is concealed from the observer under substrate or furnishings |
| Exposed | Less than half of the body is concealed from the observer under substrate or furnishings |
| Species | Behavior | Percent of Time | Statistic (S) | Z-Score | Pr > |S-Mean| | |||
|---|---|---|---|---|---|---|---|---|
| (Mean ± SE) | Median (Range) | |||||||
| Zoo Closed | Zoo Open | Zoo Closed | Zoo Open | |||||
| Catalina Island Rattlesnake | Visibility | 99.61 ± 0.00 | 99.92 ± 0.08 | 100 (93.0–100) | 100 (97.0–100) | 1295.00 | −0.62 | 0.49 |
| Social ** | 0.37 ± 0.25 | 2.41 ± 0.70 | 0.00 (0.00–3.33) | 1.67 (0.00–10.00) | 1050.00 | −3.68 | <0.01 | |
| Investigate * | 0.09 ± 0.09 | 0.93 ± 0.19 | 0.00 (0.00–3.33) | 0.00 (0.00–10.00) | 1187.50 | −2.48 | 0.03 | |
| Inactive ** | 99.17 ± 0.46 | 95.36 ± 1.12 | 100 (90.00–100) | 96.70 (63.3–100) | 1640.00 | 4.20 | <0.01 | |
| Exposed | 38.70 ± 22.78 | 40.09 ± 11.94 | 10.00 (0.00–100) | 50.00 (0–100) | 1287.50 | −0.30 | 0.77 | |
| European Glass Lizard | Visibility ** | 81.81 ± 3.49 | 57.91 ± 4.03 | 85.00 (62.5–100) | 51.72 (28.33–90.83) | 199.50 | −3.52 | <0.01 |
| Social | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 (0.00) | 0.00 (0.00) | 306.00 | 0.00 | 1.00 | |
| Investigate | 1.67 ± 0.40 | 2.88 ± 1.40 | 1.09 (0–0.06) | 0.00 (0–23.53) | 284.00 | −0.75 | 0.46 | |
| Inactive | 92.73 ± 3.47 | 89.23 ± 1.24 | 94.67 (81.9–100) | 94.29 (52.94–100) | 309.00 | 0.10 | 0.93 | |
| Exposed ** | 34.40 ± 3.57 | 13.09 ± 2.67 | 37.50 (9.17–57.50) | 12.71 (0–42.98) | 194.00 | −3.70 | <0.01 | |
| Beaded Lizard | Visibility | 98.77 ± 1.23 | 100.00 ± 0.00 | 100 (50.00–100) | 100 (100) | 2889.00 | −1.42 | 0.49 |
| Social * | 0.50 ± 0.25 | 2.72 ± 1.09 | 0.00 (0.00–6.90) | 0.00 (0.00–33.33) | 2700.50 | −2.35 | 0.02 | |
| Investigate | 3.47 ± 0.61 | 5.15 ± 3.33 | 0.00 (0.00–46.67) | 0.00 (0.00–48.28) | 2833.50 | −0.80 | 0.43 | |
| Inactive | 93.74 ± 0.50 | 88.97 ± 4.20 | 100 (50.00–100) | 100 (34.48–100) | 3116.00 | 1.20 | 0.23 | |
| Exposed | 87.84 ± 6.12 | 98.39 ± 0.50 | 100 (0.00–100) | 100 (83.33–100) | 2945.50 | 0.02 | 0.99 | |
| Sonoran Spiny-tailed Iguana | Visibility | 100.00 ± 0.00 | 100.00 ± 0.00 | 100 (100) | 100 (100) | 333.00 | 0.00 | 1.00 |
| Social | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 (0.00) | 0.00 (0.00) | 333.00 | 0.00 | 1.00 | |
| Investigate | 0.19 ± 0.19 | 0.94 ± 0.46 | 0.00 (0.00–3.44) | 0.00 (0.00–6.90) | 307.00 | −1.37 | 0.28 | |
| Inactive | 97.38 ± 1.15 | 96.62 ± 1.31 | 100 (82.76–100) | 100 (82.76–100) | 338.00 | 0.19 | 0.85 | |
| Exposed ** | 99.81 ± 0.19 | 95.35 ± 2.72 | 100 (96.67–100) | 98.33 (50.00–100) | 407.00 | 2.98 | <0.01 | |
| Arrau Turtle | Visibility | 99.69 ± 0.10 | 99.72 ± 0.09 | 100 (96.67–100) | 100 (96.67–100) | 1098.00 | −0.15 | 1.00 |
| Social | 1.05 ± 0.003 | 1.95 ± 0.09 | 0.00 (0.00–3.45) | 0.00 (0.00–6.67) | 993.50 | −1.57 | 0.12 | |
| Investigate | 7.21 ± 3.04 | 9.91 ± 3.42 | 3.33 (0.00–30.00) | 6.67 (0.00–43.33) | 1056.50 | −0.59 | 0.56 | |
| Inactive ** | 5.97 ± 3.70 | 39.89 ± 1.60 | 3.33 (0.00–20.00 | 40.00 (0.00–100) | 660.00 | −5.48 | <0.01 | |
| Exposed | 99.79 ± 0.00 | 99.54 ± 0.09 | 100 (96.67–100) | 100 (96.67–100) | 1148.00 | 1.03 | 0.43 | |
| Dwarf Caiman | Visibility | 99.79 ± 0.00 | 99.81 ± 0.19 | 100 (96.67–100) | 100 (96.67–100) | 1085.00 | −0.09 | 1.00 |
| Social | 0.00 ± 0.00 | 0.09 ± 0.09 | 0.00 (0.00) | 0.00 (0.00–3.33) | 1072.00 | −0.96 | 1.00 | |
| Investigate | 0.21 ± 0.004 | 0.10 ± 0.10 | 0.00 (0.00–3.45) | 0.00 (0.00–3.33) | 1107.50 | 0.68 | 0.48 | |
| Inactive ** | 98.12 ± 0.83 | 99.81 ± 0.01 | 100 (86.67–100) | 100 (96.67–100) | 940.00 | −2.78 | <0.01 | |
| Exposed | 99.27 ± 0.31 | 96.59 ± 2.67 | 100 (83.33–100) | 100 (0.00–100) | 1116.00 | 0.62 | 0.60 | |
| Enclosure | Daily Observation Rate of Visitor-Glass Interaction during Zoo Open | Species | Visibility | Exposed | |
|---|---|---|---|---|---|
| Mean ± SE | Median & Range | ||||
| A | 7.51 ± 1.15 | 7.50 (0.00–16.00) | Catalina Island Rattlesnake | r = 0.07 p = 0.57 n = 72 | r = −0.03 p = 0.83 n = 72 |
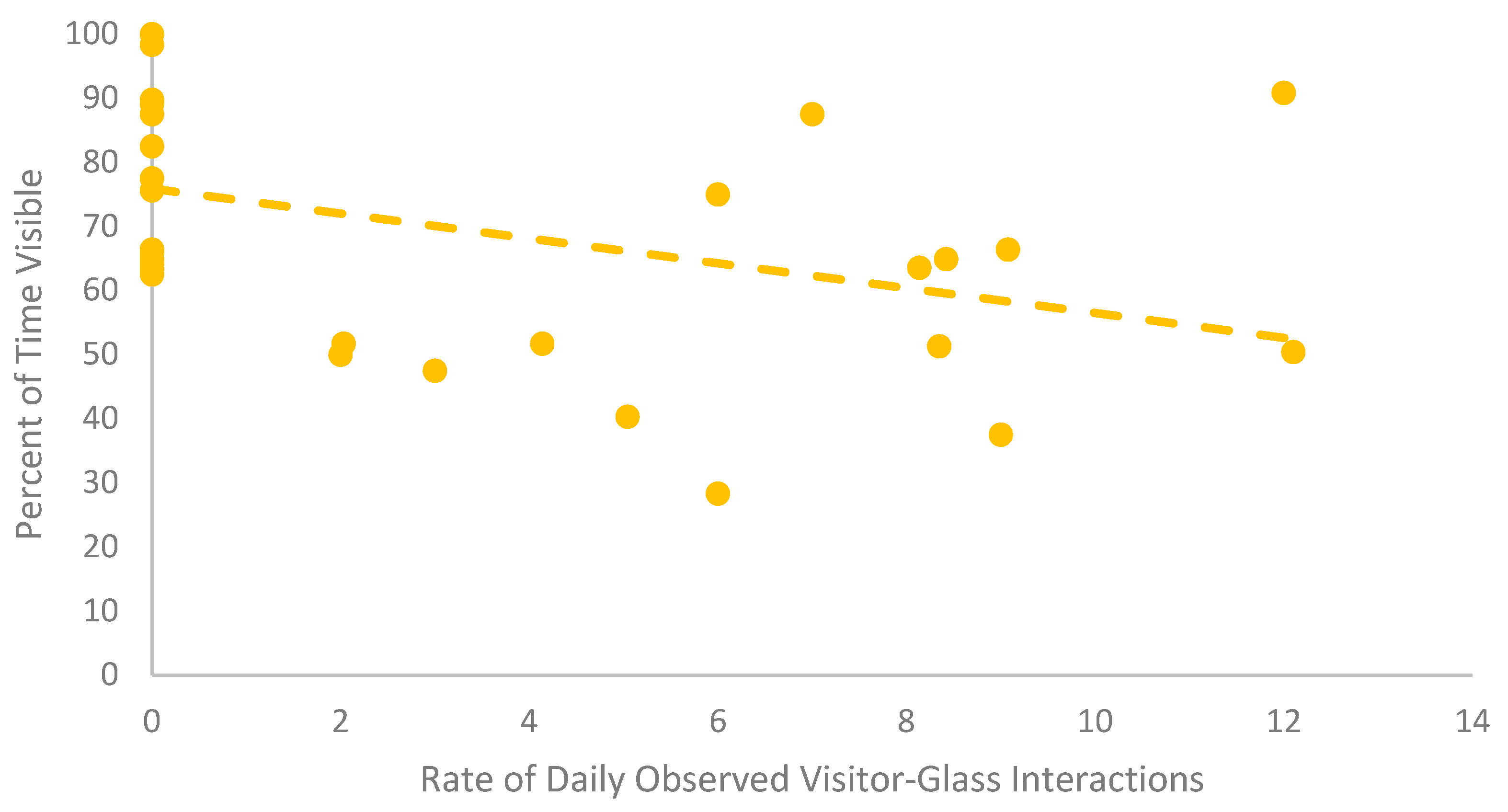
| B | 6.02 ± 0.91 | 6.00 (0.00–12.10) | European Glass Lizard | r = 0.46 p = <0.01 ** n = 35 | r = −0.55 p = <0.01 ** n = 35 |
| C | 7.56 ± 1.08 | 6.50 (0.00–17.00) | Beaded Lizard | r = 0.12 p = 0.23 n = 108 | N/A |
| Sonoran Spiny-tailed Iguana | N/A | N/A | |||
| D | 0.06 ± 0.06 | 0.00 (0.00–1.00) | Arrau Turtle | N/A | N/A |
| Dwarf Caiman | N/A | N/A | |||
| Species | Shannon-Wiener Index | Statistic (S) | Z-Score | Pr > |S-Mean| | |||
|---|---|---|---|---|---|---|---|
| Mean ± SE | Median (Range) | ||||||
| Zoo Closed | Zoo Open | Zoo Closed | Zoo Open | ||||
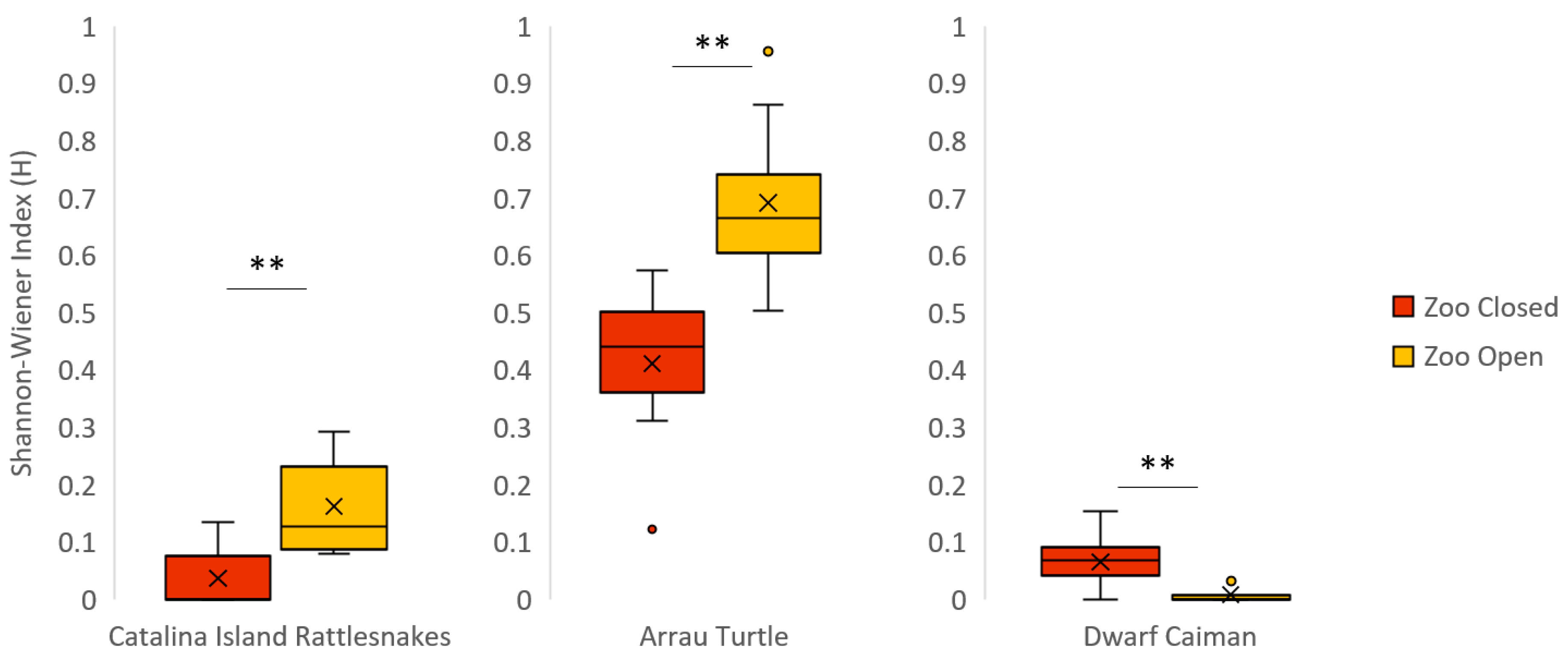
| Catalina Island Rattlesnake ** | 0.04 ± 0.02 | 0.16 ± 0.03 | 0.00 (0.00–0.13) | 0.13 (0.08–0.29) | 43.00 | −2.67 | <0.01 |
| European Glass Lizard | 0.23 ± 0.02 | 0.25 ± 0.05 | 0.22 (0.21–0.28) | 0.27 (0.13–0.34) | 18.00 | 0.00 | 1.00 |
| Beaded Lizard | 0.21 ± 0.01 | 0.30 ± 0.06 | 0.19 (0.00–0.37) | 0.31 (0.00–0.59) | 134.00 | −0.83 | 0.37 |
| Sonoran Spiny-tailed Iguana | 0.11 ± 0.04 | 0.13 ± 0.03 | 0.09 (0.03–0.23) | 0.11 (0.07–0.22) | 16.00 | −0.58 | 0.69 |
| Arrau Turtle ** | 0.41 ± 0.06 | 0.69 ± 0.01 | 0.44 (0.12–0.58) | 0.67 (0.50–0.96) | 39.00 | −3.05 | <0.01 |
| Dwarf Caiman ** | 0.07 ± 0.03 | 0.01 ± 0.01 | 0.07 (0.00–0.15) | 0.00 (0.00–0.04) | 90.00 | 2.47 | 0.01 |
| Species | Dickens’ Spread of Participation Index | Statistic (S) | Z-Score | Pr > |S-Mean| | |||
|---|---|---|---|---|---|---|---|
| Mean ± SE | Median (Range) | ||||||
| Zoo Closed | Zoo Open | Zoo Closed | Zoo Open | ||||
| Catalina Island Rattlesnake | 0.91 ± 0.02 | 0.83 ± 0.01 | 0.92 (0.82–1.00) | 0.83 (0.69–0.92) | 83.00 | 1.58 | 0.12 |
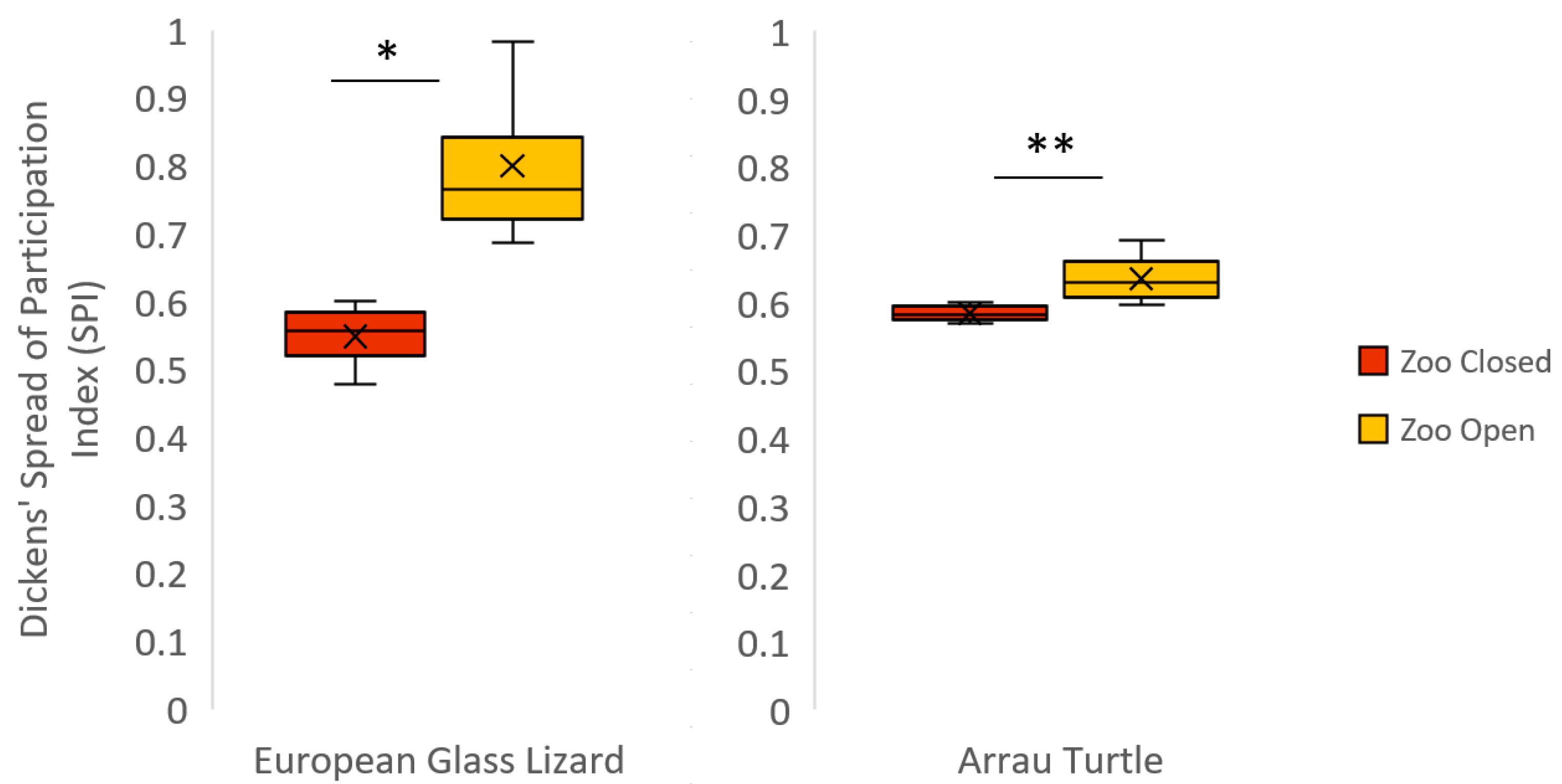
| European Glass Lizard * | 0.55 ± 0.03 | 0.80 ± 0.06 | 0.56 (0.48–0.60) | 0.77 (0.69–0.98) | 10.00 | −2.31 | 0.03 |
| Beaded Lizard | 0.88 ± 0.01 | 0.91 ± 0.01 | 0.88 (0.79–0.94) | 0.93 (0.78–0.97) | 121.00 | −1.68 | 0.10 |
| Sonoran Spiny-tailed Iguana | 0.86 ± 0.02 | 0.80 ± 0.03 | 0.87 (0.79–0.89) | 0.80 (0.73–0.86) | 23.00 | 1.44 | 0.20 |
| Arrau Turtle ** | 0.58 ± 0.005 | 0.63 ± 0.02 | 0.58 (0.57–0.60) | 0.63 (0.60–0.69) | 39.00 | −3.05 | <0.01 |
| Dwarf Caiman | 0.81 ± 0.03 | 0.88 ± 0.02 | 0.78 (0.67–0.97) | 0.88 (0.83–0.94) | 53.00 | −1.58 | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamilton, J.; Gartland, K.N.; Jones, M.; Fuller, G. Behavioral Assessment of Six Reptile Species during a Temporary Zoo Closure and Reopening. Animals 2022, 12, 1034. https://doi.org/10.3390/ani12081034
Hamilton J, Gartland KN, Jones M, Fuller G. Behavioral Assessment of Six Reptile Species during a Temporary Zoo Closure and Reopening. Animals. 2022; 12(8):1034. https://doi.org/10.3390/ani12081034
Chicago/Turabian StyleHamilton, Jennifer, Kylen N. Gartland, Megan Jones, and Grace Fuller. 2022. "Behavioral Assessment of Six Reptile Species during a Temporary Zoo Closure and Reopening" Animals 12, no. 8: 1034. https://doi.org/10.3390/ani12081034
APA StyleHamilton, J., Gartland, K. N., Jones, M., & Fuller, G. (2022). Behavioral Assessment of Six Reptile Species during a Temporary Zoo Closure and Reopening. Animals, 12(8), 1034. https://doi.org/10.3390/ani12081034