
Disease Ecology of a Low-Virulence Mycoplasma ovipneumoniae Strain in a Free-Ranging Desert Bighorn Sheep Population


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
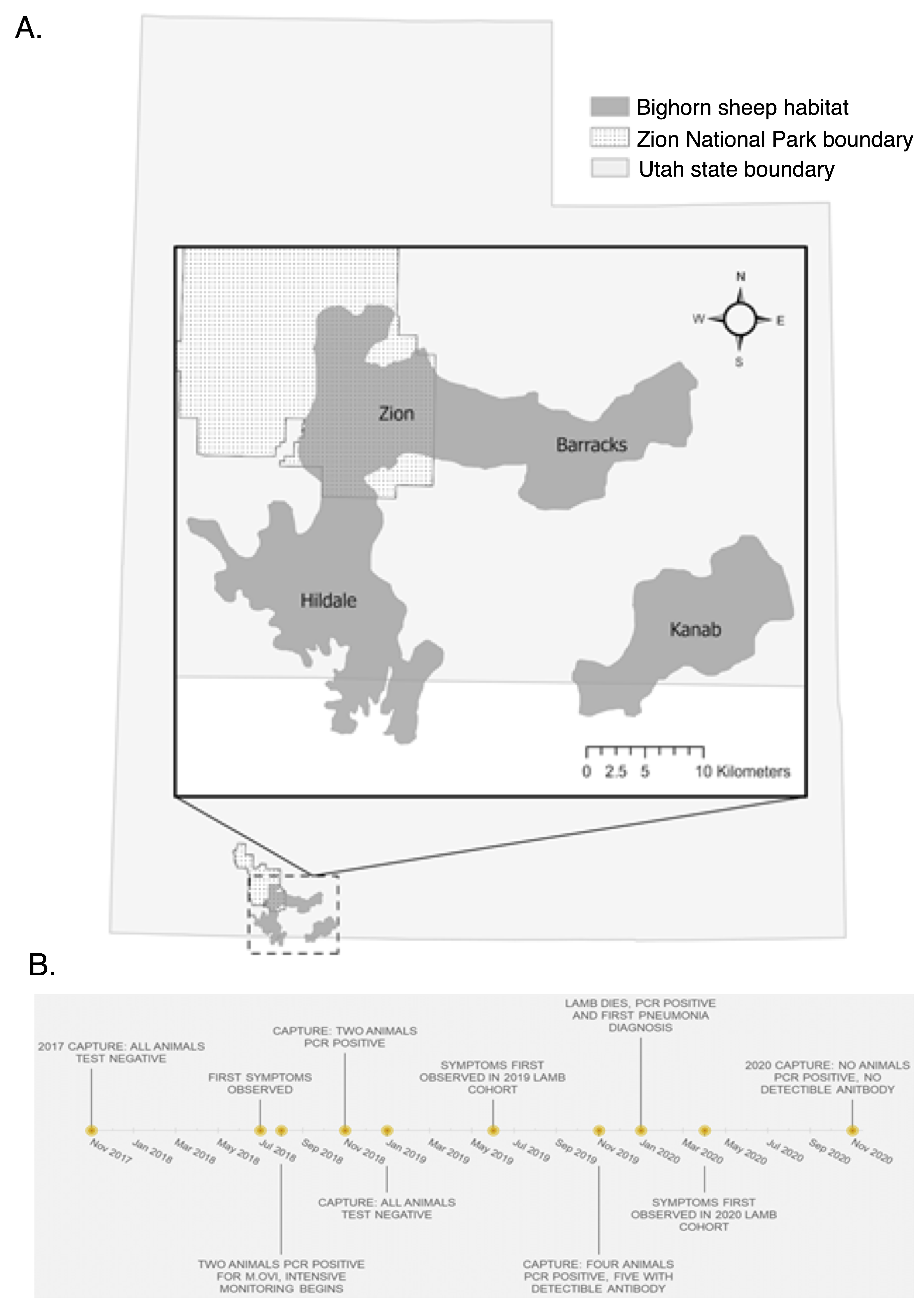
2.1. Study Area
2.2. Data Collection
2.2.1. Captures
2.2.2. Demographic Monitoring
2.2.3. Diagnostic Testing for Serology and Pathogen Load
2.2.4. Symptom Emergence and Intensity
2.3. Data Analysis
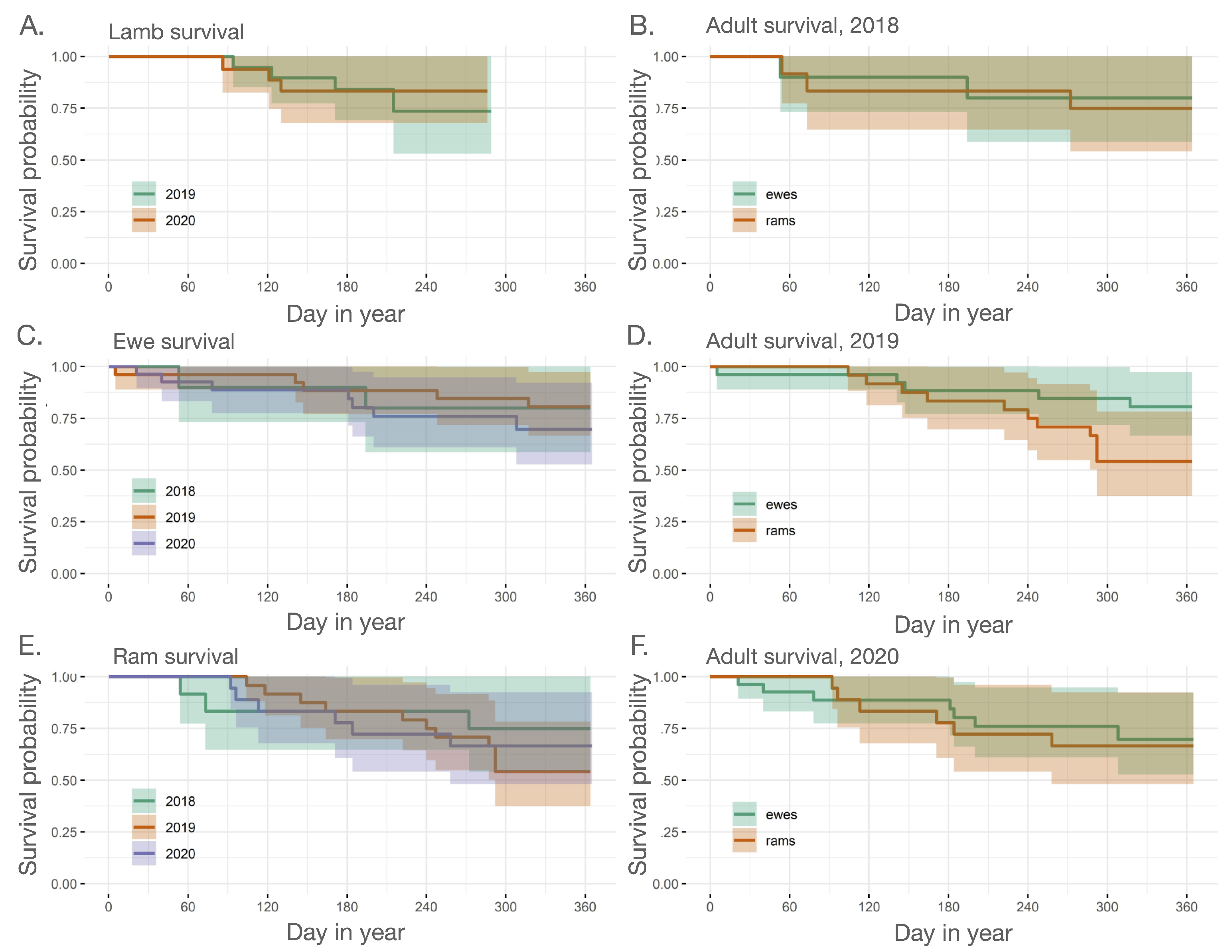
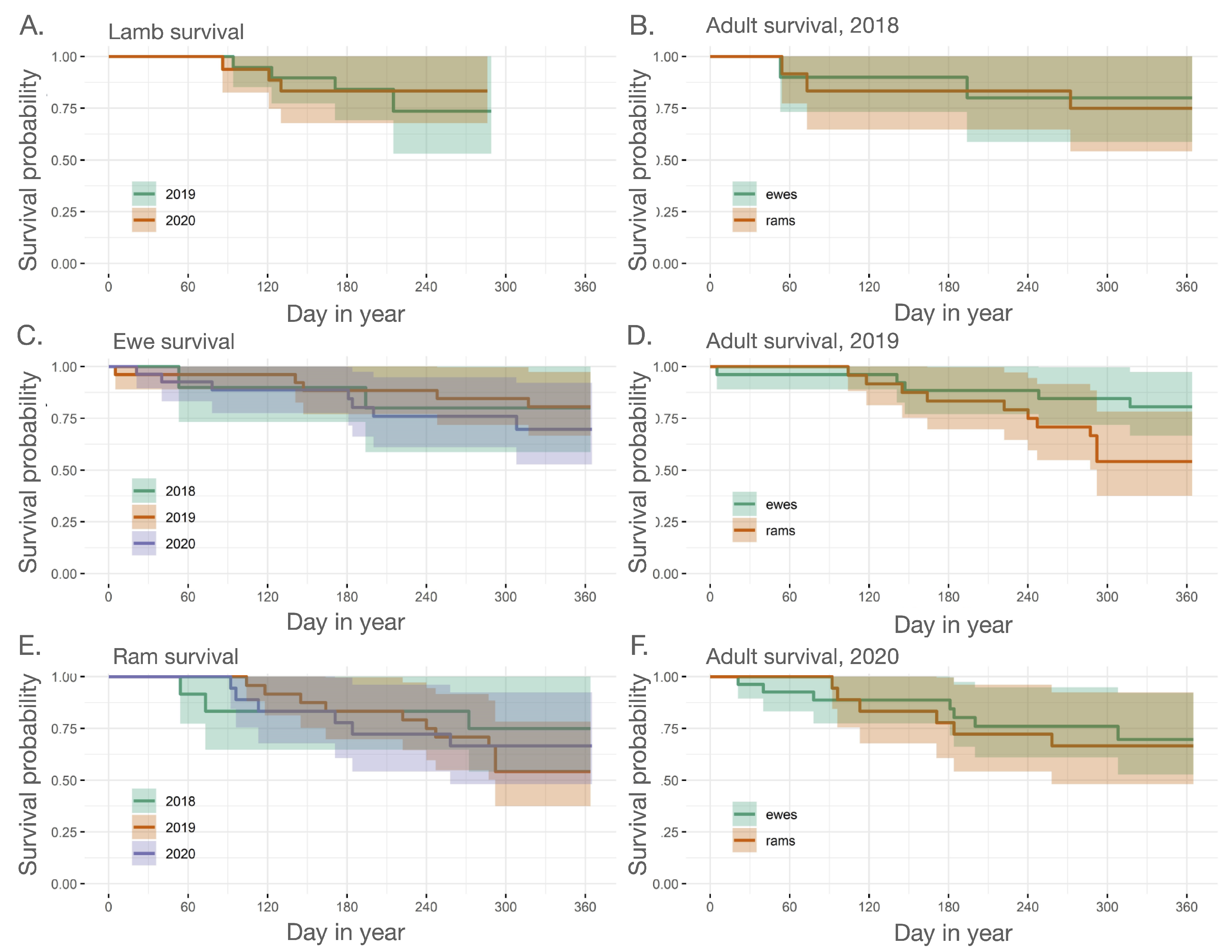
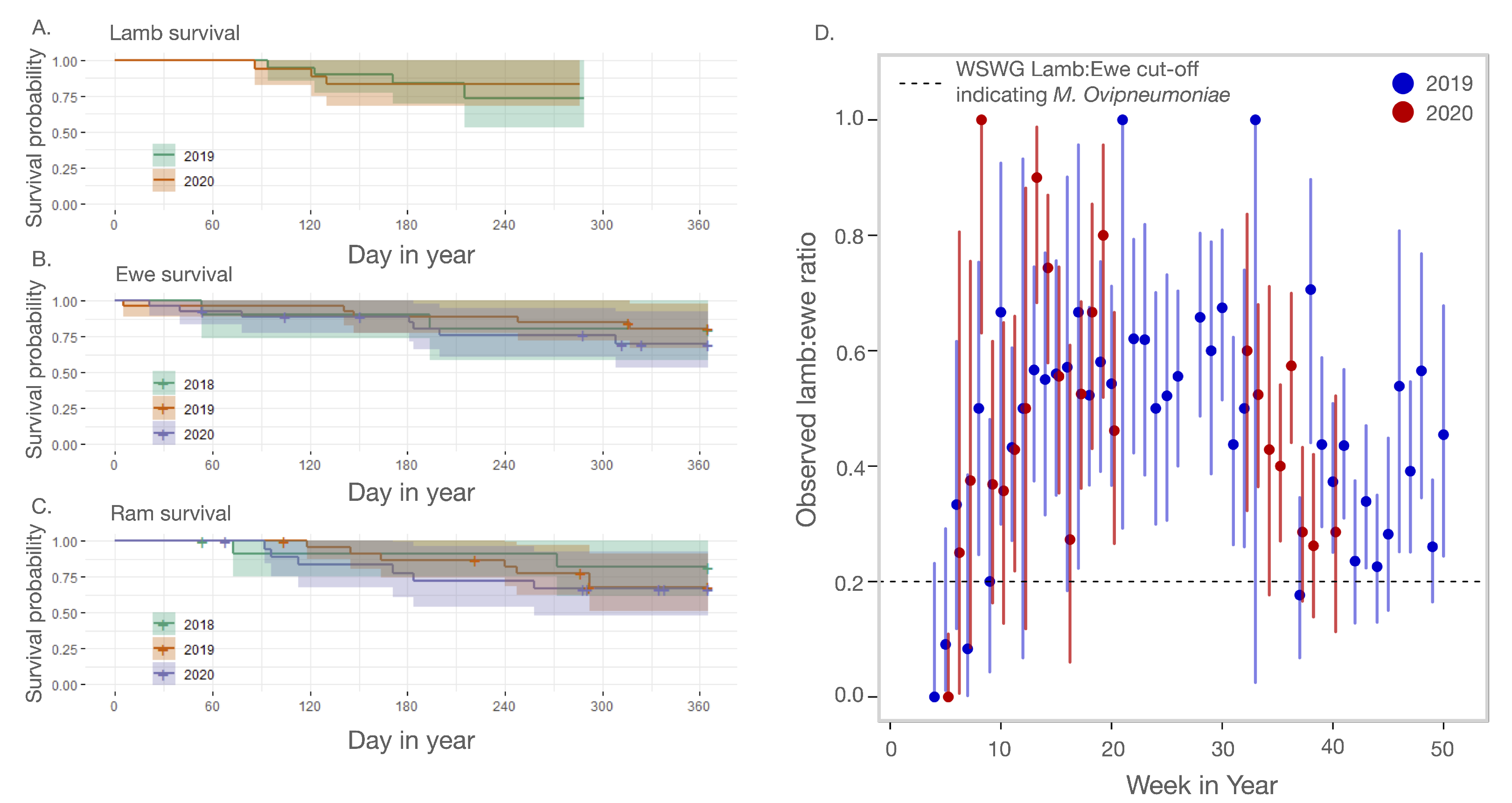
2.3.1. Adult and Juvenile Survival
2.3.2. M. ovipneumoniae Severity and Spatial Distribution of Disease
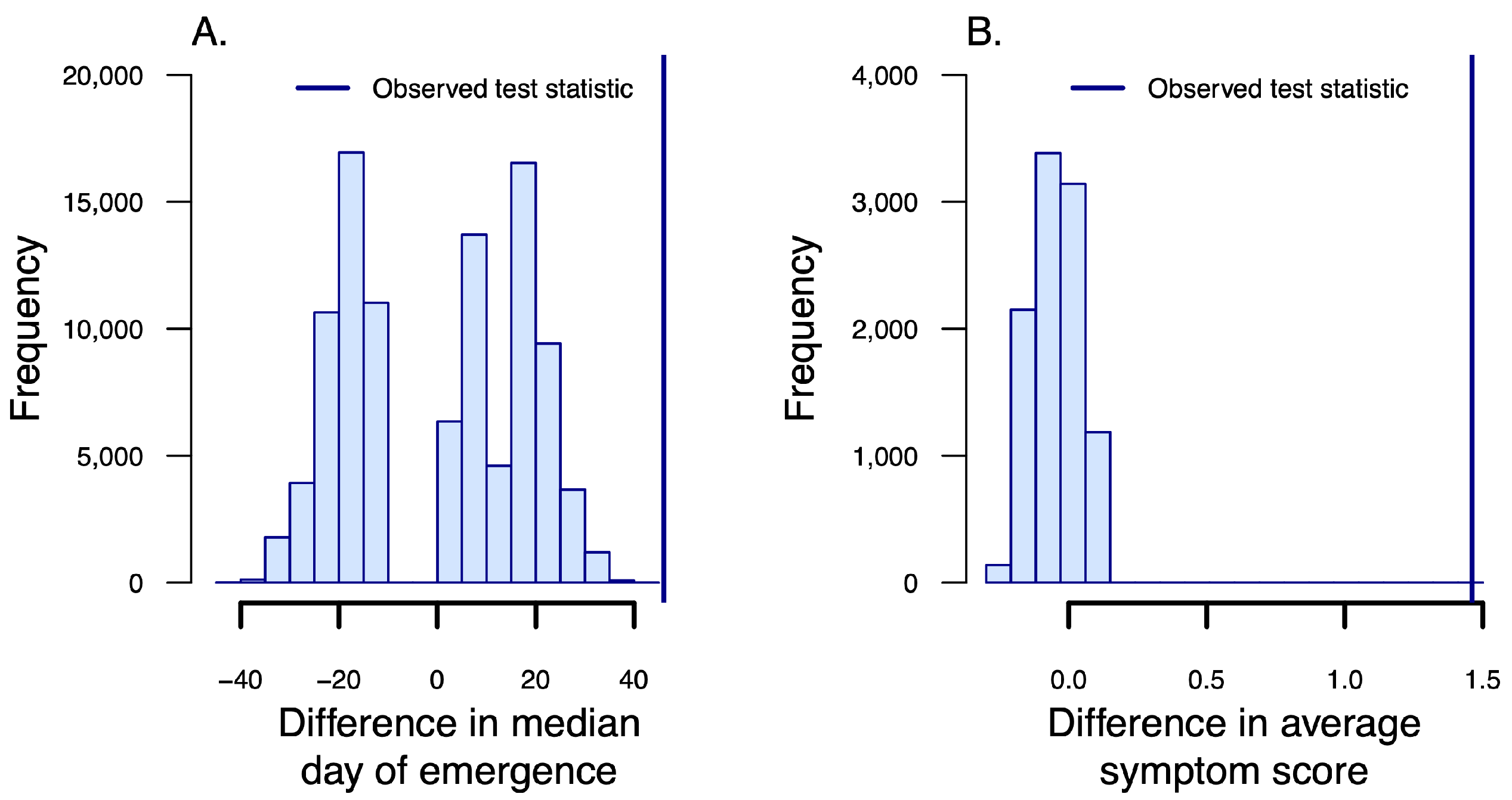
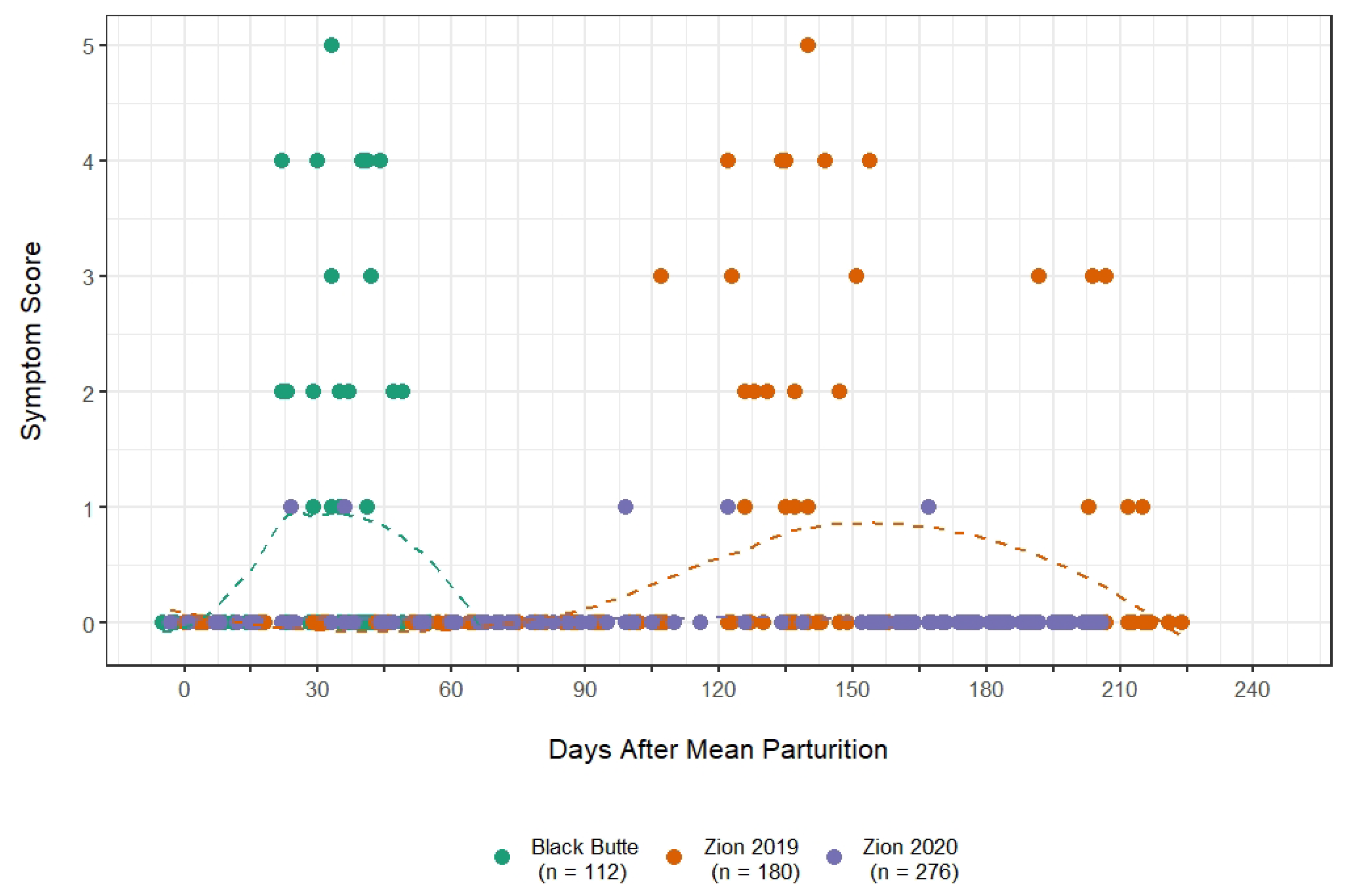
2.3.3. Symptom Emergence and Scoring
2.3.4. Comparison of Serology and Pathogen Load with More Severe Disease Events
2.3.5. Social Mixing Dynamics
3. Results
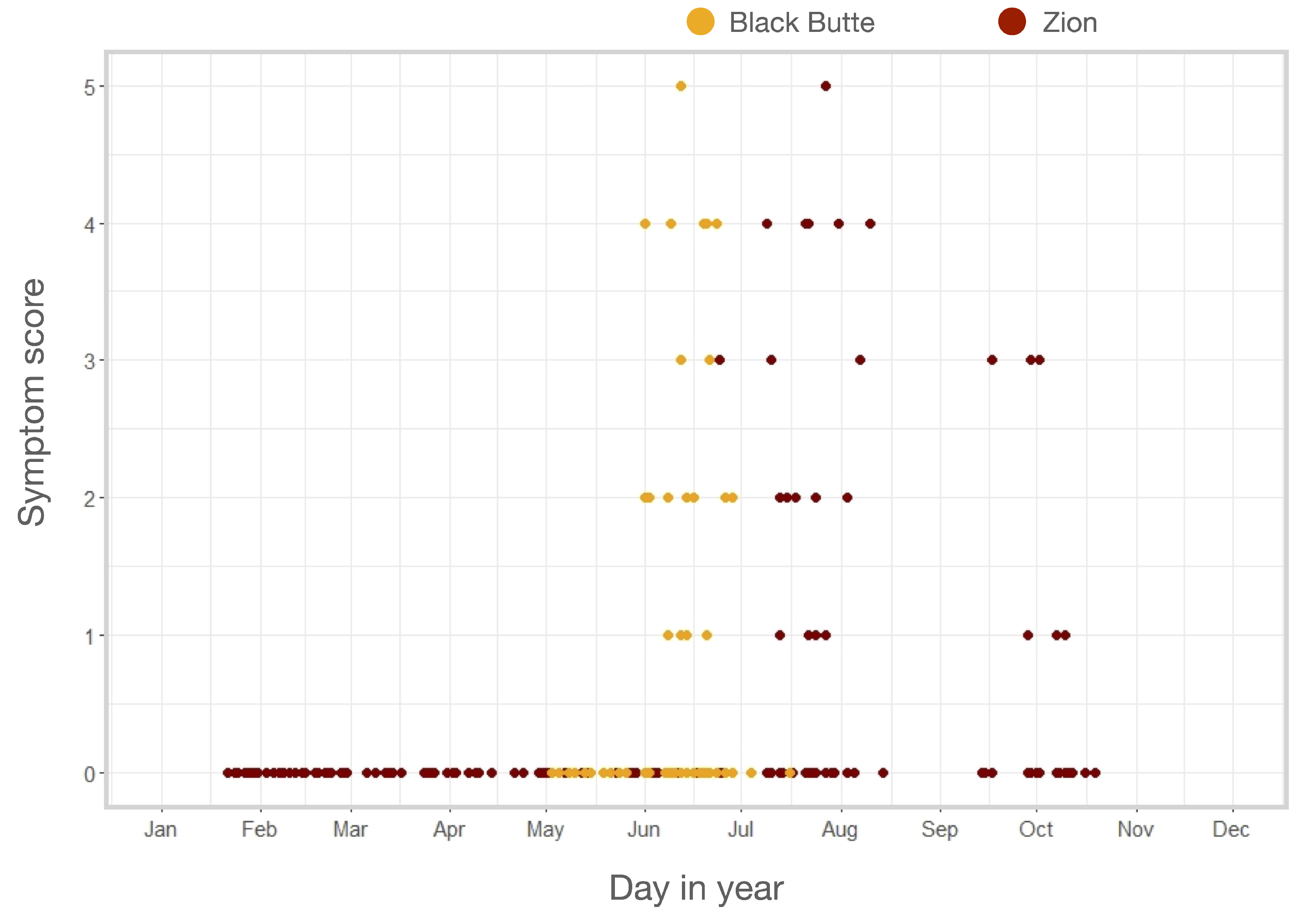
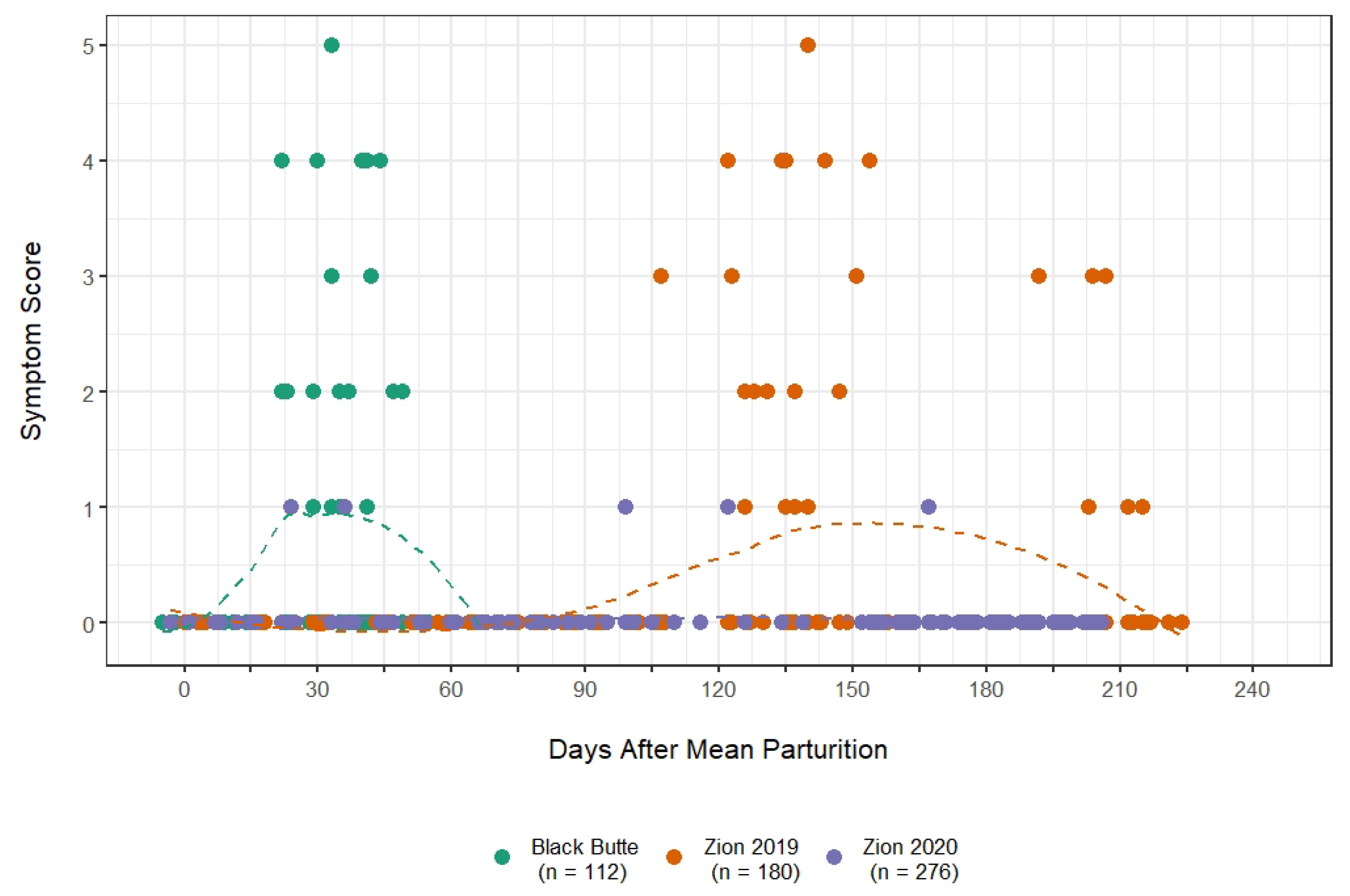
3.1. Outbreak Severity and Duration
3.2. Symptom Emergence and Intensity
3.3. Serology and Pathogen Load
3.4. Social Mixing Dynamics
4. Discussion
4.1. Comparison of the Epidemiology with Other M. ovipneumoniae Disease Events in Bighorns
4.2. Mechanisms That Could Have Produced the Reduced Disease Burden
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Subunit | Sex | Age | Date Notified | Cause |
|---|---|---|---|---|---|
| ZNP | F | adult | 6 January 2019 | Avalanche | |
| ZNP | M | adult | 29 April 2019 | Fall | |
| Barracks | F | adult | 22 May 2019 | Predation | |
| ZNP | M | adult | 26 May 2019 | Predation | |
| Barracks | F | adult | 28 May 2019 | Predation | |
| ZNP | M | adult | 14 June 2019 | Predation | |
| Barracks | M | adult | 1 July 2019 | Predation | |
| Hildale | M | adult | 29 August 2019 | Unknown | |
| Barracks | M | adult | 5 September 2019 | Trapped | |
| Barracks | F | adult | 6 September 2019 | Predation | |
| ZNP | M | adult | 15 September 2019 | Unknown | |
| Hildale | M | adult | 20 October 2019 | Predation | |
| ZNP | M | adult | 20 October 2019 | Drowning | |
| 19-L3 | ZNP | F | lamb | 9 January 2020 | Uncertain; had pneumonia |
| Kanab | F | adult | 22 January 2020 | Predation | |
| ZNP | F | adult | 10 February 2020 | Predation | |
| Hildale | F | adult | 19 March 2020 | Fall | |
| ZNP | M | adult | 2 April 2020 | Predation | |
| ZNP | M | adult | 6 April 2020 | Predation | |
| ZNP | M | adult | 23 April 2020 | Predation | |
| Hildale | M | adult | 20 June 2020 | Predation | |
| Hildale | F | adult | 30 June 2020 | Predation | |
| Hildale | F | adult | 3 July 2020 | Predation | |
| Hildale | F | adult | 19 July 2020 | Predation | |
| ZNP | M | adult | 4 November 2020 | Starvation | |
| ZNP | F | adult | 24 November 2020 | Predation | |
| ZNP | F | adult | 13 December 2020 | Fall | |
| Barracks | M | adult | 24 December 2020 | Capture myopathy | |
| ZNP | M | adult | 9 January 2021 | Septicemia | |
| Barracks | M | adult | 12 January 2021 | Fall |
| Year | Ewe’s ID | Subunit | Lamb Status | Lamb Fate |
|---|---|---|---|---|
| 2019 | ZNP | confirmed 20 June 2019 | weaned | |
| 2019 | ZNP | confirmed 13 May 2019 | unknown | |
| 2019 | ZNP | confirmed 18 July 2019 | weaned | |
| 2019 | ZNP | confirmed 27 February 2019 | weaned | |
| 2019 | ZNP | confirmed 13 March 2019 | weaned | |
| 2019 | ZNP | confirmed 13 March 2019 | weaned | |
| 2019 | ZNP | confirmed 28 May 2019 | unknown | |
| 2019 | ZNP | confirmed fall 2019 | weaned | |
| 2019 | ZNP | confirmed 11 March 2019 | drowned 03 August 2019 | |
| 2019 | ZNP | confirmed 28 March 2019 | weaned | |
| 2019 | ZNP | confirmed 28 March 2019 | weaned | |
| 2019 | ZNP | no lamb | - | |
| 2019 | Barracks | confirmed 30 January 2019 | lost by 30 May 2019 | |
| 2019 | Barracks | confirmed 18 March 2019 | weaned | |
| 2019 | Barracks | confirmed 18 March 2019 | lost by 26 May 2019 | |
| 2019 | Kanab | confirmed 21 March 2019 | weaned | |
| 2019 | Kanab | confirmed 29 January 2019 | lost by 04 April 2019 | |
| 2019 | Barracks | confirmed 11 February 2019 | weaned | |
| 2019 | Hildale | confirmed 19 February 2019 | weaned | |
| 2019 | Barracks | confirmed 9 February 2019 | weaned | |
| 2019 | Hildale | confirmed 22 February 2019 | weaned | |
| 2019 | Hildale | no lamb | - | |
| 2020 | Barracks | confirmed 8 May 2020 | weaned | |
| 2020 | Barracks | suspected before 22 April 2020 | lost by 10 May 2020 | |
| 2020 | Hildale | confirmed 21 February 2020 | unknown, ewe died 17 March 2020 | |
| 2020 | Hildale | confirmed 11 April 2020 | unknown, ewe died 19 July 2020 | |
| 2020 | Hildale | confirmed 28 May 2020 | unknown, ewe died 30 June 2020 | |
| 2020 | Kanab | confirmed 6 February 2020 | lost by 31 March 2020 | |
| 2020 | ZNP | confirmed 16 June 2020 | weaned | |
| 2020 | ZNP | confirmed 3 March 2020 | weaned | |
| 2020 | ZNP | confirmed 2 April 2020 | weaned | |
| 2020 | ZNP | confirmed 18 February 2020 | weaned | |
| 2020 | ZNP | confirmed 31 March 2020 | weaned | |
| 2020 | ZNP | confirmed 31 March 2020 | weaned | |
| 2020 | ZNP | confirmed 2 April 2020 | weaned | |
| 2020 | ZNP | confirmed 16 May 2020 | weaned | |
| 2020 | ZNP | confirmed 3 April 2020 | weaned | |
| 2020 | ZNP | confirmed 31 March 2020 | weaned | |
| 2020 | ZNP | confirmed 6 March 2020 | lost by 16 May 2020 | |
| 2020 | ZNP | confirmed 3 March 2020 | weaned | |
| 2020 | ZNP | no lamb | - | |
| 2020 | ZNP | confirmed 19 February 2020 | weaned | |
| 2020 | ZNP | confirmed 15 May 2020 | weaned |
References
- Buechner, H.K. The bighorn sheep in the United States, its past, present, and future. Wildl. Monogr. 1960, 4, 3–174. [Google Scholar]
- Miller, D.S.; Hoberg, E.; Weiser, G.; Aune, K.; Atkinson, M.; Kimberling, C. A review of hypothesized determinants associated with bighorn sheep (Ovis canadensis) die-offs. Vet. Med. Int. 2012, 2012, 796527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besser, T.E.; Cassirer, E.F.; Potter, K.A.; VanderSchalie, J.; Fischer, A.; Knowles, D.P.; Herndon, D.R.; Rurangirwa, F.R.; Weiser, G.C.; Srikumaran, S. Association of Mycoplasma ovipneumoniae infection with population-limiting respiratory disease in free-ranging Rocky Mountain bighorn sheep (Ovis canadensis canadensis). J. Clin. Microbiol. 2008, 46, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dassanayake, R.P.; Shanthalingam, S.; Herndon, C.N.; Subramaniam, R.; Lawrence, P.K.; Bavananthasivam, J.; Cassirer, E.F.; Haldorson, G.J.; Foreyt, W.J.; Rurangirwa, F.R.; et al. Mycoplasma ovipneumoniae can predispose bighorn sheep to fatal Mannheimia haemolytica pneumonia. Vet. Microbiol. 2010, 145, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Besser, T.E.; Highland, M.A.; Baker, K.; Cassirer, E.F.; Anderson, N.J.; Ramsey, J.M.; Mansfield, K.; Bruning, D.L.; Wolff, P.; Smith, J.B.; et al. Causes of pneumonia epizootics among bighorn sheep, western United States, 2008–2010. Emerg. Infect. Dis. 2012, 18, 406. [Google Scholar] [CrossRef]
- Jones, G.; Keir, W.; Gilmour, J. The pathogenicity of Mycoplasma ovipneumoniae and Mycoplasma arginini in ovine and caprine tracheal organ cultures. J. Comp. Pathol. 1985, 95, 477–487. [Google Scholar] [CrossRef]
- Niang, M.; Rosenbusch, R.F.; Andrews, J.J.; Lopez-Virella, J.; Kaeberle, M.L. Occurrence of autoantibodies to cilia in lambs with acoughing syndrome’. Vet. Immunol. Immunopathol. 1998, 64, 191–205. [Google Scholar] [CrossRef]
- Besser, T.E.; Cassirer, E.F.; Highland, M.A.; Wolff, P.; Justice-Allen, A.; Mansfield, K.; Davis, M.A.; Foreyt, W. Bighorn sheep pneumonia: Sorting out the cause of a polymicrobial disease. Prev. Vet. Med. 2013, 108, 85–93. [Google Scholar] [CrossRef]
- Cassirer, E.F.; Sinclair, A. Dynamics of pneumonia in a bighorn sheep metapopulation. J. Wildl. Manag. 2007, 71, 1080–1088. [Google Scholar] [CrossRef]
- Cassirer, E.F.; Plowright, R.K.; Manlove, K.R.; Cross, P.C.; Dobson, A.P.; Potter, K.A.; Hudson, P.J. Spatio-temporal dynamics of pneumonia in bighorn sheep. J. Anim. Ecol. 2013, 82, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Manlove, K.; Cassirer, E.F.; Cross, P.C.; Plowright, R.K.; Hudson, P.J. Disease introduction is associated with a phase transition in bighorn sheep demographics. Ecology 2016, 97, 2593–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besser, T.E.; Cassirer, E.F.; Potter, K.A.; Lahmers, K.; Oaks, J.L.; Shanthalingam, S.; Srikumaran, S.; Foreyt, W.J. Epizootic pneumonia of bighorn sheep following experimental exposure to Mycoplasma ovipneumoniae. PLoS ONE 2014, 9, e110039. [Google Scholar] [CrossRef] [PubMed]
- Manlove, K.; Branan, M.; Baker, K.; Bradway, D.; Cassirer, E.F.; Marshall, K.L.; Miller, R.S.; Sweeney, S.; Cross, P.C.; Besser, T.E. Risk factors and productivity losses associated with Mycoplasma ovipneumoniae infection in United States domestic sheep operations. Prev. Vet. Med. 2019, 168, 30–38. [Google Scholar] [CrossRef]
- Plowright, R.K.; Manlove, K.R.; Besser, T.E.; Páez, D.J.; Andrews, K.R.; Matthews, P.E.; Waits, L.P.; Hudson, P.J.; Cassirer, E.F. Age-specific infectious period shapes dynamics of pneumonia in bighorn sheep. Ecol. Lett. 2017, 20, 1325–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamath, P.L.; Manlove, K.; Cassirer, E.F.; Cross, P.C.; Besser, T.E. Genetic structure of Mycoplasma ovipneumoniae informs pathogen spillover dynamics between domestic and wild Caprinae in the western United States. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Cassirer, E.F.; Manlove, K.R.; Plowright, R.K.; Besser, T.E. Evidence for strain-specific immunity to pneumonia in bighorn sheep. J. Wildl. Manag. 2017, 81, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Cassirer, E.F.; Manlove, K.R.; Almberg, E.S.; Kamath, P.L.; Cox, M.; Wolff, P.; Roug, A.; Shannon, J.; Robinson, R.; Harris, R.B.; et al. Pneumonia in bighorn sheep: Risk and resilience. J. Wildl. Manag. 2018, 82, 32–45. [Google Scholar] [CrossRef]
- Zion National Park Bighorn Sheep Management Environmental Assessment; Technical Report; U.S. Department of Interior: National Park Service: Washington, DC, USA, 2017. Available online: https://parkplanning.nps.gov/document.cfm?parkID=113&projectID=51293&documentID=82480 (accessed on 20 March 2020).
- Krausman, P.R.; Hervert, J.J.; Ordway, L.L. Capturing deer and mountain sheep with a net-gun. Wildl. Soc. Bull. 1985, 13, 71–73. [Google Scholar]
- Ziegler, J.C.; Lahmers, K.K.; Barrington, G.M.; Parish, S.M.; Kilzer, K.; Baker, K.; Besser, T.E. Safety and immunogenicity of a Mycoplasma ovipneumoniae bacterin for domestic sheep (Ovis aries). PLoS ONE 2014, 9, e95698. [Google Scholar] [CrossRef]
- Besser, T.E.; Cassirer, E.F.; Potter, K.A.; Foreyt, W.J. Exposure of bighorn sheep to domestic goats colonized with Mycoplasma ovipneumoniae induces sub-lethal pneumonia. PLoS ONE 2017, 12, e0178707. [Google Scholar] [CrossRef] [Green Version]
- Besser, T.E.; Cassirer, E.F.; Lisk, A.; Nelson, D.; Manlove, K.R.; Cross, P.C.; Hogg, J.T. Natural history of a bighorn sheep pneumonia epizootic: Source of infection, course of disease, and pathogen clearance. Ecol. Evol. 2021, 11, 14366–14382. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.M.; Lumley, T. Package ‘survival’. R. Top. Doc. 2015, 128, 28–33. [Google Scholar]
- Fieberg, J.; DelGiudice, G.D. What time is it? Choice of time origin and scale in extended proportional hazards models. Ecology 2009, 90, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.J.; Schwager, S.J. A comparison of association indices. Anim. Behav. 1987, 35, 1454–1469. [Google Scholar] [CrossRef] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Int. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Pons, P.; Latapy, M. Computing communities in large networks using random walks. J. Graph Algorithms Appl. Citeseer 2006. [Google Scholar] [CrossRef] [Green Version]
- Farine, D.R. Measuring phenotypic assortment in animal social networks: Weighted associations are more robust than binary edges. Anim. Behav. 2014, 89, 141–153. [Google Scholar] [CrossRef]
- Cassirer, E.; Oldenburg, L.; Coggins, V.; Fowler, P.; Rudolph, K.; Hunter, D.; Foreyt, W. Overview and preliminary analysis of Hells Canyon bighorn sheep die-off, 1995–6. In Proceedings of the Tenth Biennial Symposium of the Northern Wild Sheep and Goat Council, Silverthorne, CO, USA, 30 April–3 May 1996. [Google Scholar]
- Sah, P.; Leu, S.T.; Cross, P.C.; Hudson, P.J.; Bansal, S. Unraveling the disease consequences and mechanisms of modular structure in animal social networks. Proc. Natl. Acad. Sci. USA 2017, 114, 4165–4170. [Google Scholar] [CrossRef] [Green Version]
- Manlove, K.R.; Cassirer, E.F.; Cross, P.C.; Plowright, R.K.; Hudson, P.J. Costs and benefits of group living with disease: A case study of pneumonia in bighorn lambs (Ovis canadensis). Proc. R. Soc. B Biol. Sci. 2014, 281, 20142331. [Google Scholar] [CrossRef] [Green Version]
- Borg, N.J.; Mitchell, M.S.; Lukacs, P.M.; Mack, C.M.; Waits, L.P.; Krausman, P.R. Behavioral connectivity among bighorn sheep suggests potential for disease spread. J. Wildl. Manag. 2017, 81, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, L.L.; Diamond, B.; Spraker, T.R.; Sirochman, M.A.; Walsh, D.P.; Machin, C.M.; Bade, D.J.; Miller, M.W. A bighorn sheep die-off in southern Colorado involving a Pasteurellaceae strain that may have originated from syntopic cattle. J. Wildl. Dis. 2010, 46, 1262–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.B.; Jenks, J.A.; Grovenburg, T.W.; Klaver, R.W. Disease and predation: Sorting out causes of a bighorn sheep (Ovis canadensis) decline. PLoS ONE 2014, 9, e88271. [Google Scholar] [CrossRef] [PubMed]
- Justice-Allen, A.E.; Butler, E.; Pebworth, J.; Munig, A.; Wolff, P.; Besser, T.E. Investigation of pneumonia mortalities in a Mycoplasma-positive desert bighorn sheep population and detection of a different strain of Mycoplasma ovipneumoniae. In Proceedings of the Biennial Symposium Northern Wild Sheep Goat Council, Moscow, ID, USA, 9–13 May 2016; Volume 20, pp. 68–72. [Google Scholar]
- Dekelaita, D.J.; Epps, C.W.; Stewart, K.M.; Sedinger, J.S.; Powers, J.G.; Gonzales, B.J.; Abella-Vu, R.K.; Darby, N.W.; Hughson, D.L. Survival of adult female bighorn sheep following a pneumonia epizootic. J. Wildl. Manag. 2020, 84, 1268–1282. [Google Scholar] [CrossRef]
- Shirkey, N.; Roug, A.; Besser, T.; Bleich, V.C.; Darby, N.; Dekelaita, D.; Galloway, N.L.; Gonzales, B.; Hughson, D.; Konde, L.; et al. Previously Unrecognized Exposure of Desert Bighorn Sheep (Ovis canadensis nelsoni) to Mycoplasma ovipneumoniae in the California Mojave Desert. J. Wildl. Dis. 2021, 57, 447–452. [Google Scholar] [CrossRef]
- Stephen, C. Toward a modernized definition of wildlife health. J. Wildl. Dis. 2014, 50, 427–430. [Google Scholar] [CrossRef]
- Butler, C.J.; Edwards, W.H.; Jennings-Gaines, J.E.; Killion, H.J.; Wood, M.E.; McWhirter, D.E.; Paterson, J.T.; Proffitt, K.M.; Almberg, E.S.; White, P.J.; et al. Assessing respiratory pathogen communities in bighorn sheep populations: Sampling realities, challenges, and improvements. PLoS ONE 2017, 12, e0180689. [Google Scholar] [CrossRef] [Green Version]
- Rubin, E.S.; Boyce, W.M.; Bleich, V.C. Reproductive strategies of desert bighorn sheep. J. Mammal. 2000, 81, 769–786. [Google Scholar] [CrossRef]
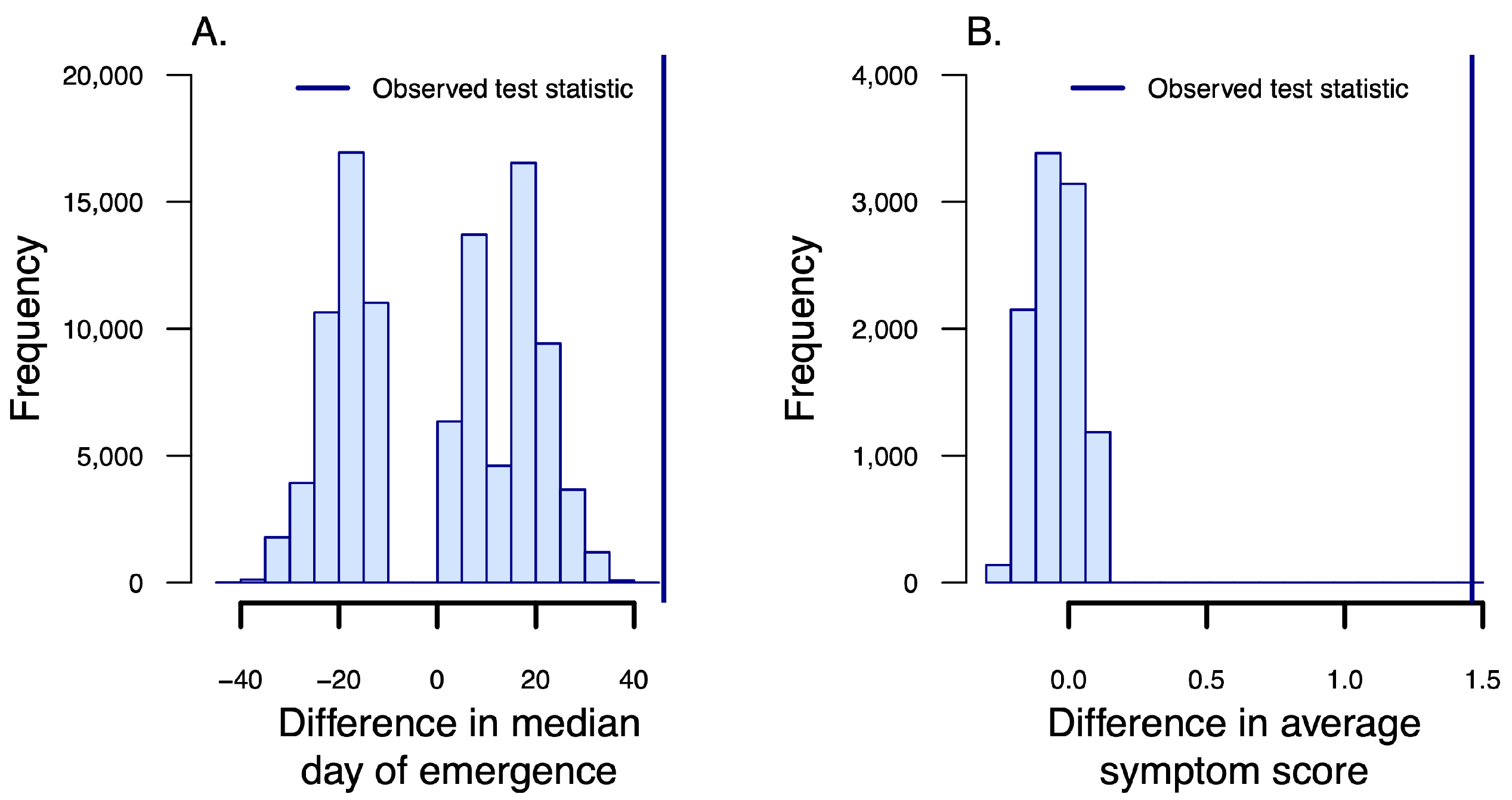
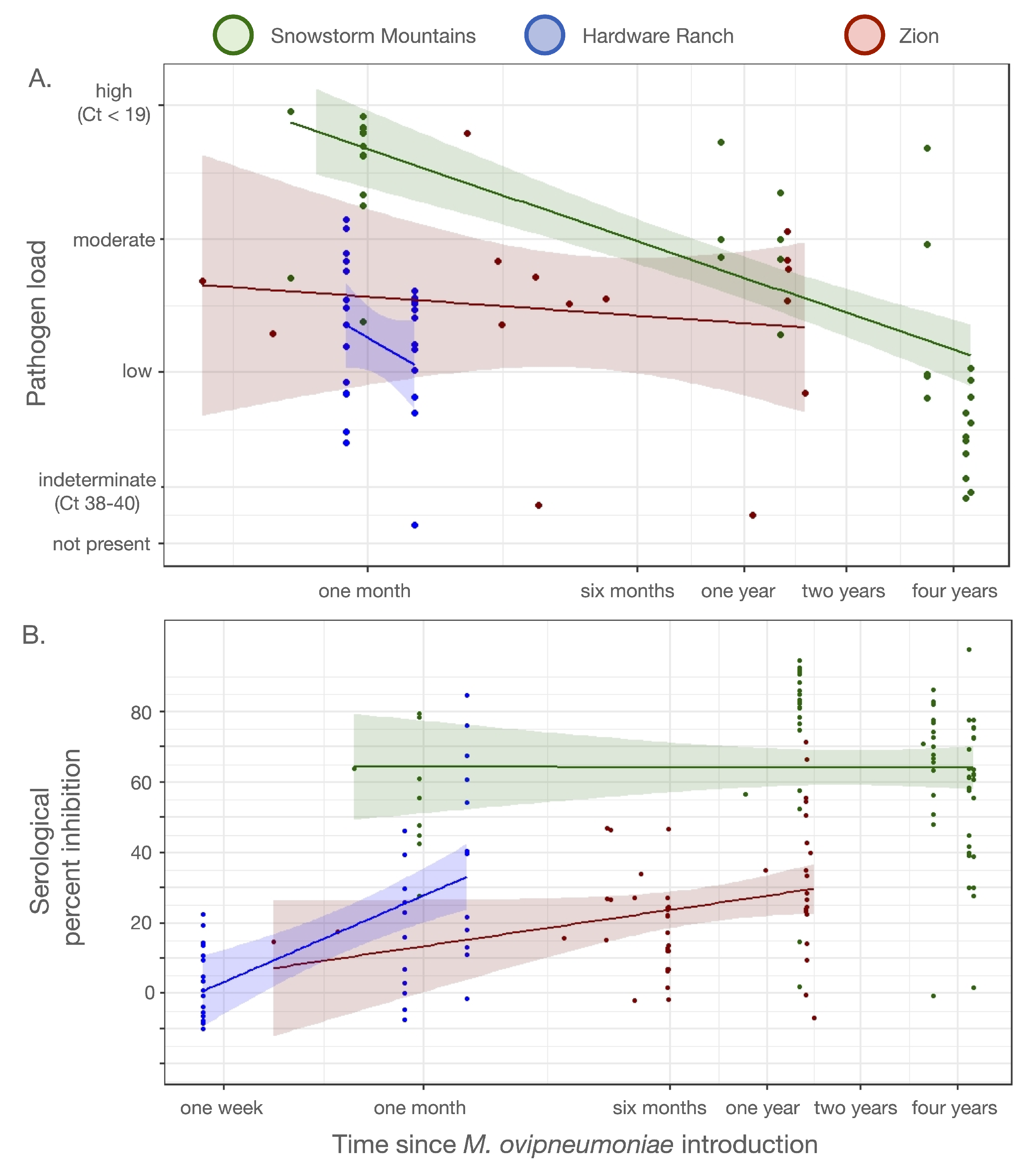
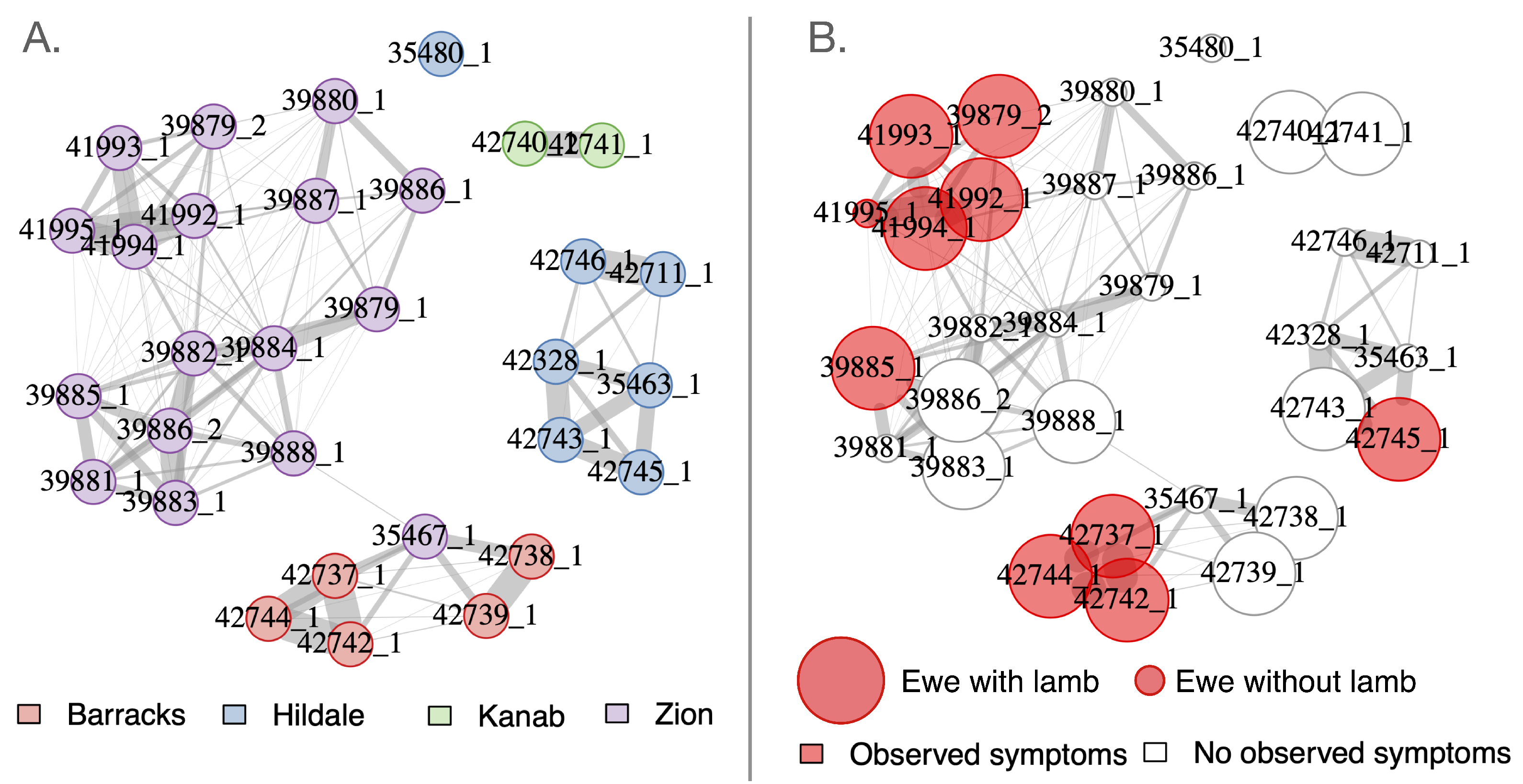
| Subunit & Season | Dates | Total Sampled | Ewes | Rams | Juv. | Collars Deployed | PCR-pos. (Total Tests) | cELISA-pos., indeterm. (Total Tests) | Agency | Method |
|---|---|---|---|---|---|---|---|---|---|---|
| Barracks 2015 | 31 October 2015 | 6 | 6 | 0 | 0 | 6 | 0 (6) | 0, 0 (6) | UDWR | helicopter |
| Hildale 2015 | 30–31 October 2015 | 10 | 10 | 0 | 0 | 10 | 0 (10) | 0, 0 (10) | UDWR | helicopter |
| ZNP 2017 | 15 November 2017 and 4 December 2017 | 23 | 10 | 13 | 0 | 23 | 0 (23) | 0, 0 (23) | NPS & UDWR | helicopter |
| ZNP 2017 | 12–13 December 2017 | 50 * | 41 * | 5 * | 4 | 0 | 0 (50) | 0, 0 (49) | NPS & UDWR | helicopter |
| Barracks 2018–19 | 9 January 2019 | 9 | 5 | 4 | 0 | 9 | 0 (9) | 0, 1 (9) | UDWR | helicopter |
| Hildale 2018–19 | 9 January 2019 | 9 | 5 | 4 | 0 | 9 | 0 (9) | 0, 0 (9) | UDWR | helicopter |
| Kanab 2018–19 | 8 January 2019 | 4 | 2 | 2 | 0 | 4 | 0 (4) | 0, 0 (4) | UDWR | helicopter |
| ZNP 2018–19 | 5 November–1 December 2018 | 11 | 6 | 5 | 0 | 11 | 2 (10) | 0, 2 (10) | NPS & UDWR | ground |
| ZNP 2019–20 | 7 November 2018–30 January 2019 | 21 | 7 | 8 | 6 | 15 | 4 (18) | 5, 1 (18) | NPS & UDWR | ground |
| Barracks 2020 | 13 December 2020 | 6 | 4 | 2 | 0 | 6 | 0 (6) | 2,0 (6) | UDWR | helicopter |
| Hildale 2020 | 11 December 2020 | 7 | 5 | 2 | 0 | 7 | 0 (7) | 0, 1 (7) | UDWR | helicopter |
| Kanab 2020 | 13 December 2020 | 2 | 1 | 1 | 0 | 2 | 0 (2) | 0, 0 (2) | NPS & UDWR | group & helicopter |
| ZNP 2020 | 6 November–12 December 2020 | 20 | 17 | 3 | 0 | 19 | 0 (20) | 1, 0 (18) | NPS & UDWR | ground & helicopter |
| Level | Communities | Assortativity | Assort. p-Value | Betweenness | Between. p-Value | Modularity |
|---|---|---|---|---|---|---|
| Whole population all years | 16 | 0.16 | 0.007 | 666 | 0.1896 | 0.16 |
| Ewes all years | 8 | 0.21 | 0.003 | - | - | 0.19 |
| ZNP all years | 16 | 0.1 | 0.005 | 374 | 0.0312 | 0.12 |
| ZNP ewes all years | 3 | 0.08 | 0.002 | - | - | 0.42 |
| ZNP 2018 | 6 | 0.21 | 0.003 | 127 | 0.639 | 0.19 |
| ZNP 2019 | 7 | 0.19 | 0.004 | 292 | 0.027 | 0.12 |
| ZNP 2020 | 8 | 0.13 | 0.003 | 140.5 | 0.309 | 0.11 |
| ZNP ewes 2018 | 3 | 0.35 | 0.002 | - | - | 0.23 |
| ZNP ewes 2019 | 4 | 0.1 | 0.002 | - | - | 0.07 |
| ZNP ewes 2020 | 3 | 0.21 | 0.002 | - | - | 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, B.M.; Stroud-Settles, J.; Roug, A.; Manlove, K. Disease Ecology of a Low-Virulence Mycoplasma ovipneumoniae Strain in a Free-Ranging Desert Bighorn Sheep Population. Animals 2022, 12, 1029. https://doi.org/10.3390/ani12081029
Johnson BM, Stroud-Settles J, Roug A, Manlove K. Disease Ecology of a Low-Virulence Mycoplasma ovipneumoniae Strain in a Free-Ranging Desert Bighorn Sheep Population. Animals. 2022; 12(8):1029. https://doi.org/10.3390/ani12081029
Chicago/Turabian StyleJohnson, Brianna M., Janice Stroud-Settles, Annette Roug, and Kezia Manlove. 2022. "Disease Ecology of a Low-Virulence Mycoplasma ovipneumoniae Strain in a Free-Ranging Desert Bighorn Sheep Population" Animals 12, no. 8: 1029. https://doi.org/10.3390/ani12081029
APA StyleJohnson, B. M., Stroud-Settles, J., Roug, A., & Manlove, K. (2022). Disease Ecology of a Low-Virulence Mycoplasma ovipneumoniae Strain in a Free-Ranging Desert Bighorn Sheep Population. Animals, 12(8), 1029. https://doi.org/10.3390/ani12081029