
Atolla reynoldsi sp. nov. (Cnidaria, Scyphozoa, Coronatae, Atollidae): A New Species of Coronate Scyphozoan Found in the Eastern North Pacific Ocean †

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. ROV Collections
2.2. DNA Extraction and Amplification
2.3. DNA Analyses
3. Results
3.1. Collection Information
3.1.1. Atolla Species Sequenced
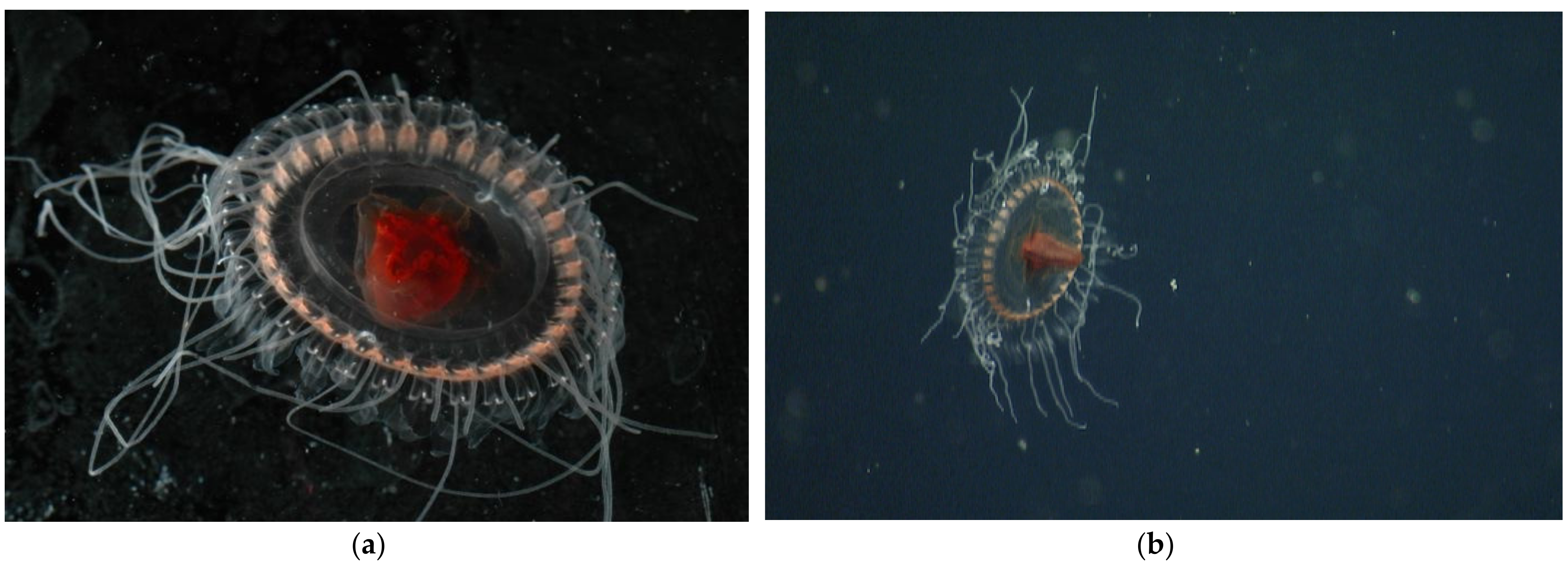
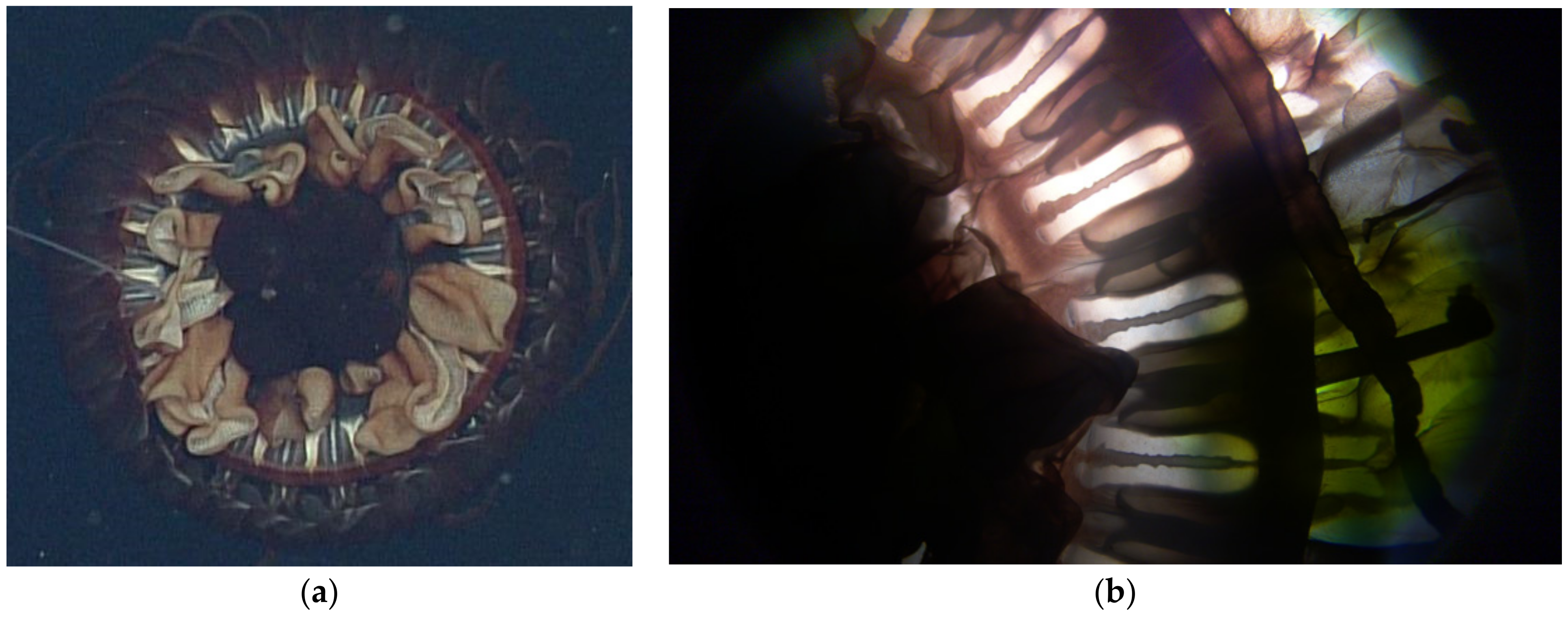
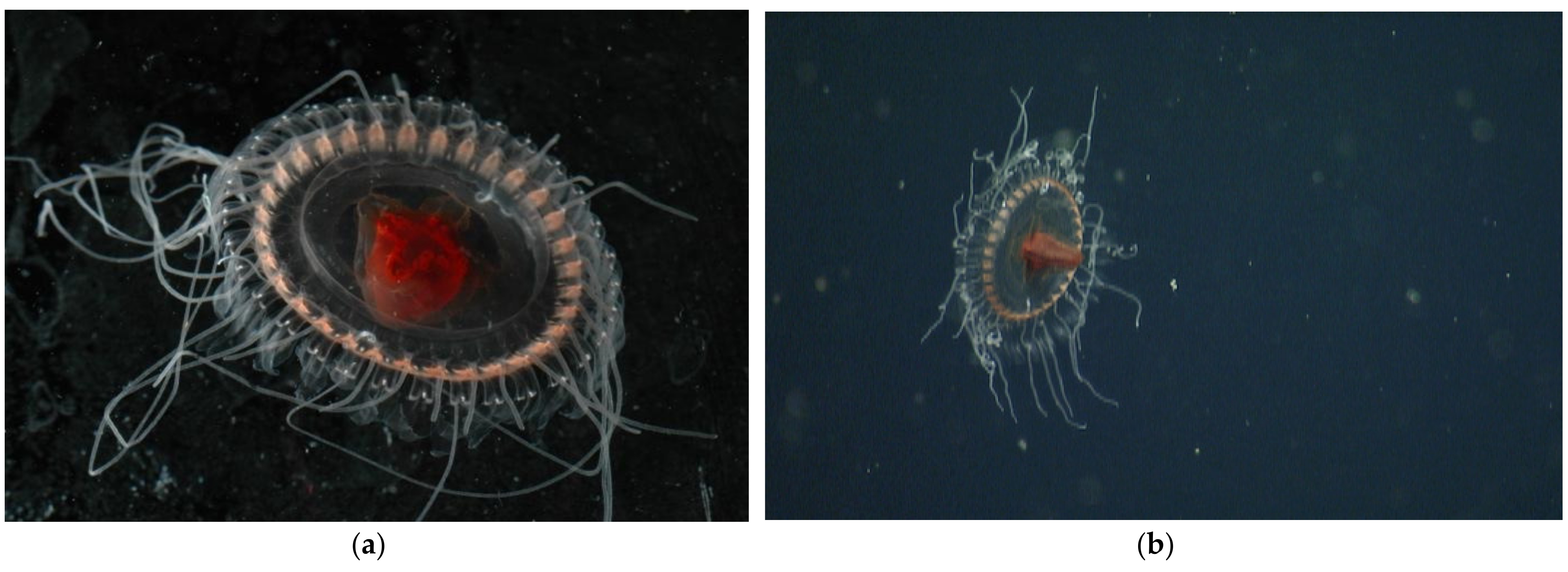
3.1.2. Atolla reynoldsi sp. nov.
3.1.3. Atolla Species A
3.1.4. Atolla Species B
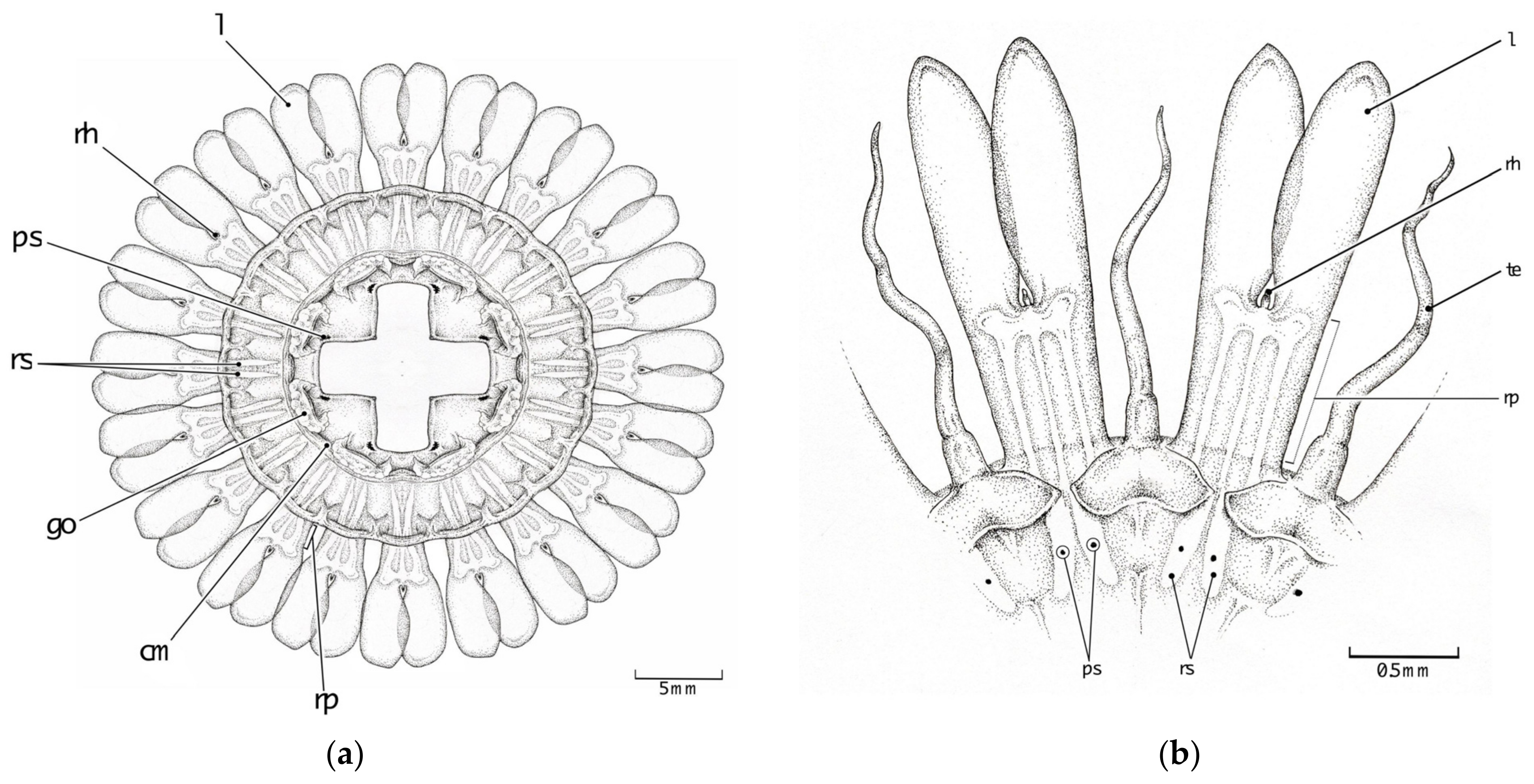
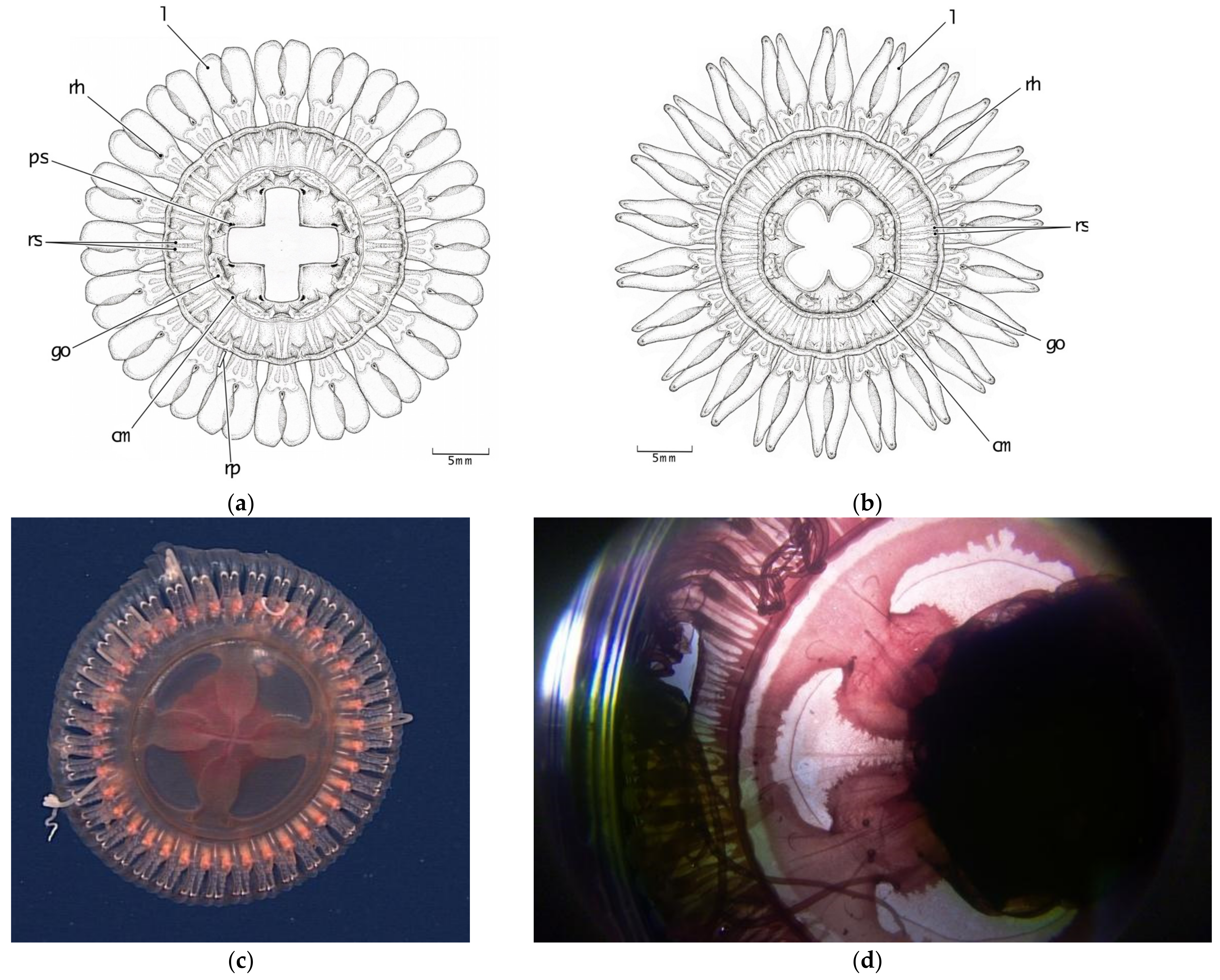
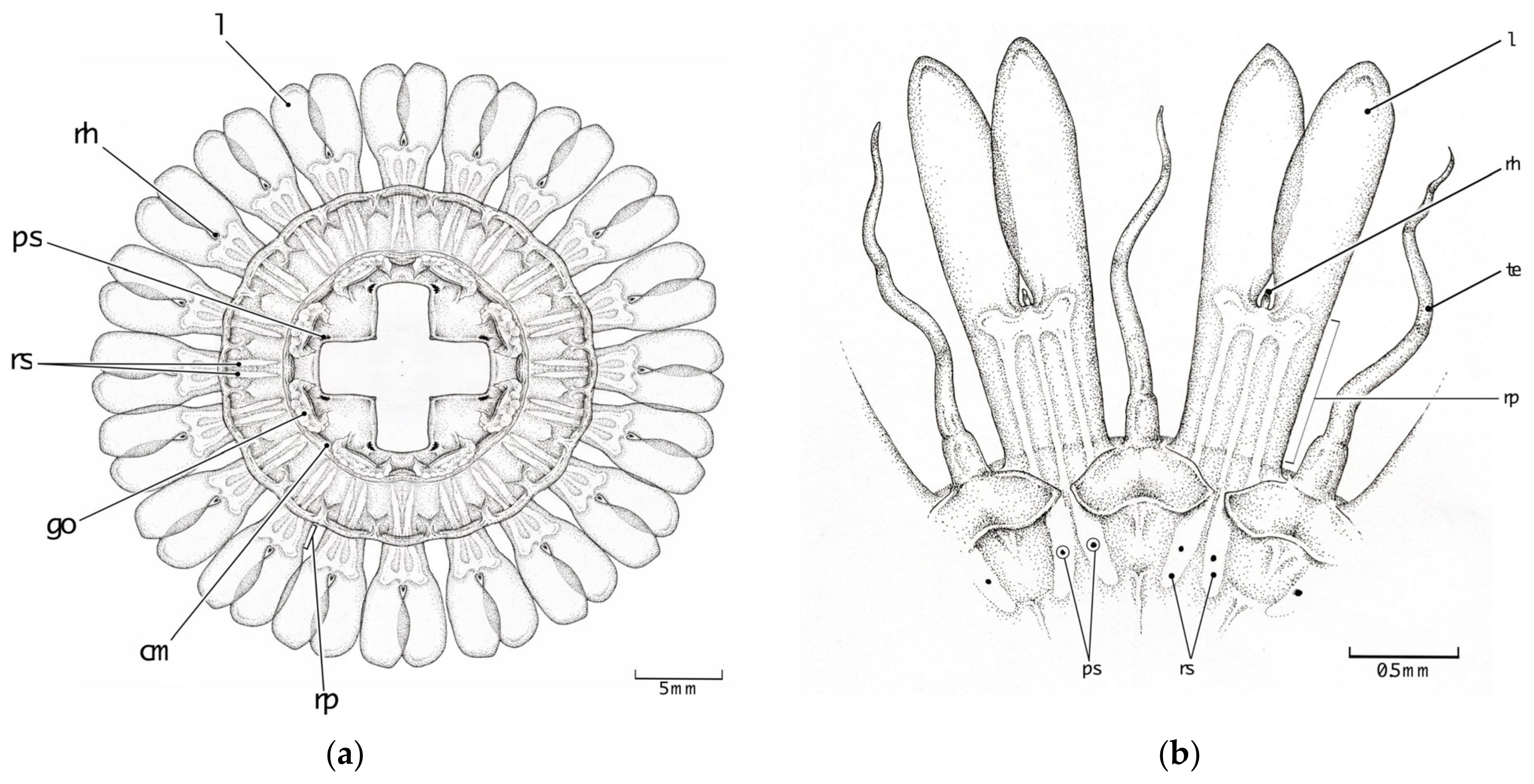
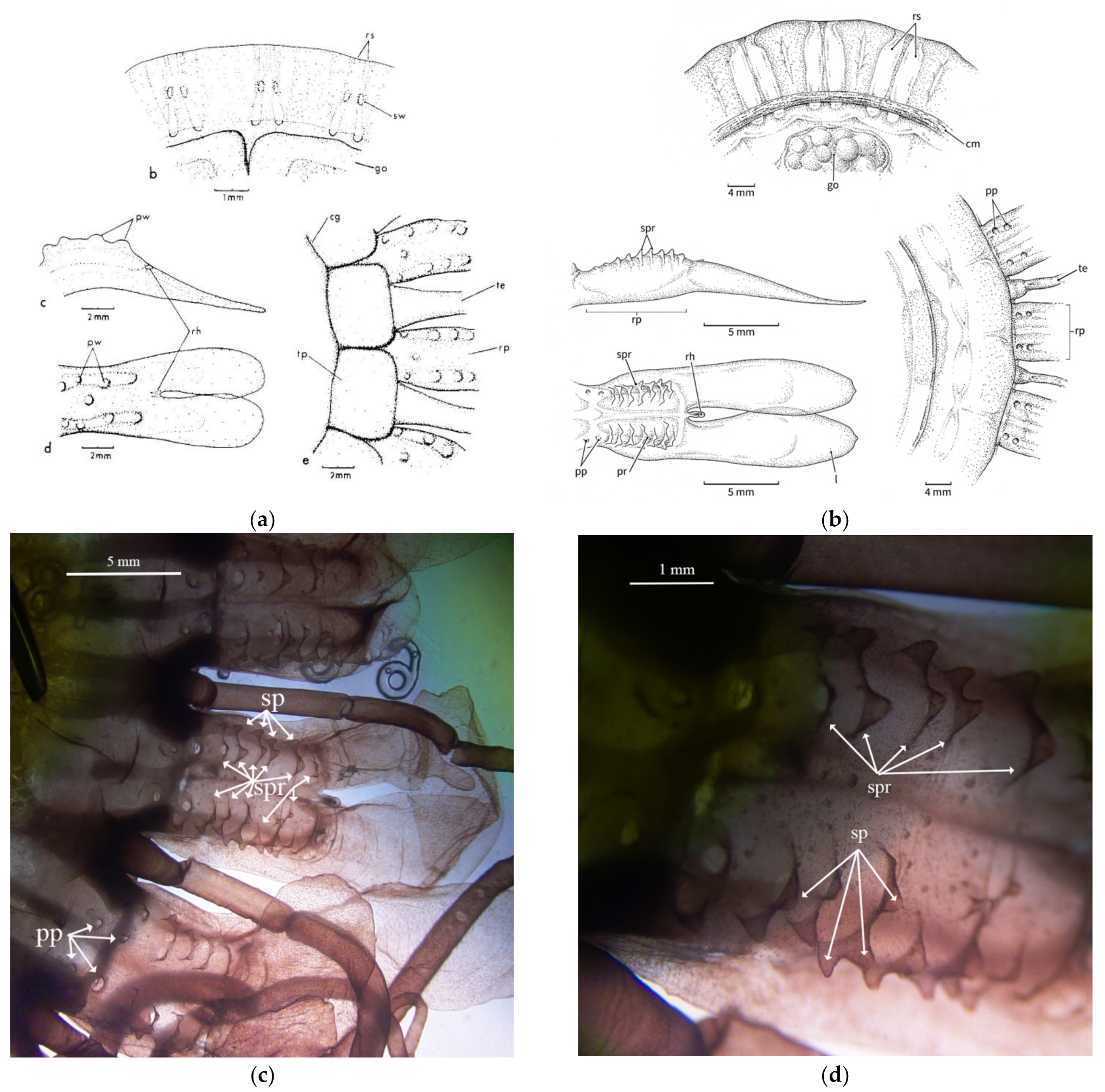
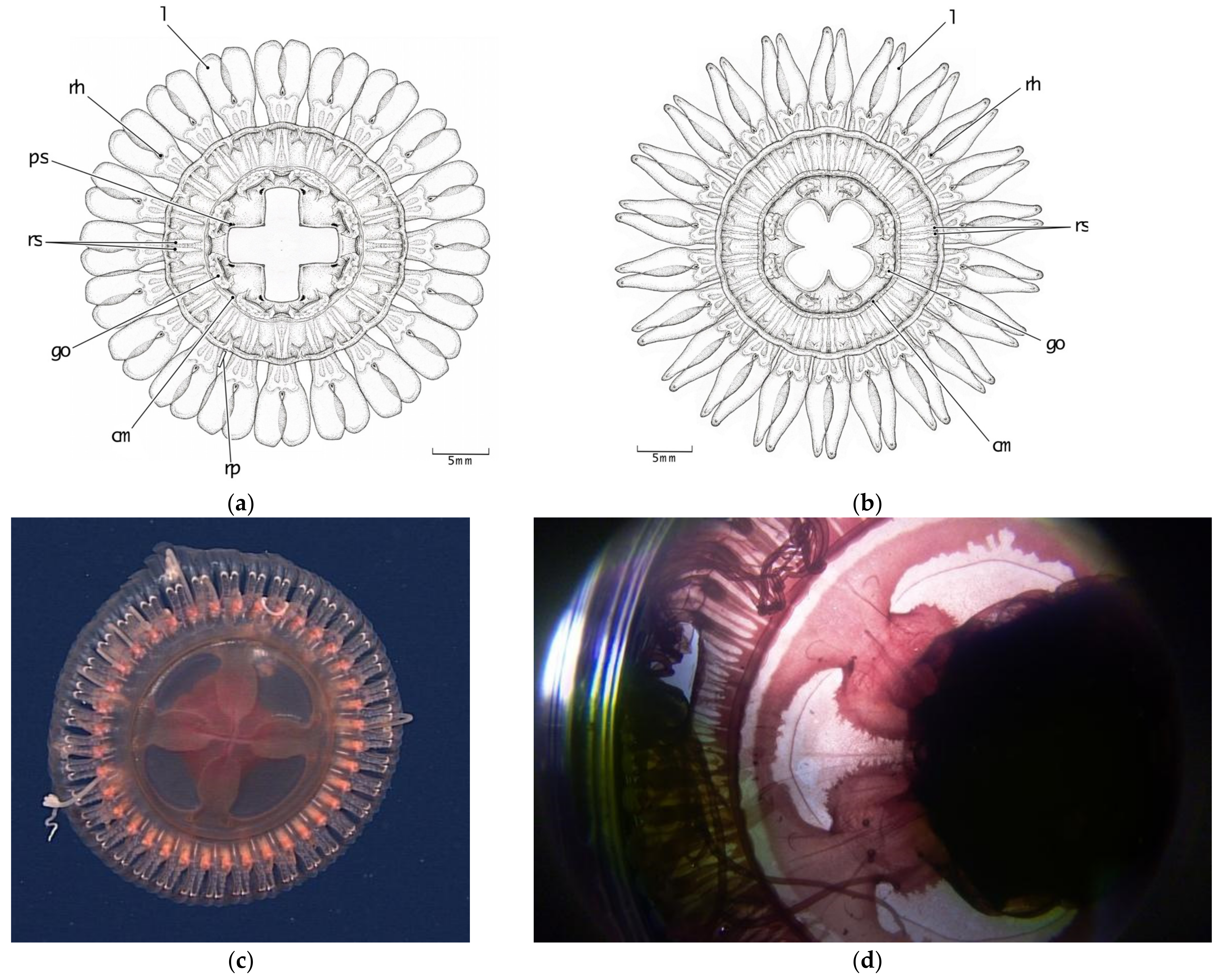
3.2. Morphological Distinctions
3.2.1. Pigmentation
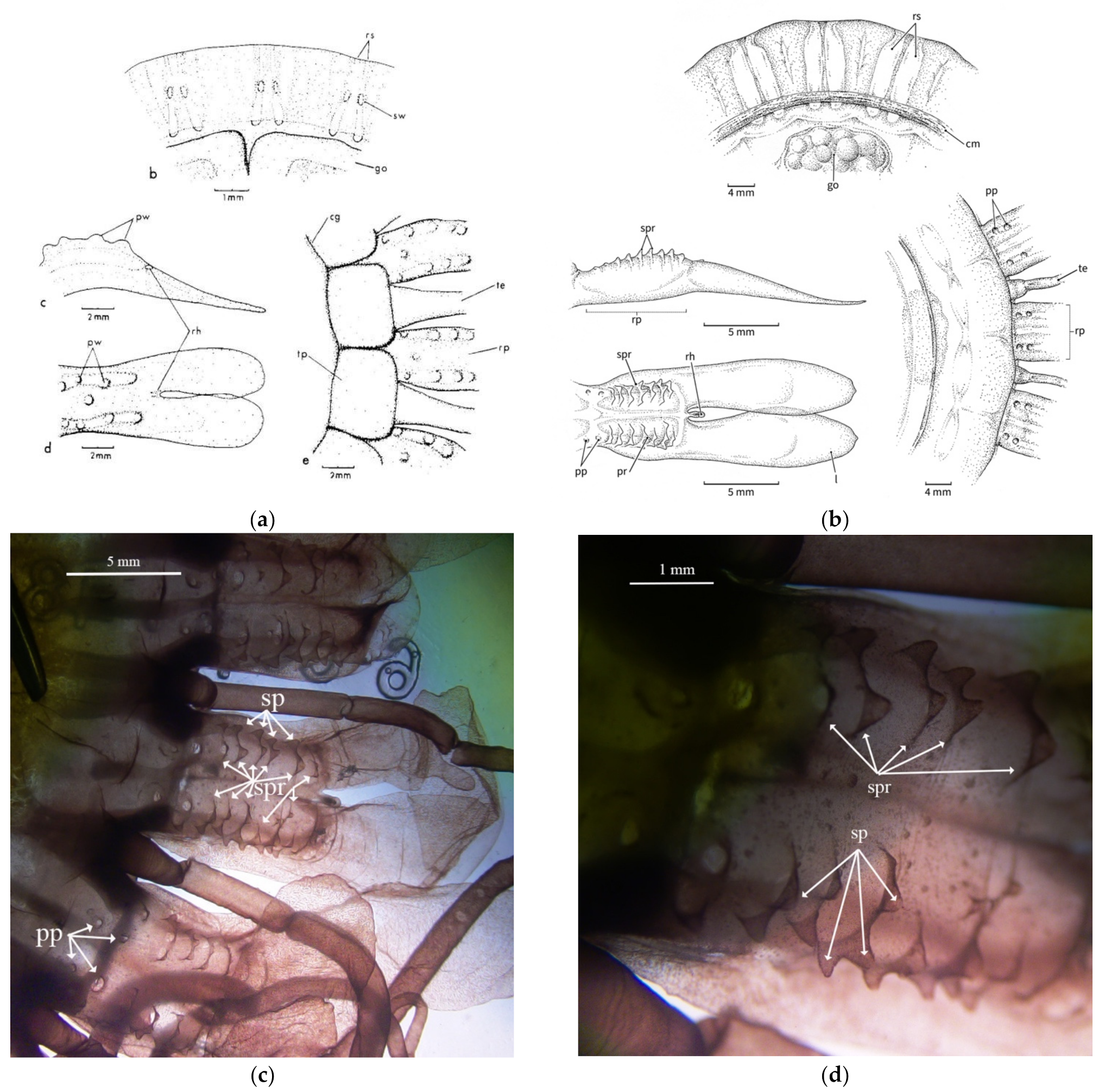
3.2.2. Papillae
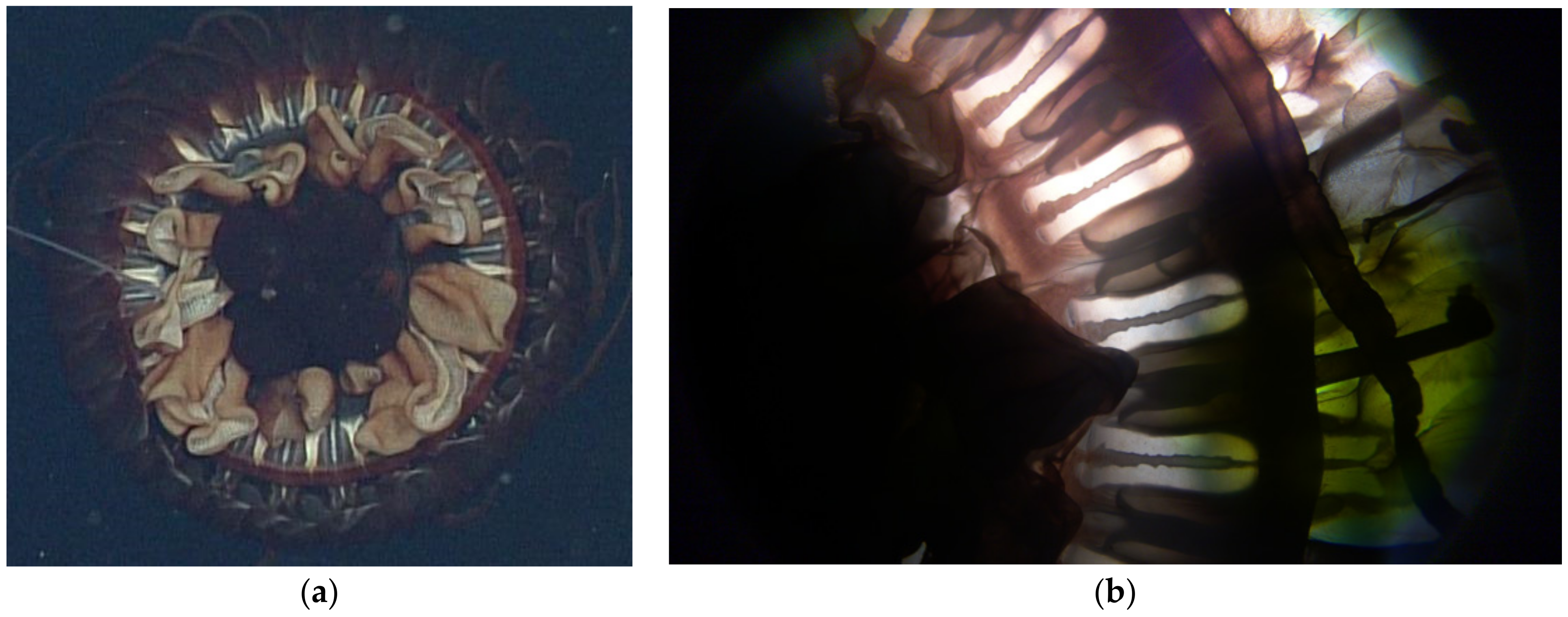
3.2.3. Stomach Morphology
3.2.4. Radial Septa
3.2.5. Tentacles
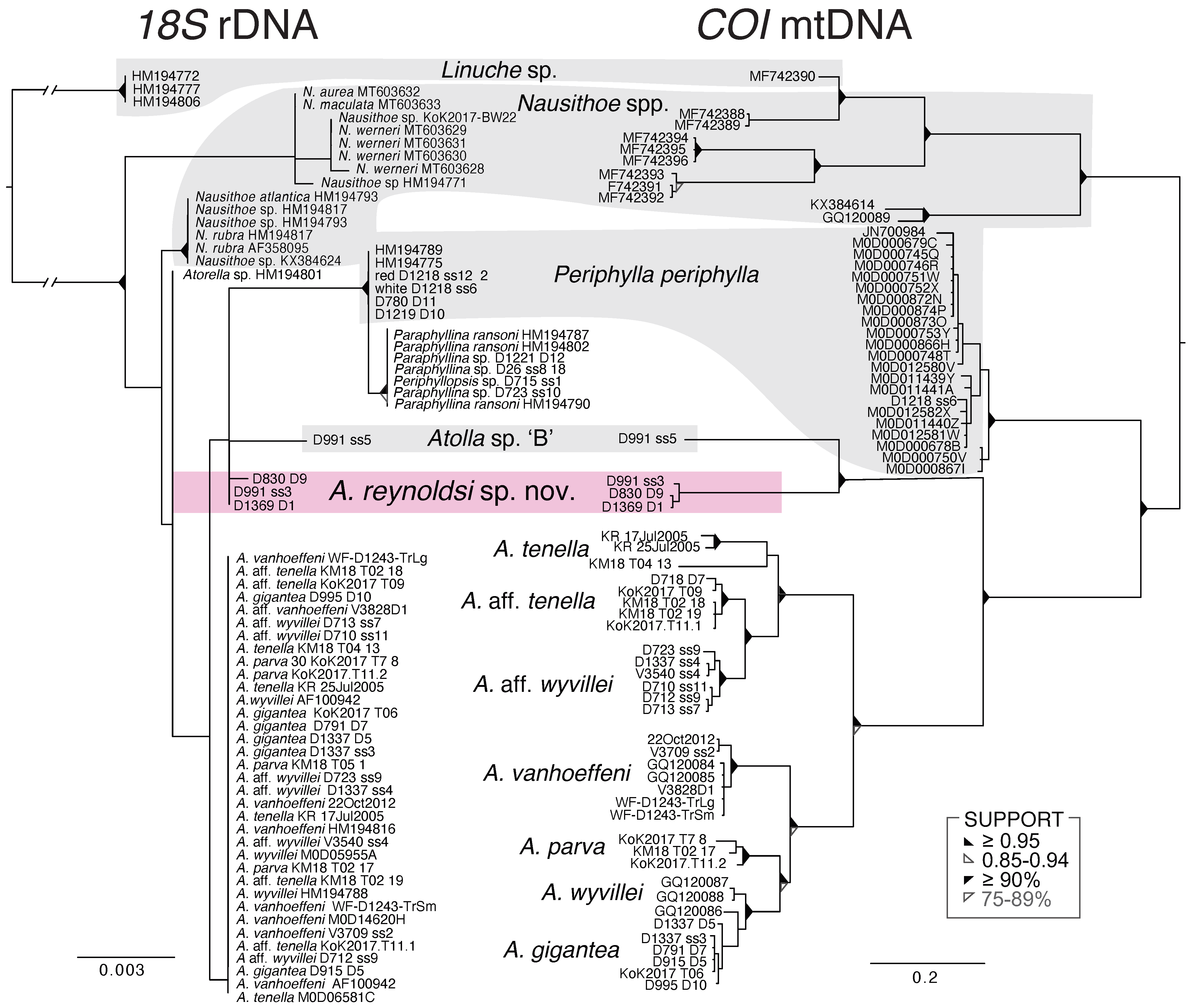
3.3. Molecular Results
4. Discussion
4.1. Systematics
4.2. Molecular Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species ID | Date | Sample ID | Depth m | Temp °C | Sal PSU | Oxy ml/L | Lat Decimal | Long Decimal |
|---|---|---|---|---|---|---|---|---|
| A. gigantea | 10 August 2015 | D0791D5 | 1112 | 3.809 | 34.457 | 0.373 | 36.531771 | −122.507713 |
| A. gigantea | 14 December 2016 | D0915D5 | 1016 | 3.852 | 34.467 | 0.411 | 36.260438 | −122.593946 |
| A. gigantea | 10 December 2017 | D0995D10 | 897 | 4.199 | 34.427 | 0.297 | 36.748584 | −122.103112 |
| A. gigantea | 19 March 2017 | KOK- T06 | NA | NA | NA | NA | 19.275 | 156.133333 |
| A. gigantea | 30 May 2021 | D1337sss3 | 1253 | 4.278 | 34.464 | 0.332 | 33.850068 | −119.850222 |
| A. gigantea | 30 May 2021 | D1337D5 | 936 | 4.436 | 34.45 | 0.321 | 33.850169 | −119.651757 |
| A. parva | 20 March 2017 | KOK-T7/8 | NA | NA | NA | NA | 19.483333 | 156.13333 |
| A.parva | 21 March 2017 | KOK17-T11 | NA | NA | NA | NA | 19.666667 | 156.13333 |
| A. parva | 6 November 2018 | KM-T02.17 | NA | NA | NA | NA | 19.426389 | 156.408611 |
| A. parva | 7 November 2018 | KM18 T05 | NA | NA | NA | NA | 19.318333 | 156.185833 |
| A. tenella | 17 July 2005 | KR-17 | NA | NA | NA | NA | Arctic | |
| A. tenella | 25 July 2005 | KR-25 | NA | NA | NA | NA | Arctic | |
| A. aff. tenella | 2 March 2015 | D718D7 | 1696 | 2.633 | 34.615 | 1.212 | 24.412664 | −109.095732 |
| A. aff. tenella | 20 March 2017 | KOK17-T09 | NA | NA | NA | NA | 19.483333 | 156.13333 |
| A. aff. tenella | 21 March 2017 | KOK17-T11 | NA | NA | NA | NA | 19.666667 | 156.13333 |
| A. aff. tenella | 6 November 2018 | KM- T02.18 | NA | NA | NA | NA | 19.426389 | 156.408611 |
| A. aff. tenella | 6 November 2018 | KM-T02.19 | NA | NA | NA | NA | 19.426389 | 156.408611 |
| A. tenella | 7 November 2018 | KM18 T04-13 | 2500 | NA | NA | NA | 19.318333 | 156.185833 |
| A. vanhoeffeni | 22 October 2012 | WF trawl | NA | NA | NA | NA | 36.699558 | −122.049488 |
| A. vanhoeffeni | 24 April 2013 | V3709ss2 | 512 | 6.002 | 34.251 | 0.343 | 36.700040 | −122.048422 |
| A. vanhoeffeni | 27 May 2015 | V3828D1 | 424 | 6.876 | 34.179 | 0.924 | 36.703304 | −122.052176 |
| A. vanhoeffeni | 31 January 2020 | WF-D1243trawl | NA | NA | NA | NA | 36.16666 | −119.25 |
| A. vanhoeffeni | 31 January 2020 | WF-D1243trawl | NA | NA | NA | NA | 36.16666 | −119.25 |
| A. aff. wyvillei | 18 March 2010 | V3540ss4 | 626 | 5.077 | 34.319 | 0.195 | 36.705400 | −122.053820 |
| A. aff. wyvillei | 22 February 2015 | D710ss11 | 746 | 6.020 | 34.509 | 0.023 | 24.277515 | −109.360873 |
| A. aff. wyvillei | 24 February 2015 | D712ss9 | 706 | 5.997 | 34.516 | 0.032 | 25.430911 | −109.835949 |
| A. aff. wyvillei | 25 February 2015 | D713ss7 | 774 | 6.002 | 34.516 | 0.032 | 25.446143 | −109.848168 |
| A. aff. wyvillei | 9 March 2015 | D723ss9 | 697 | 5.956 | 34.516 | 0.034 | 25.442727 | −109.852024 |
| A. aff. wyvillei | 30 May 2021 | D1337ss4 | 985 | 4.436 | 34.45 | 0.321 | 33.850169 | −119.651757 |
| A reynoldsi sp. nov. | 5 December 2015 | D0830D9 | 1013 | 3.861 | 34.436 | 0.348 | 36.688186 | −122.118768 |
| A reynoldsi sp. nov. | 6 December 2017 | D0991ss3 | 1878 | 2.229 | 34.602 | 1.373 | 36.548736 | −122.541753 |
| A reynoldsi sp. nov. | 30 July 2021 | D1369D1 | 3189 | 1.576 | 34.665 | 2.470 | 35.499466 | −123.99876 |
| Atolla type A | 30 October 2021 | D1399-ss3 | 1253 | 3.264 | 34.515 | 0.764 | 36.7009226 | −122.067752 |
| Atolla type B | 6 December 2017 | D991ss5 | 1783 | 2.363 | 34.592 | 1.271 | 36.548400 | −122.542593 |
| Nausithoe sp. | 23 March 2017 | KOK2017-BW22 | 30 | NA | NA | NA | 20.756111 | −157.255833 |
| Paraphyllina sp. | 27 May 2019 | D0026 ss8 | 2385 | 1.858 | 34.592 | 1.943 | 36.116665 | −122.75 |
| Paraphyllina sp. | 9 March 2015 | D0723 ss10 | 651 | 6.199 | 34.512 | 0.025 | 25.442516 | −109.852324 |
| Paraphyllina sp. | 18 November 2019 | D1221 D12 | 2088 | 2.006 | 34.815 | 1.851 | 36.545798 | −122.538197 |
| Periphyllopsis sp. | 27 February 2015 | D0715 ss1 | 1761 | 2.839 | 34.819 | 1.087 | 28.182585 | −119.599956 |
| Periphylla periphylla | 1 July 2015 | D0780 D11 | 534 | 5.231 | 34.169 | 0.433 | 36.15082 | −124.2852 |
| Periphylla periphylla | 15 November 2015 | D1218 ss12 | 384 | 7.542 | 34.158 | 0.996 | 36.695557 | −122.004649 |
| Periphylla periphylla | 15 November 2015 | D1218 ss6 | 392 | 6.940 | 34.184 | 0.763 | 36.698180 | −122.010072 |
| Periphylla periphylla | 16 November 2015 | D1219 D10 | 923 | 4.237 | 34.416 | 0.321 | 36.544387 | −122.537005 |
| Species | A. chuni * | A. parva | A. vanhoeffeni | A. tenella and A. aff. tenella | A. gigantea | A. wyvillei and A. aff. wyvillei | A. reynoldsi sp. nov. | Atolla species A—tall, rounded dome | Atolla species B—White, very flat |
|---|---|---|---|---|---|---|---|---|---|
| Pigment spots | No | No | Yes 2 per quadrant | Yes? 2 per tentacle a | No | No | No | No | no |
| Papillae | Yes | No | No | No | No | No | Yes | No | Yes, varies b |
| Ridges with spikes on rhopalia | No | No | No | No | No | No | Yes | No | No |
| Stomach | Clover shaped | Clover shaped | Cross shaped | Clover shaped | Clover shaped | Clover shaped | Greek-cross shaped | Greek-cross shaped—evaginated | Greek-cross shaped |
| Tentacles | 24 | 18–24 | 18–20 | 22–30 | 24, 28 | 22–30 | 26–39 coiled | 59–64 | 32–60 |
| Trailing tentacle | Yes | Yes | Yes | Yes | Yes | Yes | No | No | No |
| Septa path c | ? | Straight or slightly divergent, club shaped | Straight | Divergent in description; straight in our specimens | Divergent (preserved look straight) | Divergent | Straight or slightly divergent | Straight or slightly divergent | Straight |
| Septa extend to muscle d | ? | Yes | No | No | Yes | Yes e | Yes | Yes | Yes |
| Gonad shape f | Oval/bean | oval | Horseshoe | Circular with irregular edges | Horseshoe | Oval to large auricular | Oval but horseshoe shaped when mature | Horseshoe | Immature, horseshoe |
| COI mtDNA | Position | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | 91 | 94 | 97 | 100 | 103 | 106 | 109 | 118 | 122 | 124 | 125 | 127 | 133 | 151 | 158 | 160 | 163 | 166 | 172 | 179 | 181 | 196 | 199 | 205 | 208 | 220 | 223 |
| Atolla sp. B | G | T | T | T | C | A | A | T | T | A | T | A | A | T | A | A | A | A | T | C | A | A | T | A | T | T | A |
| A. gigantea | A | T | T | T | A | A | T | T | T | A | T | A | A | T | A | A | T | T | T | T | A | T | C | T | T | C | A |
| A. reynoldsi | A | G | C | C | T | C | C | C | C | T | G | G | T | A | G | G | T | T | A | T | A | G | G | G | C | A | G |
| A. parva | A | T | T | T | T | A | T | T | T | A | T | A | A | T | A | A | T | T | A | T | A | T | T | T | T | T | A |
| A. aff. tenella | A | T | T | T | G | A | T | T | T | A | T | A | A | T | A | A | T | T | T | T | G | T | T | T | T | C | A |
| A. tenella | A | T | T | T | T | A | T | T | T | A | C | A | A | T | A | A | C | T | G | T | A | T | T | T | T | T | A |
| A. vanhoeffeni | A | T | T | T | G | T | C | T | T | A | T | A | A | T | A | A | T | T | T | T | A | T | T | A | T | C | A |
| A. aff. wyvillei | A | T | T | T | T | A | T | T | T | A | T | A | A | T | A | A | T | T | A | T | A | T | T | T | T | T | A |
| A. wyvillei | A | T | T | T | A | A | T | T | T | A | T | A | A | T | A | A | C | T | T | T | A | T | C | T | T | C | A |
| COI cont. | Position | ||||||||||||||||||||||||||
| Species | 224 | 232 | 233 | 245 | 250 | 253 | 256 | 265 | 266 | 268 | 283 | 284 | 286 | 298 | 300 | 301 | 304 | 316 | 328 | 331 | 334 | 335 | 337 | 340 | 341 | 346 | 352 |
| Atolla sp. B | T | T | A | T | A | A | C | A | T | A | A | T | A | A | C | C | T | T | C | A | T | G | T | A | T | T | T |
| A. gigantea | T | T | A | C | A | A | T | T | T | A | T | T | A | A | T | T | T | T | C | A | T | G | T | T | T | T | T |
| A. reynoldsi | T | C | G | T | G | T | T | C | C | T | G | A | T | G | T | T | A | A | T | G | T | G | T | T | T | T | A |
| A. parva | T | T | A | C | C | A | T | T | T | A | T | T | A | A | T | A | T | T | T | A | T | G | T | T | T | T | T |
| A. aff. tenella | T | T | A | C | C | A | T | T | T | A | T | T | A | A | T | A | T | T | C | A | T | G | T | C | C | T | T |
| A. tenella | T | T | A | C | T | A | T | T | T | A | T | T | A | A | T | A | G | T | T | A | C | G | C | T | T | A | T |
| A. vanhoeffeni | C | T | A | C | A | A | T | C | T | A | T | T | A | A | T | T | T | T | A | A | T | A | T | T | A | T | T |
| A. aff. wyvillei | T | T | A | C | C | A | T | T | T | A | T | T | A | A | T | G | T | T | T | A | T | G | T | T | T | T | T |
| A. wyvillei | T | T | A | C | A | A | T | T | T | A | T | T | A | A | T | A | T | T | C | A | T | G | C | T | T | T | T |
| COI cont. | Position | ||||||||||||||||||||||||||
| Species | 358 | 361 | 364 | 368 | 371 | 373 | 376 | 379 | 382 | 385 | 394 | 398 | 410 | 415 | 421 | 430 | 439 | 440 | 442 | 445 | 452 | 454 | 455 | 457 | 461 | 465 | 466 |
| Atolla sp. B | A | T | A | T | A | A | A | T | T | T | T | T | T | T | C | T | T | A | A | T | A | C | A | A | C | T | T |
| A. gigantea | T | A | A | C | G | T | A | T | T | A | A | G | T | T | A | A | A | A | T | T | T | A | A | T | A | C | A |
| A. reynoldsi | T | A | G | T | G | C | T | C | T | A | A | C | C | A | A | C | T | G | G | C | G | T | C | T | C | C | A |
| A. parva | T | A | T | T | G | T | A | T | T | A | A | C | T | T | A | A | T | A | T | T | T | T | A | T | G | C | A |
| A. aff. tenella | T | A | A | T | G | C | A | T | T | A | A | G | T | T | A | A | A | A | A | T | T | A | A | T | G | C | A |
| A. tenella | T | A | T | T | G | T | A | T | T | A | A | A | T | T | A | A | C | A | A | T | T | T | A | T | G | C | A |
| A. vanhoeffeni | T | A | A | T | G | T | A | T | T | A | A | G | A | T | A | A | A | A | A | T | T | A | A | T | G | C | A |
| A. aff. wyvillei | T | A | T | T | G | T | A | T | C | A | A | C | T | T | A | A | T | A | A | T | T | T | A | T | G | C | A |
| A. wyvillei | T | A | A | T | G | T | A | T | T | A | A | G | T | T | A | A | A | A | T | T | T | A | A | T | A | C | A |
| COI cont. | Position | ||||||||||||||||||||||||||
| Species | 481 | 482 | 484 | 490 | 496 | 500 | 502 | 503 | 511 | 518 | 520 | 529 | 535 | 538 | 541 | 547 | 548 | 549 | 556 | 565 | 583 | 586 | 589 | 592 | 610 | 616 | 619 |
| Atolla sp. B | A | T | A | T | T | T | A | G | T | T | A | T | T | G | A | A | G | C | T | A | T | T | A | T | G | T | A |
| A. gigantea | T | T | A | G | A | T | A | G | T | C | T | T | A | A | A | A | G | C | A | A | T | T | C | T | A | T | T |
| A. reynoldsi | T | C | T | G | C | C | G | G | A | C | T | A | G | A | G | A | T | G | A | G | C | C | G | C | C | T | T |
| A. parva | C | T | A | A | T | T | A | G | T | C | T | T | A | T | A | G | G | C | A | A | T | T | T | T | T | T | T |
| A. aff. tenella | T | T | A | A | A | T | A | G | T | C | T | T | G | A | A | A | G | C | A | A | T | T | T | T | A | C | T |
| A. tenella | T | T | A | A | T | T | A | G | T | C | T | T | A | T | A | A | G | C | A | A | T | T | T | T | T | T | T |
| A. vanhoeffeni | T | T | A | A | A | T | A | A | C | C | T | T | A | A | A | A | G | C | A | A | T | T | T | T | G | T | T |
| A. aff. wyvillei | C | T | A | A | T | T | A | G | T | C | T | T | A | T | A | A | G | C | A | A | T | T | T | T | T | T | T |
| A. wyvillei | T | T | A | A | T | T | A | G | T | C | T | T | A | A | A | A | G | C | A | A | T | T | C | T | A | T | T |
| COI cont. | Position | ||||||||||||||||||||||||||
| Species | 640 | 646 | 652 | 664 | 670 | 671 | 673 | 676 | 677 | 685 | 694 | 695 | 697 | 700 | 712 | 718 | 721 | 724 | 760 | 763 | 769 | 778 | 779 | 781 | |||
| Atolla sp. B | T | T | G | A | T | T | A | A | C | T | A | A | A | A | T | T | T | T | T | G | G | T | T | A | |||
| A.gigantea | T | T | A | A | T | T | A | T | T | T | A | G | A | T | A | T | T | T | T | T | T | T | A | A | |||
| A. reynoldsi | C | C | T | G | T | C | T | T | T | C | G | G | T | T | A | C | T | T | C | T | A | C | C | T | |||
| A. parva | T | T | C | T | T | T | A | T | T | T | A | G | G | T | A | T | T | T | T | T | T | T | A | C | |||
| A. aff. tenella | T | T | C | T | T | T | A | T | T | T | A | G | A | T | A | T | C | C | T | T | T | T | A | A | |||
| A. tenella | T | T | C | C | C | T | G | T | T | T | A | G | A | T | A | T | T | T | T | T | T | T | A | A | |||
| A. vanhoeffeni | T | T | T | T | T | T | A | T | T | T | A | G | A | T | A | T | T | T | T | T | T | T | A | A | |||
| A. aff wyvillei | T | T | C | T | T | T | A | T | T | T | A | G | A | T | A | T | T | T | T | C | T | T | A | C | |||
| A. wyvillei | T | T | A | C | T | T | A | T | T | T | A | G | A | T | A | T | T | T | T | T | T | T | A | A | |||
References
- Götte, A. Entwickelungsgeschichte der Aurelia aurita und Cotylorhiza tuberculata. Abh. Entwickelungsgeschichte Tiere 1887, 4, 1–79. (In German) [Google Scholar]
- Calder, D.R. Cubozoan and scyphozoan jellyfishes of the Carolinian Biogeographic Province, southeastern USA. R. Ont. Mus. Contrib. Sci. 2009, 3, 1–58. [Google Scholar]
- Vanhöffen, E. Die Akalephen der Plankton-Expedition. Ergeb. Plankton-Exped. Humboldt-Stift. 1892, 2, 3–28. (In German) [Google Scholar]
- Hickson, S.J. Coelenterata and Ctenophora. In The Cambridge Natural History; Harmer, S.F., Shipley, A.E., Eds.; MacMillan and Company: London, UK, 1906; pp. 243–424. [Google Scholar]
- Haeckel, E. System der Acraspeden. Zweite Halfte des Systems der Me dusen. Denkschr. Medicinisch-Nat. Ges. Jena 1880, 2, 361–672. (In German) [Google Scholar]
- Collins, A.G.; Jarms, G.; Morandini, A.C. World List of Scyphozoa Atolla Haeckel, 1880. World Register of Marine Species. 2021. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=135248 (accessed on 30 November 2021).
- Pages, F.; Gill, J.; Bouillon, J. Medusae (Hydrozoa, Scyphozoa, Cubozoa) of the Benguela current (southeast Atlantic). Sci. Mar. 1992, 56, 1–64. [Google Scholar]
- Larson, R.J. Pelagic scyphomedusae (Scyphozoa: Coronatae and Semaestomeae) of the southern ocean. Biol. Antarct. Seas XVI Antarct. Res. Ser. 1986, 41, 59–165. [Google Scholar]
- Bigelow, H.B. The Medusae. In Reports on the Scientific Results of the Expedition to the Eastern Tropical Pacific, in Charge of Alexander Agassiz, by the U.S. Fish Commission Steamer “Albatross” from October 1904 to March 1905. XVI; Memoirs of the Museum of Comparative Zoölogy at Harvard College: Cambridge, MA, USA, 1909; Volume 37, pp. 1–243. Available online: http://www.biodiversitylibrary.org/item/30084 (accessed on 30 November 2021).
- Gibbons, M.J.; Morandini, A.C.; Straehler-Pohl, I.; Bezio, N. Identification Guide to Macro Jellyfishes of West Africa; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Russell, F.S. The Medusae of the British Isles II. Pelagic Scyphozoa with a Supplement to the First Volume on Hydromedusae; Cambridge University Press: London, UK, 1970; 296p. [Google Scholar]
- Hartlaub, C. Méduses. In Croisière océanographique: Accomplie à bord de la Belgica dans la Mer du Grönland, 1905; C. Bulens: Bruxelles, Belgium, 1909; pp. 463–478. Available online: https://www.biodiversitylibrary.org/page/41467593 (accessed on 30 November 2021). (In French)
- Raskoff, K.A.; Purcell, J.E.; Hopcroft, R.R. Gelatinous zooplankton of the Arctic Ocean: In situ observations under the ice. Polar Biol. 2005, 28, 207–217. [Google Scholar] [CrossRef]
- Russell, F.S. Some observations on the scyphomedusa Atolla. J. Mar. Biol. Assoc. UK 1959, 3833, 40. [Google Scholar]
- Hunt, J.C.; Lindsay, D.J. Observations on the behavior of Atolla (Scyphozoa: Coronatae) and Nanomia (Hydrozoa: Physonectae): Use of the hypertrophied tentacle in prey capture. Plankton Biol. Ecol. 1998, 45, 239–242. [Google Scholar]
- Walker, A. The Form and Function of the Hypertrophied Tentacle of the Deep-Sea Jelly Atolla spp. Unpublished Intern Report. 2011, pp. 1–19. Available online: https://www.mbari.org/wp-content/uploads/2015/10/Walker.pdf (accessed on 30 November 2021). Unpublished Intern Report.
- Larson, R.J.; Mills, C.E.; Harbison, G.R. Western Atlantic midwater hydrozoan and scyphozoan medusae: In situ studies using manned submersibles. Hydrobiologia 1991, 216, 311–317. [Google Scholar] [CrossRef]
- Moore, P.G.; Rainbow, P.S.; Larson, R.J. The mesopelagic shrimp Notostomus robustus Smith (Decapoda: Oplophoridae) observed in situ feeding on the medusan Atolla wyvillei Haeckel in the Northwest Atlantic, with notes on gut contents and mouthpart morphology. J. Crustacean Biol. 1993, 13, 690–696. [Google Scholar] [CrossRef]
- Robison, B.H.; Reisenbichler, K.R.; Sherlock, R.E. The coevolution of midwater research and ROV technology at MBARI. Oceanography 2017, 30, 26–37. [Google Scholar] [CrossRef]
- Youngbluth, M.J. Manned Submersibles and Sophisticated Instrumentations: Tools for Oceanographic Research. In SUBTECH ’83 Symposium: The Design and Operation of Underwater Vehicles. Soc. Underw. Technol. 1984, 22, 351–355. [Google Scholar]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.B.; Hoeh, W.R.; Lutz, R.A.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernoor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. Q-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Vanhöffen, E. Die Acraspeden Medusen der deutschen Tiefsee-Expedition 1898–1899. Wiss. Ergeb. Dtsch. Tiefsee Exped. Dampfer Valdivia 1898 1899, 1902, 3–52. (In German) [Google Scholar] [CrossRef] [Green Version]
- Russell, F.S. On a new species of scyphomedusa, Atolla vanhöffeni n. sp. J. Mar. Biol. Assoc. UK 1957, 36, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, D.; Furushima, Y.; Miyake, H.; Kitamura, M.; Hunt, J.C. The Scyphomedusan Fauna of the Japan Trench: Preliminary Results from a Remotely-Operated Vehicle. In Coelenterate Biology; Springer: Dordrecht, The Netherlands, 2004; pp. 537–547. [Google Scholar]


| Species ID | Date | Accession Number 18s | Accession Number COI | Sample ID | Depth m | Lat Decimal | Long Decimal | Tentacle Number |
|---|---|---|---|---|---|---|---|---|
| A. gigantea | 10 August 2015 | OM260068 | OM214504 | D0791D7 | 1112 | 36.531771 | −122.507713 | 24 |
| A. gigantea | 14 December 2016 | OM260069 | OM214505 | D0915D5 | 1016 | 36.260438 | −122.593946 | 24 |
| A. gigantea | 10 December 2017 | OM260070 | OM214503 | D0995D10 | 897 | 36.748584 | −122.103112 | 24 |
| A. gigantea | 19 March 2017 | OM260073 | OM214506 | KOK-T06 | 1584–888 | 19.275 | 156.133333 | 59 |
| A. gigantea | 30 May 2021 | OM260072 | OM214502 | D1337ss3 | 1253 | 33.850068 | −119.850222 | 24 |
| A. gigantea | 30 May 2021 | OM260071 | OM214501 | D1337D5 | 936 | 33.850169 | −119.651757 | 24 |
| A. parva | 20 March 2017 | OM260077 | OM214507 | KOK-T7/8 | 838–581 | 19.483333 | 156.13333 | 30 |
| A. parva | 21 March 2017 | OM260076 | OM214509 | KOK17-T11 | 1700–1115 | 19.666667 | 156.13333 | 30 |
| A. parva | 6 November 2018 | OM260074 | OM214508 | KM-T02.17 | NA | 19.426389 | 156.408611 | 26 |
| A. parva | 7 November 2018 | OM260075 | NA | KM18-T05 | NA | 19.318333 | 156.185833 | 26 |
| A. tenella | 17 July 2005 | OM260079 | OM214511 | KR−17 | NA | Arctic | NA | |
| A. tenella | 25 July 2005 | OM260080 | OM214512 | KR−25 | NA | Arctic | NA | |
| A. aff. tenella | 2 March 2015 | NA | OM214513 | D718D7 | 1696 | 24.412664 | −109.095732 | 22 |
| A. aff. tenella | 20 March 2017 | OM260060 | OM214515 | KOK17-T09 | 1719–1033 | 19.483333 | 156.13333 | 24 |
| A. aff. tenella | 21 March 2017 | OM260061 | OM214516 | KOK17-T11 | 1700–1115 | 19.666667 | 156.13333 | 30 |
| A. aff. tenella | 6 November 2018 | OM260058 | OM214517 | KM-T02.18 | NA | 19.426389 | 156.408611 | 30 |
| A. aff. tenella | 6 November 2018 | OM260059 | OM214514 | KM-T02.19 | NA | 19.426389 | 156.408611 | 30 |
| A. tenella | 7 November 2018 | OM260078 | OM214510 | KM18 T04-13 | 2500 | 19.318333 | 156.185833 | 30 |
| A. vanhoeffeni | 22 October 2012 | OM260084 | OM214496 | WF trawl | NA | 36.699558 | −122.049488 | NA |
| A. vanhoeffeni | 24 April 2013 | OM260085 | OM214497 | V3709ss2 | 512 | 36.700040 | −122.048422 | NA |
| A. vanhoeffeni | 27 May 2015 | OM260081 | OM214498 | V3828D1 | 424 | 36.703304 | −122.052176 | NA |
| A. vanhoeffeni | 31 January 2020 | OM260082 | OM214499 | WF-trawl | NA | 36.16666 | −119.25 | NA |
| A. vanhoeffeni | 31 January 2020 | OM260083 | OM214500 | WF-trawl | NA | 36.16666 | −119.25 | NA |
| A. aff. wyvillei | 18 March 2010 | OM260067 | OM214523 | V3540ss4 | 626 | 36.705400 | −122.053820 | NA |
| A. aff. wyvillei | 22 February 2015 | OM260062 | OM214518 | D710ss11 | 746 | 24.277515 | −109.360873 | 22 |
| A. aff. wyvillei | 24 February 2015 | OM260063 | OM214519 | D712ss9 | 706 | 25.430911 | −109.835949 | 22 |
| A. aff. wyvillei | 25 February 2015 | OM260064 | OM214520 | D713ss7 | 774 | 25.446143 | −109.848168 | 22 |
| A. aff. wyvillei | 9 March 2015 | OM260065 | OM214521 | D723ss9 | 697 | 25.442727 | −109.852024 | 22 |
| A. aff. wyvillei | 30 May 2021 | OM260066 | OM214522 | D1337ss4 | 985 | 33.850169 | −119.651757 | 24 |
| A reynoldsi sp. nov. | 5 December 2015 | OM260086 | OM214493 | D0830D9 | 1013 | 36.688186 | −122.118768 | 32 |
| A reynoldsi sp. nov. | 6 December 2017 | OM260087 | OM214492 | D0991ss3 | 1878 | 36.548736 | −122.541753 | 32 |
| A reynoldsi sp. nov. | 30 July 2021 | OM260088 | OM214494 | D1369D1 | 3189 | 35.499466 | −123.99876 | 38 |
| Atolla type B | 6 December 2017 | OM260056 | OM214495 | D991ss5 | 1783 | 36.548400 | −122.542593 | 60 |
| Atolla type A | 30 October 2021 | OM260057 | NA | D1399ss3 | 1253 | 36.700923 | −122.067752 | 59 |
| Date | Sample ID | Depth m | Temp °C | Sal PSU | Oxy ml/L | Lat Decimal | Long Decimal | Tentacle Number | Diameter cm | Status |
|---|---|---|---|---|---|---|---|---|---|---|
| 4 April 2006 | T0960ss6 | 2848 | 1.657 | 34.621 | 2.29 | 36.573417 | −122.5221505 | 38 | 13 | CHN |
| 7 April 2006 | T0964ss5 | 1400 | 2.897 | 34.5 | 0.81 | 36.328862 | −122.898496 | 26 | NA | Frozen @ |
| 20 June 2006 | T0996D6 damaged | 1435 | 2.987 | 34.489 | 0.71 | 36.551573 | −122.5109275 | 30 | 7.5 | Damaged |
| 20 October 2009 | D0087D8 CASIZ 233652 | 1133 | 3.427 | 34.401 | 0.639 | 36.334888 | −122.917099 | 32 | 8 | Paratype @ |
| 9 November 2013 | D0546D12 CASIZ 233653 | 2705 | 1.697 | 34.656 | 2.354 | 36.535624 | −122.508432 | 39 | 7.3 | Paratype @ |
| 3 August 2014 | D0642D11 | 1500 | 2.777 | 34.541 | 0.858 | 36.533088 | −122.509816 | 36 | 8 | MBARI |
| 5 December 2015 | D0830D9 | 1013 | 3.861 | 34.436 | 0.348 | 36.688186 | −122.118768 | 32 | NA | Frozen $ |
| 6 December 2017 | D0991ss3 | 1878 | 2.229 | 34.602 | 1.373 | 36.548736 | −122.541753 | 32 | 5.8 | MBARI $ |
| 10 August 2018 | D1050D11 CASIZ 233650 | 1445 | 2.906 | 34.528 | 0.821 | 36.534146 | −122.565410 | 26 | 7.7 | Paratype |
| 30 July 2021 | D1369D1 CASIZ 233651 | 3189 | 1.576 | 34.665 | 2.470 | 35.499466 | −123.99876 | 38 | 8.5 | Holotype $ |
| Date | Sample ID | Depth m | Temp °C | Sal PSU | Oxy mL/L | Lat Decimal | Long Decimal | Tentacle Number | Diameter cm | Status |
| 13 June 2002 | T0439 | 1197 | 3.371 | 34.342 | 0.62 | 36.329753 | −122.900502 | 59 | NA | NA@ |
| 30 October 2021 | D1399ss3 | 1253 | 3.264 | 34.515 | 0.764 | 36.700923 | −122.067752 | 59 | 8.5 | & |
| 14 November 2021 | D1402ss9 | 1913 | 2.210 | 34.603 | 1.434 | 36.543934 | −122.536996 | 64 | 5.4 | &@ |
| Date | Sample ID | Depth m | Temp °C | Sal PSU | Oxy mL/L | Lat Decimal | Long Decimal | Tentacle Number | Diameter cm | Status |
|---|---|---|---|---|---|---|---|---|---|---|
| 18 November 2004 | T764 | 3247 | 1.603 | 34.6 | 2.61 | 36.329555 | −122.899157 | 39 | 5.5 | & |
| 22 June 2006 | T0998D4 | 3275 | 1.632 | 34.589 | 2.41 | 36.341234 | −122.916458 | 32 | 1.7 | & |
| 14 April 2007 | T1088D4 | 2570 | 1.807 | 34.437 | 2.12 | 36.551916 | −122.502087 | 32 | NA | NA |
| 20 May 2014 | D0613ss8 | 3302 | 1.697 | 34.652 | 2.47 | 36.501258 | −122.866931 | 42 | 7.4 | & |
| 6 December 2017 | D0991ss5 | 1783 | 2.363 | 34.592 | 1.271 | 36.548400 | −122.542593 | 60 | NA | $@ |
| Species ID | Date | Accession Number 18s | Sample ID | Depth m | Lat Decimal | Long Decimal |
|---|---|---|---|---|---|---|
| Nausithoe sp. | 23 March 2017 | OM237455 | KOK2017-BW22 | 30 | 20.756111 | −157.255833 |
| Paraphyllina sp. | 27 May 2019 | OM201136 | D0026 ss8 | 2385 | 36.116665 | −122.75 |
| Paraphyllina sp. | 9 March 2015 | OM201137 | D0723 ss10 | 651 | 25.442516 | −109.852324 |
| Paraphyllina sp. | 18 November 2019 | OM201138 | D1221 D12 | 2088 | 36.545798 | −122.538197 |
| Periphyllopsis sp. | 27 February 2015 | OM201143 | D0715 ss1 | 1761 | 28.182585 | −119.599956 |
| Periphylla periphylla | 1 July 2015 | OM201139 | D0780 D11 | 534 | 36.15082 | −124.2852 |
| Periphylla periphylla | 15 November 2015 | OM201141 | D1218 ss12 | 384 | 36.695557 | −122.004649 |
| Periphylla periphylla | 15 November 2015 | OM201140 | D1218 ss6 | 392 | 36.698180 | −122.010072 |
| Periphylla periphylla | 16 November 2015 | OM201142 | D1219 D10 | 923 | 36.544387 | −122.537005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, G.I.; Christianson, L.M.; Robison, B.H.; Haddock, S.H.D.; Johnson, S.B. Atolla reynoldsi sp. nov. (Cnidaria, Scyphozoa, Coronatae, Atollidae): A New Species of Coronate Scyphozoan Found in the Eastern North Pacific Ocean. Animals 2022, 12, 742. https://doi.org/10.3390/ani12060742
Matsumoto GI, Christianson LM, Robison BH, Haddock SHD, Johnson SB. Atolla reynoldsi sp. nov. (Cnidaria, Scyphozoa, Coronatae, Atollidae): A New Species of Coronate Scyphozoan Found in the Eastern North Pacific Ocean. Animals. 2022; 12(6):742. https://doi.org/10.3390/ani12060742
Chicago/Turabian StyleMatsumoto, George I., Lynne M. Christianson, Bruce H. Robison, Steven H. D. Haddock, and Shannon B. Johnson. 2022. "Atolla reynoldsi sp. nov. (Cnidaria, Scyphozoa, Coronatae, Atollidae): A New Species of Coronate Scyphozoan Found in the Eastern North Pacific Ocean" Animals 12, no. 6: 742. https://doi.org/10.3390/ani12060742
APA StyleMatsumoto, G. I., Christianson, L. M., Robison, B. H., Haddock, S. H. D., & Johnson, S. B. (2022). Atolla reynoldsi sp. nov. (Cnidaria, Scyphozoa, Coronatae, Atollidae): A New Species of Coronate Scyphozoan Found in the Eastern North Pacific Ocean. Animals, 12(6), 742. https://doi.org/10.3390/ani12060742