1. Introduction
Species ecological niche is a complex concept that reflects relationships to the environment, which are developed throughout the evolution of each species [
1,
2,
3]. By adapting morphological, ecological, and physiological characters, species occupy ecological niches defined by environmental conditions and available resources. When two species co-exist in the same habitat, the habitat overlap occurs [
4]. Under the conditions of habitat overlap, species compete for the common resources and this competition ultimately results in the local extinction of a less effective competitor [
5] and/or in niche partitioning [
6]. This process allows competing species to utilise the same resources in different ways, and thus promotes the co-existence of species [
7].
Habitat loss may contribute to the increased habitat overlap in European landscapes due to agricultural practices and deforestation [
8,
9]. To avoid this overlap, species are colonising other suitable and available habitats, such as urban areas [
10]. Colonisation of urban areas by birds has been occurring since the middle of the 20th century, and it is a process consisting of three stages: Arrival, adjustment, and spread [
11]. At the same time, closely related species require a long time to develop interspecific differences [
12]. Moreover, the number of species that have adapted to cities is still low since a small amount of species have developed the ecological and life history traits that lead them to be urban-tolerant species [
13]. As a result, low interspecific competition and low species richness occur in cities, which are accompanied with high habitat overlap in urban areas.
The members of
Columbidae family rank among the most common and widely distributed species in European landscapes. Their distribution overlaps in habitats, such as farmlands, urban areas, and natural forests [
9]. To date, previous studies have analysed diet [
14] and nest site overlap [
15] in the
Columbidae family. Additionally, several studies on
Streptopelia turtur (Linnaeus, 1758) in the Mediterranean region focus on the effects of different types of landscape on the species [
2,
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
13,
14,
15,
16]. However, to our knowledge, there are no records of studies concerning the distribution and habitat overlap of
Columbidae species breeding in the Czech Republic, i.e., feral pigeon (
Columba livia forma domestica Linnaeus, 1766), stock dove (
Columba oenas Linnaeus, 1758), wood pigeon (
Columba palumbus Linnaeus, 1758), Eurasian collared dove (
Streptopelia decaocto Frivaldszky, 1838), and European turtle dove (
Streptopelia turtur). According to a study on the performance of molecular phylogeny,
Columba and
Streptopelia species belong to different clades, i.e.,
Columba belong to the Old World clade and
Streptopelia to a separate clade [
17]. These birds may be an interesting group for studying habitat overlap due to the recent colonization of urban areas by some of these species [
18,
19]. For instance, in Finland, researchers have reported an increase in the abundance of
C. palumbus in cities [
20]. Furthermore,
C. palumbus and
S. decaocto are recognised as urban species and their populations are increasing in urban areas in Britain [
21] and Baltic region [
22]. In addition, according to specialisation traits in species tolerant to urbanisation,
C. livia has the second highest relative urban tolerance score [
13]. From the case of the Czech Republic,
S. turtur is not considered an urban species due to its feeding behaviour, which requires open habitats, such as farmlands and grasslands [
23].
S. decaocto is almost exclusively an urban species, and
C. palumbus colonised urban areas recently in comparison to other European countries, where the colonisation process occurred earlier in the century [
11].
Quantifying habitat overlap provides insight into species competition in specific habitats, thus allowing us to understand in which habitats competition is occurring and which types of land use require more conservation attention [
2]. Here, we specifically focus on the role of landscape heterogeneity in the association with
Columbidae species since it can provide more insight into species habitat type affinities. For instance, whether they are attracted to open space habitats with lower edge density, such as farmlands or to habitats with high edge density and fragments, such as urban areas. For this purpose, our study assesses whether two landscape heterogeneity metrics, land use richness and edge density, are significant predictors of
Columbidae species distribution.
Moreover, studying the ecology of species, such as
C. oenas and
S. turtur, can provide insight for conservation strategies due to their potential to serve as umbrella species. Commonly, effective umbrella species should cover a large geographical area, and have high spatial and habitat overlap with co-occurring species [
24,
25]. For instance,
S. turtur is a vulnerable species listed on the Annex II of the Birds Directive, that aims to allow for the hunting of species to be sustainable [
26]. Therefore, the prohibition of hunting
S. turtur during specific periods allows other farmland species to benefit from these measures, as well.
The main aim of our study is to quantify the habitat overlap of five species of the
Columbidae family present in the Czech Republic regarding land use composition and land use heterogeneity. Within this goal, our objectives were to map the species, calculate the species habitat overlap index, assess the habitat selection, and determine the relationship of land use types and landscape heterogeneity with the occurrence of the following five species:
C. livia,
C. oenas,
C. palumbus,
S. decaocto, and
S. turtur. We predicted a high level of habitat overlap among most of the species, and that the habitat overlap would occur in farmlands and urban areas [
11] due to the species spatial distribution and the detected increasing number of
Columbidae species in urban areas. Furthermore, regarding species phylogeny, we can predict two possible outcomes: (i) Species within the same clade will show a greater overlap than distantly related species from different clades due to the relatively short time since their divergence; (ii) closely related species from the same clade will show a smaller overlap due to the higher interspecific competition.
2. Materials and Methods
The data were collected at 118 study sites scattered in different land use types and altitudes throughout the Czech Republic within the Bird Breeding Monitoring Program in 2015 and 2016. The Program is conducted by voluntary ornithologists using point counts [
27]. Each site is represented by a transect comprising approx. 20 sampling points located 300–500 m apart. In total, 2324 point counts were visited. Birds are visited twice per breeding season at each sampling point, to cover both early and late breeders. During one visit, birds detected both visually and acoustically are counted for 5 min at each sampling point. We considered only the counts within a 100-m radius around each sampling point.
In this study, we used the presence and absence of four Columbidae species (specifically,
C. livia,
C. oenas,
C. palumbus,
S. decaocto, and
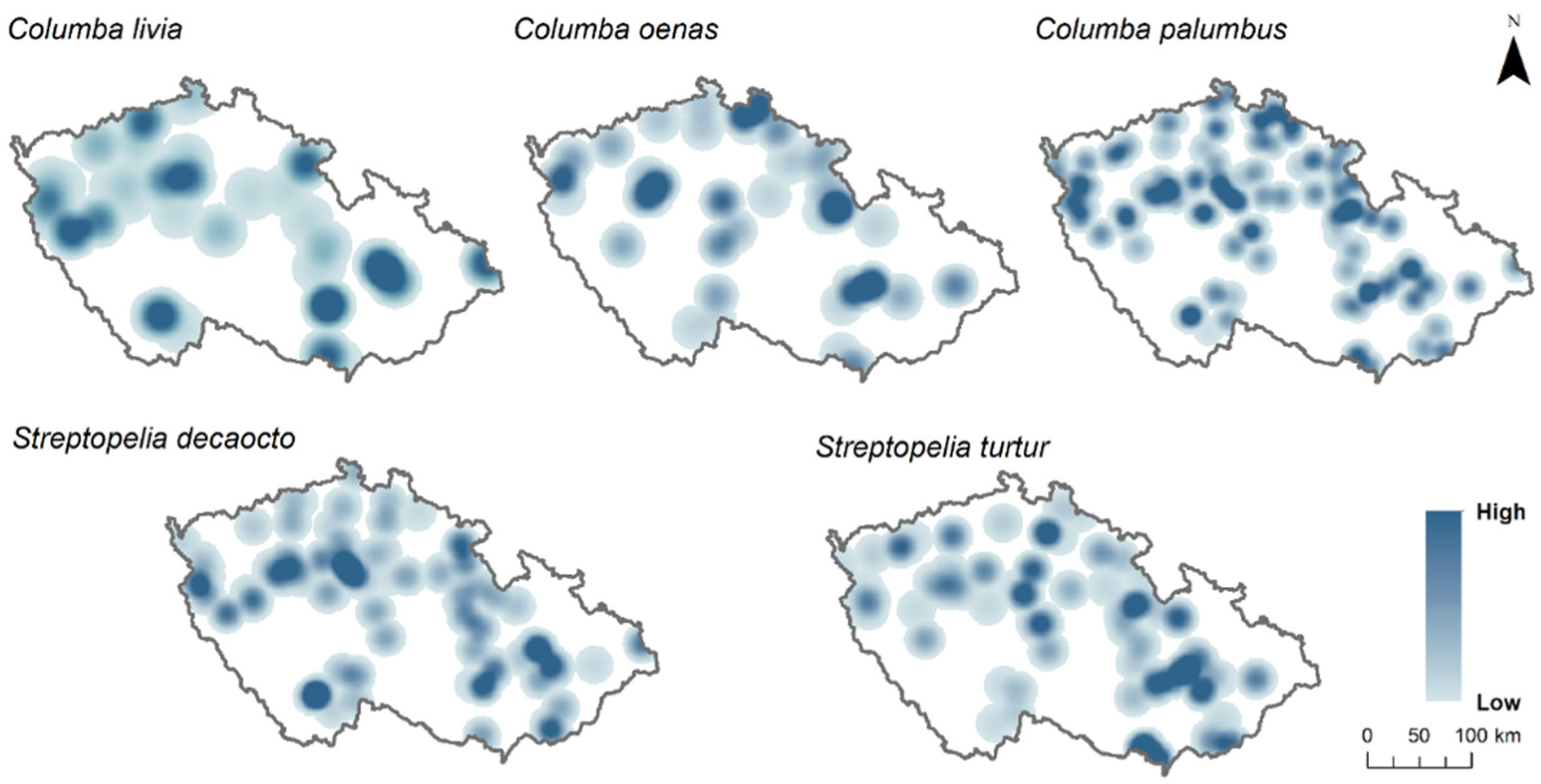
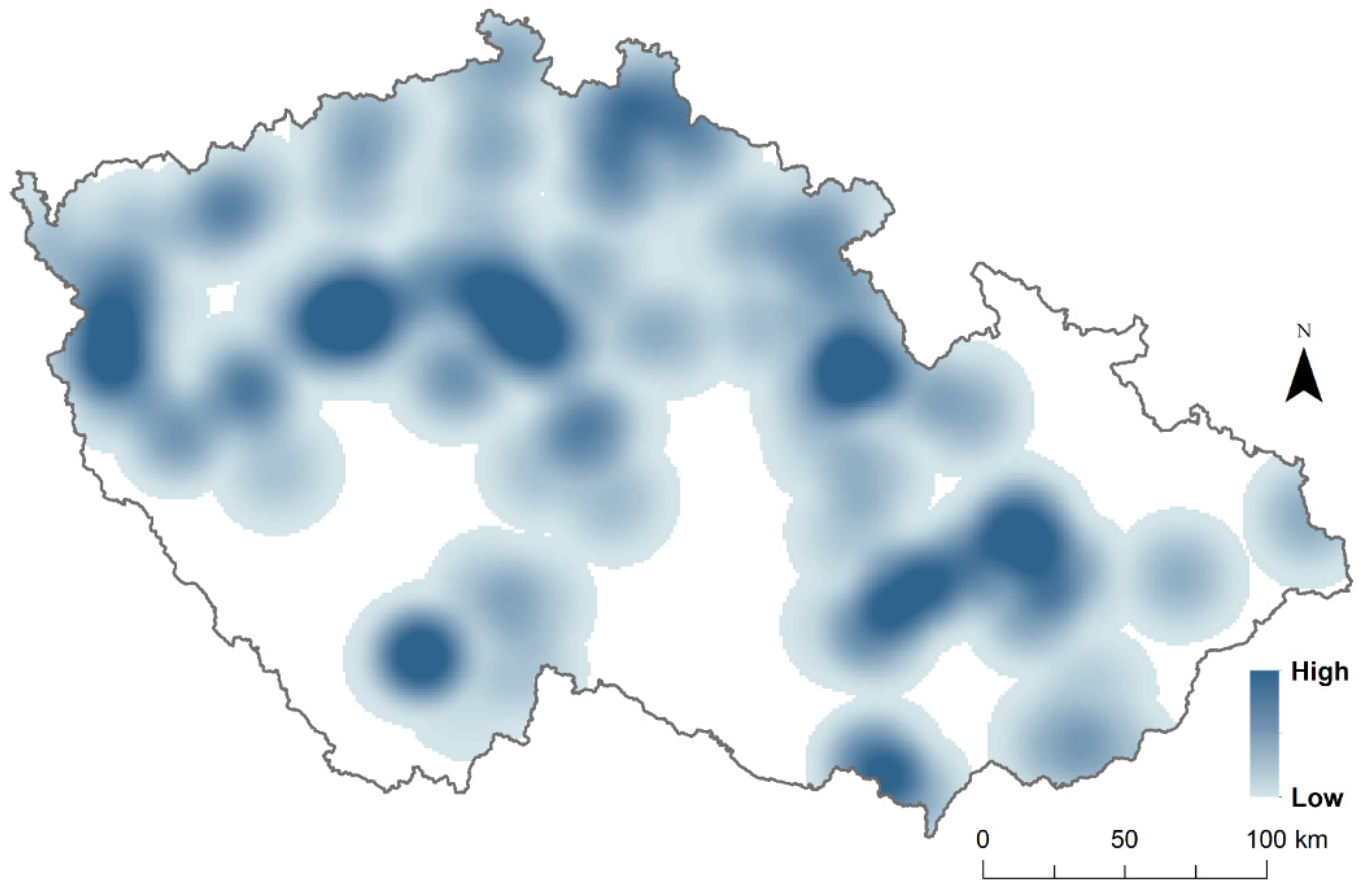
S. turtur) at the sampling points. Presence at sampling points was attributed when the species was observed at least once during the 2 years of observations, while absence was assumed when the species was absent in both survey years. For better visualisation of sampling points where each species was present and species richness per each sampling point, we mapped the sampling points using the Kernel density interpolation method from ArcGIS (
Figure 1 and
Figure 2).
The land use map of the study area was provided by the Nature Conservation Agency of the Czech Republic as the consolidated layer of ecosystems (CLE). CLE is based on a country-wide habitat mapping performed during the early 2000s and updated to 2018 [
28]. We used ESRI 2011. ArcGIS Desktop: Release 10. Redlands, CA, USA: Environmental Systems Research Institute [
29] to calculate land use composition around the 100-m radius of each point count. Land use composition consists of seven land use types: Deciduous, coniferous, and mixed forests, farmlands, grasslands, urban areas, and land use types, which were grouped into one category as “other”, such as shrubs, quarries, rocks, and water. Furthermore, to quantify landscape heterogeneity, we calculated two landscape metrics: Land use richness, which is the number of different land use types per site [
30], and edge density, which is the ratio between total lines or perimeters and total area of each site [
31]. Both landscape metrics were calculated within the 100-m radius of each point count.
We compared the Columbidae species affinities regarding the land use types using the function “habitat overlap”, from the “indicspecies” package in RStudio, which compares pairs of resource niches. The function returns the overlap index between each pair of species [
32], using the amount of resource utilised by each species. The index value ranges from zero (no resources are shared by the two species) to one (all of the resources are shared).
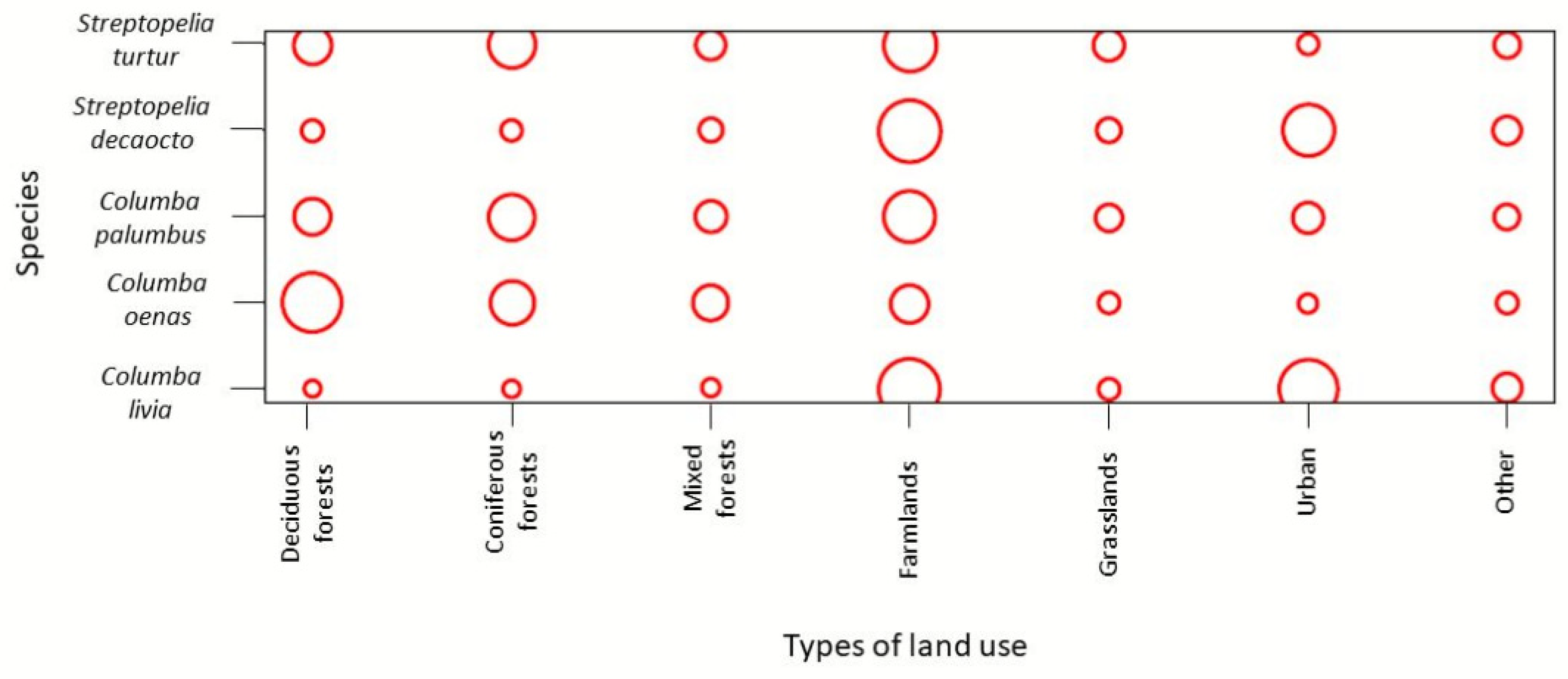
To analyse the differences in utilisation of land use among the species, we used the habitat overlap module from the “EcoSimR” package in RStudio [
33]. It allowed us to plot the resource utilisation matrix. The plot provides a visualisation of the observed utilisation matrix, the area of each circle is proportional to the utilisation of a land use type by each species. If there is no circle, the utilisation matrix is zero.
To analyse the effects of land use types, land use richness, and edge density on species distribution, we performed generalised linear models (GLM) [
34] for each species. To account for the potential spatial autocorrelation (SAC) between sampling points, we applied a Mantel test [
35]. The Mantel statistic (r
M) varies between −1 and +1. It evaluates the similarity between two matrices, first calculated as a geometric distance, and the second one with a geographical distance among the sampling sites [
36]. To test for the significance on the Mantel test, we ran the Monte Carlo permutations with 999 randomisations [
37]. No significant autocorrelation was detected in the dataset used for the analyses (Mantel test, 999 randomisations: r
M = 0.09, simulated
p > 0.05). Furthermore, to assess the relationship between species richness and land use types, as well as between land use richness and edge density, we performed a separate GLM with Poisson distribution.
In the GLM species, presence and absence were used as a response variable assuming a binomial distribution. In addition, land use type (deciduous forest, coniferous forest, mixed forest, urban areas, farmlands, grasslands, etc.), land use richness, and edge density, were used as predictors. After building full models, i.e., the models containing a complete set of predictors, for each species, a test of variance inflation factor (VIF) was applied to check for potential multicollinearity issues among predictor variables, using the function “check_collinearity” from a “performance” package [
38] for RStudio. Only variables with VIF < 6 were introduced in the final models [
39].
All of the statistical tests were performed with RStudio: Integrated Development for R. RStudio, PBC: Boston, MA, USA [
40].
3. Results
The habitat overlap analysis included a total of 2324 sampling points of presence and absence of five Columbidae species, seven different land use types, and two landscape metrics. The most widely distributed species of Columbidae in the Czech Republic was
C. palumbus, with 62% (
n = 1448) of occupied points, followed by
S. decaocto with 21% (
n = 499),
S. turtur with 15% (
n = 357),
C. oenas with 8% (
n = 191), and
C. livia with 7% (
n = 168) of the total occurrence in the country (
Figure 1). In addition, no sampling point had all five species present, nine sampling points had four species present, 131 sampling points had three species present, 558 sampling points had two species present, and 1118 sampling points had one single species present (
Figure 2).
The habitat overlap index based on land use composition at sampling points shows an almost complete habitat overlap between pairs of
C. livia and
S. decaocto, as well as between
C. palumbus and
S. turtur. On the contrary, the smallest habitat overlap was estimated between
C. livia and
C. oenas, as well as between
S. decaocto and
C. oenas. Furthermore,
C. oenas had an 0.8 habitat overlap index with
S. turtur and
C. palumbus (
Table 1). When looking at the mean habitat overlap,
C. palumbus had the highest habitat overlap with the other Columbidae species, followed by
S. turtur. On the contrary,
C. oenas had the lowest mean habitat overlap (
Table 1). According to the utilisation plot, mixed forests, farmlands, grasslands, and other land use types were equally used by all five species, except for
C. oenas, which utilised farmlands less than the other species. In deciduous forests, mostly
C. oenas was present, while in coniferous forests, in addition to
C. oenas,
C. palumbus and
S. turtur were present. Urban areas were inhabited mainly by
C. livia and
S. decaocto (
Figure 3).
The presence of
C. livia was negatively associated with deciduous, coniferous, and mixed forests, farmlands, grasslands, and other types of land use, and positively with edge density (
Table 2).
C. oenas showed negative associations with farmlands and urban areas, and positive associations with deciduous and mixed forests (
Table 3). A model for
C. palumbus showed only the positive associations—with coniferous and mixed forests (
Table 4).
S. decaocto was negatively associated with deciduous, coniferous, and mixed forests, farmlands, grasslands, and other land use types. However, a positive association was observed with edge density in this species (
Table 5).
S. turtur was negatively associated with urban areas and edge density. However, it showed a positive association with grasslands (
Table 6).
Species richness was positively associated with urban areas and edge density. However, coniferous forests and land use richness showed a negative association with species richness (
Table 7).
4. Discussion
Our results confirmed the hypothesis of high habitat overlap among all five Columbidae species in farmland areas. However, they mainly co-exist in three land use types: Farmlands, forests, and urban areas. Furthermore, we recorded an almost complete habitat overlap between C. livia and S. decaocto, as well as between C. palumbus and S. turtur. Regarding land use utilisation, C. livia and S. decaocto mainly utilised the farmlands and urban areas, and avoided all three types of forests (i.e., deciduous, coniferous, and mixed forests), grasslands, and other land use types. At the same time, forests were utilised mainly by C. oenas and C. palumbus. Specifically, C. oenas mainly utilised deciduous forests and avoided the farmlands and urban areas. C. palumbus utilised coniferous and mixed forests. Finally, S. turtur utilised mostly grasslands and avoided urban areas. Regarding species richness, it was higher in urban areas and areas with higher edge density. However, it was lower in coniferous forests and areas with higher land use richness.
Almost all of the complete habitat overlap between
C. livia and
S. decaocto was associated with their presence in areas with higher edge density, i.e., in urban areas. Therefore, habitat overlap between these two species is not surprising since both species are common in urban avian assemblages [
21]. Additionally, high habitat overlap was recorded between
C. palumbus and
S. turtur, mainly in farmlands, deciduous, and coniferous forests. The overlap of these species in farmlands occurred due to the fact that
C. palumbus, which used to be a forest specialist, started colonising other habitats, such as farmlands in the 21st century in the Czech Republic [
41]. In addition,
S. turtur is a highly specialised species that has specific foraging habitat [
42], which depends on open farmlands and grasslands [
23]. Furthermore,
C. palumbus was positively associated with forest, specifically coniferous and mixed stands, probably due to the fact that it depends on the shrubs and trees as nesting habitats [
43]. However, several studies in western and northern Europe have reported that
C. palumbus colonised urban areas in the beginning of the 21st century [
20,
21,
22]. Although our results did not show a positive association between
C. palumbus and urban areas, the species was present in cities. The lack of positive association in our study could be due to the fact that the colonisation process is still in progress since it started later in the eastern parts of Europe [
11], including the Czech Republic or due to the sample bias and lack of sampling points in urban areas.
Regarding
C. oenas, it had the lowest mean habitat overlap with all of the species. Therefore, it was the least congruent with other species distributions. However, the high habitat overlap occurred with
C. palumbus mainly in forests, which aligns with the aforementioned results since both species utilise this land use type. Moreover,
C. oenas was primarily present in deciduous and mixed forests since it depends on old beech trees for nesting [
41]. Consequently, we can say that
C. oenas is a forest specialist, confirmed by previous studies from the Czech Republic that have listed it as a forest species [
44,
45]. Furthermore,
C. oenas showed a negative relationship with urban areas, which is in accord with its aforementioned preference for old trees [
41], which are not common in cities. Regarding habitat overlap,
C. oenas overlapped with
S. turtur, the second highest habitat overlap from all of the other species, which is not surprising since both species feed on farmlands. According to a new study, populations of farmland specialists, including
S. turtur, have declined in Europe since the 1980s [
46]. Therefore, exploring the possibility of using
S. turtur as an umbrella species is useful for future conservation measures.
In general, landscape heterogeneity did not play an essential factor in the spatial distribution of the studied species. The results show that land use richness does not seem to impact the presence or absence of any of the studied species. However, edge density was an important predictor for three species.
C. livia and
S. decaocto are primarily present in urban areas where land use richness is low and edge density is high due to small fragmented patches [
47]. Next,
S. turtur is a farmland specialist, and thus requires open habitats, such as farmlands, for feeding. In addition, it is negatively associated with edge density.
The higher species richness present in urban areas and areas with higher edge density was expected since most of the studied species are associated with cities, except for S. turtur. On the contrary, species richness was lower in coniferous forests and in more heterogeneous areas, in terms of land use richness.
Interestingly, no sampling point had all of the five species present, and only nine sampling points had four species present. From these patterns, we can speculate that an interspecific competition exists between these five species of Columbidae. Furthermore, our predictions that closely related species, pigeons, will have lower habitat overlap than distantly related species, pigeons and doves, were confirmed with several cases. Habitat overlap was the highest between C. livia and S. decaocto; C. palumbus and S. turtur; and finally, between C. palumbus and S. decaocto. However, additional detailed studies are necessary to further confirm this hypothesis.




{kind=link}
{kind=link}
{kind=link}